外来入侵植物剑叶金鸡菊的结实动态和种子萌发特性
2019-02-15,,
,,
(1.贵州民族大学生态环境工程学院,贵阳 550025; 2.凯里学院大健康学院,贵州 凯里 556011)
剑叶金鸡菊(CoreopsislanceolataL.)是菊科金鸡菊属多年生草本植物,头状花序大而艳丽,原产于北美,20世纪初作为观赏花卉引入青岛、庐山等地[1-2]。剑叶金鸡菊生态适应性强,且能借助克隆生长快速扩增形成单一优势种群,排挤本土物种,影响生物多样性[3]。剑叶金鸡菊已从最初引种栽培的庭园逸生、扩散成为入侵物种,在华东、中南广泛分布[4-5],甚至扩散到了西北[6]和西南地区。国内外对剑叶金鸡菊的研究,一方面集中在资源的开发利用,关注剑叶金鸡菊的化学成分及其生物活性[7-8],或是从花卉资源的角度研究剑叶金鸡菊的种子生物学特性[9];另一方面,关于剑叶金鸡菊生物入侵方面的研究也越来越受重视。目前已有关于剑叶金鸡菊的化感作用[10]、光合作用[11]、逆境生理[5]、对环境变化的响应[12]和对群落结构的影响[2,13]等方面的研究,探讨剑叶金鸡菊的生理生态特征与其入侵能力之间的关系。
剑叶金鸡菊有较强的无性生殖能力,在早期通过密集型克隆生长快速扩增来抢占生境,而在后期主要进行游击型克隆生长,形成间隔相对较远的分株,利于种群的扩散[3-4]。入侵物种快速占据新生境、形成优势种群排挤本土物种的能力,还与有性生殖产生种子的潜力和种子萌发成苗的特性密切相关。然而,目前尚未见对剑叶金鸡菊种子产量的动态变化以及不同时期所产种子的萌发能力方面的报道,缺乏种子特性与其入侵机制关系方面的研究。因此,本试验通过对剑叶金鸡菊的结实动态与种子萌发特性进行研究,为探索剑叶金鸡菊的入侵机制和综合治理提供理论依据。
1 材料与方法
1.1 试验材料
剑叶金鸡菊取自贵州省凯里市的一个野外居群(26°31′N,107°53′E,海拔690 m)。试验地位于云贵高原东缘,属亚热带季风区,气候温暖,雨量充沛。剑叶金鸡菊的头状花序生于茎端,在试验地于5月中下旬始花。随着头状花序总苞的展开,周围黄色的舌状花先开,随后中间的管状花由外而内依次开放,每天开1~2轮。在头状花序开放后的6~7 d,外围的舌状花开始枯萎,管状花也从外向内逐渐脱落,整个花序的开放时间为8~9 d。种子成熟所需时间极短,管状花谢花后只需8~9 d,黑色的瘦果即可成熟。剑叶金鸡菊的盛花期可持续到8月上旬,直到11月初还有零星的头状花序开放。剑叶金鸡菊6月初即有种子(瘦果)成熟,一直持续到11月还有零星植株产生少量种子。
1.2 人工授粉试验与传粉者观察
