花生地方品种骨干种质的遴选
2019-02-10闫彩霞李春娟郑奕雄韩柱强陈静王娟单世华
闫彩霞 李春娟 郑奕雄 韩柱强 陈静 王娟 单世华

摘要:我国花生地方品种众多,遗传多样性丰富,是花生育种的主要亲本来源。本研究以259份初选花生骨干种质为材料,采用“逐步UPGMA聚类+个体优先取样”策略,首次构建了包含171份种质的中国花生地方品种骨干种质,占全部种质的6.24%,遗传代表性为82.4%。利用t测验、F测验、卡方测验、极差、表型保留比例等对骨干种质的代表性进行检验和评价,结果表明:骨干种质的大多数性状与全部种质无显著差异,植物学类型组成和生态分布一致,且保持了全部种质的表型分布频率和表型相关性。本研究构建的骨干种质具有很好的代表性,可为花生种质发掘和品种遗传改良奠定良好的基础。
关键词:花生;地方品种;骨干种质;代表性评价
中图分类号:S565.202.4 文献标识号:A 文章编号:1001-4942(2019)12-0001-06
Abstract Peanut landraces with rich genetic diversity in China are the main parent sources. In the study, we primarily chose 259 peanut germplasms as material, and 171 key germplasms were constructed for the first time by the strategy of stepwise UPGMA clustering approach and preferred sampling, which accounted for 6.24% of the total germplasms and the genetic representativeness was 82.4%. We further verified and evaluated the 171 germplasms by t-test, F-test, chi-squared test, range and the ration of phenotypic retention. The results showed that there were no significant differences between the key germplasms and the total germplasms in the majority of thirteen traits. Additionally, the composition of botanical type and ecological distribution of key germplasms corresponded very well with those of the total germplasms. Thus, the key germplasms had preserved the phenotypic distribution frequency and phenotypic correlation of the total germplasms. The key germplasms constructed in the study was very representative and could lay a good foundation for peanut germplasm excavation and genetic improvement.
Keywords Peanut; Landraces; Key germplasm; Representative evaluation
花生(Arachis hypogaea L.,2n=4x=40),又名落花生、長生果、地豆等,属蝶形花科落花生属一年生草本植物,起源于玻利维亚南部和阿根廷北部地区。我国收集保存于种质库中的花生资源已达7 400余份,包括地方品种4 000余份,其中用作花生育种亲本的约有400个,直接或间接育成大面积推广品种408个,极大地推动了花生的产业发展。