2016年6月,在剑叶金鸡菊的盛花期,以整个头状花序为单位分别进行同株授粉、异株授粉和不授粉3种处理,另设自然对照,每处理设10个重复。3种处理的头状花序均在总苞片张开前套上硫酸纸袋,进行相应处理后继续套袋至花序开放完毕。同株授粉处理用同株上的不同花序(套袋防止异源花粉污染)进行人工授粉,而异株授粉采用的是20 m以外植株的花粉。为保证授粉效果,每种授粉处理每天进行1次,连续进行3~4 d。全部处理的头状花序均在总苞开放后用相机记录管状花总数,结实后统计各处理中头状花序产生的种子数。结实率(%)=(种子数/管状花序数)×100%。选择晴好天气,结合样线法与定点观察法进行访花昆虫调查。用相机拍摄访花昆虫,结合用捕虫网捕捉传粉昆虫标本,鉴定主要传粉昆虫种类。
1.3 开花动态和种子产量测算
通过前期观察,确定剑叶金鸡菊在6—11月均能产生种子,但种子产生的高峰主要在前期,而10—11月仅有零星的植株产生零星的种子。故本试验重点考察从始花到9月底的花序数量和种子产量。2017年6月,在样地内随机选取18株剑叶金鸡菊挂牌标记,统计始花期至6月底、6月底至7月底、7月底至8月底、8月底至9月底共4个时段内各植株产生的已谢花结实的花序数。在每个时段内,每个植株随机选取5个结实的花序,考察种子数量。将花序数和5个花序的平均种子数相乘,估算每株剑叶金鸡菊在该时段内产生的种子总数。8月底至9月底部分植株的花序数少于5,则以实际产量为准。最后将各时段的花序数和种子数量累计相加,算出一株剑叶金鸡菊年产花序数,估算出种子的年产量。
1.4 种子萌发和出苗试验
分别在6月底采集早期成熟的种子,在10月初采集晚期成熟的种子,采后立即开展种子萌发和出苗试验。其中,种子萌发试验在2016年和2017年连续进行了2年,而出苗试验只在2017年进行了1年。种子萌发试验在铺有浸湿脱脂棉的培养皿中进行,每个培养皿中随机放置50粒形态大小基本一致的种子,设3次重复。将培养皿放置在光照状况相对良好的试验室边台上,保持窗户开放,使试验条件尽可能接近室外自然状况。培养过程中每天换一次蒸馏水并记录发芽情况。当胚根长度约为种子直径的一半时视为种子发芽,培养时间为3周。
发芽率(%)=(萌发的种子总数/50)×100%;
平均发芽时间=∑(ti×ni)/∑ni;式中ti为从试验开始的天数,ni为当天发芽的种子数。
出苗试验和萌发试验操作类似,区别在于用花盆代替培养皿,基质为取自剑叶金鸡菊生境中去除表土后混匀的泥土。每个花盆中放置50粒种子,浇水使种子半没于土中,设3次重复。培养地点和发芽试验一致,每天浇水,并记录出苗情况,培养时间为4周。
出苗率(%)=(出苗的种子总数/50)×100%;
平均出苗时间=∑(ti×ni)/∑ni;式中ti为从试验开始的天数,ni为当天出苗的种子数。
1.5 数据分析
对人工授粉试验的结实率以及不同时期的头状花序数量和种子产量数据进行单因素方差分析,并利用LSD检验进行不同处理间的比较。对早期和晚期种子的发芽和出苗数据进行t检验。所有数据分析均用SPSS 19.0统计软件完成。
2 结果与分析
2.1 剑叶金鸡菊的繁育系统
如表1所示,剑叶金鸡菊自然对照的结实率最高,异花授粉处理结实率次之,而套袋后不授粉和自花授粉处理几乎都不结实。单因素方差分析结果表明,套袋处理对剑叶金鸡菊的结实率产生了极显著的影响(F=64.092,p<0.001)。自然对照与其它处理间的差异均达到了极显著水平(p<0.001),异花授粉处理与套袋后不授粉,以及自花授粉2种处理间的差异也都达到了极显著水平(p<0.01)。由此判断,剑叶金鸡菊是自交不亲和的物种,需要依靠异交产生种子。观察到访问剑叶金鸡菊的昆虫共有12种,其中传粉昆虫有中华蜜蜂(Apiscerana)、幽矍眼蝶(Ypthimaconjuncta)、黄粉蝶(Euremablanda)、无标黄粉蝶(Euremabrigitta)、鹿蛾(Amatasp.)、食蚜蝇(Syrphidaesp.)、黑额光叶甲(Smaragdinanigrifrons)、叶甲(Chrysomelidaesp.)等。能帮助剑叶金鸡菊实现有性生殖的传粉昆虫种类众多,蜂类、蝶类、蛾类、蝇类和甲虫类均有,最主要的传粉昆虫为中华蜜蜂(A.cerana)。
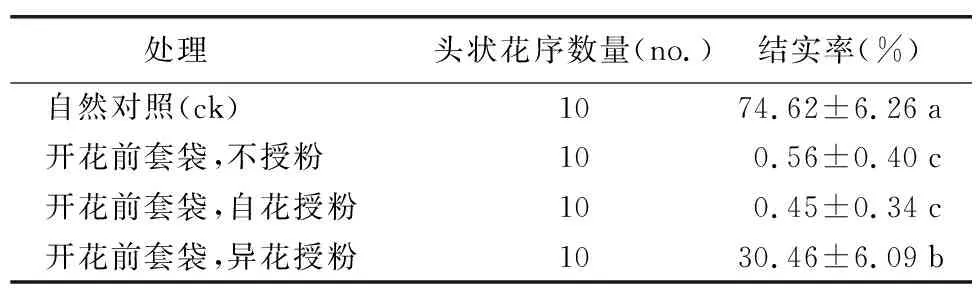
表1 人工授粉试验结果
注:表中数值为平均数±标准误,同一列内不同小写字母表示处理间的差异显著(p<0.05)。下同。
2.2 剑叶金鸡菊的花序数量和种子产量
单因素方差分析结果表明,剑叶金鸡菊的头状花序数(F=24.162,p<0.001)、单花序种子数(F=48.645,p<0.001)和单株种子产量(F=33.704,p<0.001)在不同时期均有极显著的差异。3个指标的最高值均在早期获得,而随着花期的持续,头状花序数、单花序种子数和单株种子产量均呈下降趋势(表2)。除了7月份和8月份的头状花序数之间、8月份和9月份的单花序种子数之间没有显著差异外,3个指标在其它时期之间的差异均达到了显著水平(p<0.05)。平均每株剑叶金鸡菊可产生约135个头状花序,每个花序平均产生约93粒种子,每株的种子总产量约12 600粒(5 128~25 433粒,n=18)。值得注意的是,6月底前产生的种子平均达到了7 740.50粒,说明剑叶金鸡菊的种子主要是在早期产生的。

表2 各时期头状花序数量和种子产量
2.3 剑叶金鸡菊的种子萌发和出苗
如表3所示,2年的种子萌发试验均表明,后期成熟的种子(10月初采集)发芽率高于前期成熟的种子(6月底采集)。但只有在2017年的试验中,二者发芽率之间的差异达到了极显著水平(t=12.438,p<0.001)。无论是前期还是后期采集的种子,在室温下培养,2~3 d即有种子陆续发芽,发芽率48%~86%,3~4 d即有种子开始出苗,两期种子出苗率均超过了50%。在平均发芽时间、出苗率和平均出苗时间方面,前期成熟的种子和后期成熟的种子之间,均不存在显著差异(表3)。以上试验结果说明,剑叶金鸡菊种子成熟后不存在休眠特性,有立即萌发成苗的潜力。
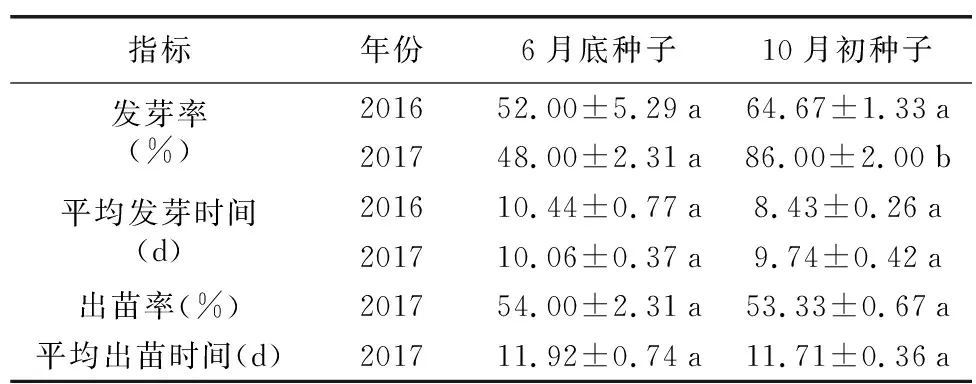
表3 早晚期种子的发芽和出苗情况
3 结论与讨论
剑叶金鸡菊套袋后不授粉,或者自花授粉处理后,绝大多数花序都不结实,说明该物种自交不亲和。本研究结果与曾建军等[4,14]的研究结果均表明,剑叶金鸡菊的交配系统为异交,种子需要通过不同植株之间的异交产生。剑叶金鸡菊虽需异交才能结实,但能为其传粉的昆虫众多,泛化的传粉系统使得剑叶金鸡菊能相对轻易地获得异源花粉,不影响种子的产生。本试验只是在白天对访花昆虫进行了调查,考虑到夜间活动昆虫传粉的可能性,估计其有效的传粉者应该更多。剑叶金鸡菊克隆生长能力强,在早期通过密集型生长抢占生境,在后期则通过游击型生长分散同一基因型植株在种群中的分布[3-4]。有性生殖产生的种子利于入侵物种的扩散和初期种群的建立,而无性生殖则有利于在新生境中快速形成单一优势种群[15]。剑叶金鸡菊可能正是通过同时发挥有性生殖和无性生殖的双重优势,才成为广泛分布的入侵物种。
另外,发现管状花中有蓟马活动,而套袋处理无法隔离此类小型昆虫。这类小型昆虫带来的外来花粉,可能是套袋自花授粉以及套袋不授粉处理中有极个别种子产生的原因。剑叶金鸡菊头状花序中,能产生种子的是中间逐步成熟的管状花。在进行人工授粉操作时,一个花序只有部分花开放,而且花粉孔裂,能用于授粉的花粉数量较少。基于剑叶金鸡菊的开花和散粉特点,即便进行了多次授粉,但花粉量依然可能不足,这应该是造成人工异花授粉结实率偏低的原因。
剑叶金鸡菊花序多,种子产量大,有性生殖能力极强,本研究中高产植株的种子总产量超过了25 000粒。剑叶金鸡菊在入侵日本后平均每个花序产生98粒种子,是北美原产地的两倍多[16],和本研究的结果较为相近。入侵物种一般具有种子产量大、易于萌发的特点,因而扩散和传播能力强[17]。平均每株剑叶金鸡菊种子产量超过12 000粒,其中大部分是在6月底前产生的,且种子成熟后立即可以发芽和出苗,在当年就具备极强的种群建成潜力。合适的温度更适宜剑叶金鸡菊种子发芽[18]。本研究中,种子发芽和出苗试验分别在夏季和初秋进行,10月份的温度更为温和,这可能是后期采收种子的发芽率和出苗率均较高的原因。即便因水分限制等原因,条件暂时不适宜,剑叶金鸡菊的种子也可以在地下保持活力,转入休眠状态,形成种子库,以躲避不良环境的影响[19]。剑叶金鸡菊种子产量巨大,而这种二次休眠特性更是有助于其占据新生境、形成新种群,强化其入侵能力。
剑叶金鸡菊具有花期长、花序数多,有效的传粉昆虫多、结实早、种子产量高、种子萌发率和出苗率高等特征,表明其有性繁殖系数极高,扩散风险极大。结合剑叶金鸡菊的结实特征和萌发特性,采取防除措施不能迟于有性生殖的早期,以防止种子的产生。下一步,可以动态监测不同时期成熟的种子落地后的成苗和后续生长状态,以便在自然状态下,更准确地评价剑叶金鸡菊的有性繁殖种群建成能力。