核心(骨干)种质能最大限度地代表整个资源的遗传多样性,水稻[1]、玉米[2]、大豆[3]等作物已经构建了核心种质,小麦[4]、糜子[5]、陆地棉[6]则建立了骨干种质,并进行了广泛的鉴定与评价,极大地提高了种质资源的利用效率[7]。花生方面,国际半干旱作物研究所(ICRISAT)基于植物分类、地理来源和表型描述构建了包含1 704份材料的核心种质,占全部种质的11.9%[8];美国利用6个形态性状数据构建起包含831份材料的核心种质,占全部种质的11.2%[9];我国则根据15个性状数据,构建了由576份资源组成的核心种质,占全部种质的9.0%[10]。然而,对于一些多年多点数量性状的取得和某些性状的抗性鉴定,这样的群体规模偏大,利用不便。因此,Upadhyaya等[11]提出了微核心种质(mini core collection)的概念,将核心种质进一步压缩,仅用1%的样品数代表整个群体大部分的有用变异。
目前国内水稻、小麦、大豆、花生等作物业已构建微核心种质,其提取规模约在2%~5%[4,12-14]。本研究以259份中国花生地方品种初选骨干种质为材料,利用13个表型数据循环聚类,结合优先取样法,构建中国花生地方品种骨干种质,并对其代表性进行评价,以期为花生种质发掘和品种遗传改良奠定良好基础。
1 材料与方法
1.1 材料及数据整理
以259份中国花生地方品种初选骨干种质为研究对象,涉及生育期、百果重、百仁重、出仁率、株高、株型、开花习性、分枝型、植物学类型共9个农艺性状和粗蛋白、粗脂肪、油酸、亚油酸含量共4个品质性状。质量性状根据表型进行赋值。其中,株型:直立=1,半蔓=3,匍匐=5;开花习性:交替=1,连续=3;分枝型:密枝=1,疏枝=3;植物学类型:多粒型=1,珍珠豆型=2,龙生型=3,普通型=4,中间型=5。数量性状数据采用0.5个标准差为间距进行标准化,共分为10级:1级≤X-2δ,10级>X+2δ,中间每级间差为 0.5δ(X为性状平均值,δ为标准差)。
1.2 取样策略
利用SPSS 22.0统计分析软件,对初选骨干种质的26个分组分别进行各性状的逐步UPGMA聚类分析。遗传距离相同或极相近的同类材料优先选择具有最大或最小性状表型值的种质,进入下一轮聚类;如组内仅一份材料,则该材料直接进入下一轮聚类分析。计算每一轮聚类后入选种质的遗传代表性估计值(genetic estimate,GE),GE(%)=1-│1-RPR│,RPR为表型保留比例(ration of phenotypic retention),公式为:RPR=∑ni=1Mi/∑ni=1Mio(式中Mio为全部种质中某性状的表型变异数,Mi为骨干种质中该性状的表型变异数,n为性状总数),直至遗传代表性接近70%,并适当补充极值材料和特殊性状种质。
1.3 骨干种质的评价
利用SPSS 22.0中的t测验、F测验、χ2测验等评价骨干种质在均值、方差、表型分布频率、变异系数、Shannon-Weaver多样性指数(香农多样性指数,H′)等指标上是否很好地代表初选骨干种质和全部种质的遗传多样性。极差分析、表型保留比例和表型相关性分析分别用于确定全部种质和初选骨干种质的变异范围、变异丰度和性状相关性是否在骨干种质中得到了相应保持。
2 结果与分析
2.1 骨干种质的构建、植物学类型组成及生态分布
259份初选骨干种质,经多次UPGMA聚类分析后共选取161份资源,初步评价后补充了极值材料和特殊性状种质10份,构建了由171份资源组成的中国花生地方品种骨干种质,占全部种质的6.24%,对全部种质的遗传代表性达82.4%,对初选骨干种质的遗传代表性达99.2%(表1)。骨干种质包含全部花生植物学类型,其中多粒型13份(7.6%)、珍珠豆型52份(30.4%)、龙生型24份(14.0%)、普通型72份(42.1%)、中间型10份(5.9%)。卡方测验表明,骨干种质很好地代表了初选骨干种质(P=0.907)和全部种质(P=0.450)的植物学类型组成。此外,骨干种质覆盖了全部7个花生生态区,其中黄河流域55份(32.2%)、长江流域44份(25.7%)、东南沿海44份(25.7%)、云贵高原10份(5.9%)、黄土高原4份(2.3%)、东北12份(7.0%)和西北2份(1.2%),代表了初选骨干种质(P=0.941)和全部种质(P=0.592)的生态分布。
2.2 平均值、方差和极差比较
平均值t测验表明,骨干种质的生育期与初选骨干种质存在显著差异粗蛋白含量与全部种质存在极显著差异,粗脂肪含量与初选骨干种质存在显著差异,其余性状在三个群体中无明显差异,且骨干种质中植物学类型、株型、分枝型、生育期、百果重、百仁重、粗脂肪含量等性状的平均值都大于初选骨干种质和全部种质(表2)。F测验表明,除骨干种质的粗蛋白含量与全部种质差异显著外,其余性状的方差均为齐性,无显著差异(表2)。极差分析表明,除百仁重和粗蛋白含量外,骨干种质保留了全部种质其余性状81%~100%的变异(表2)。从上述结果可见,骨干种质遗传冗余度小、变异率高,对初选骨干种质和全部种质均有很好的代表性。
2.3 变异系数和香农多样性指数的比较
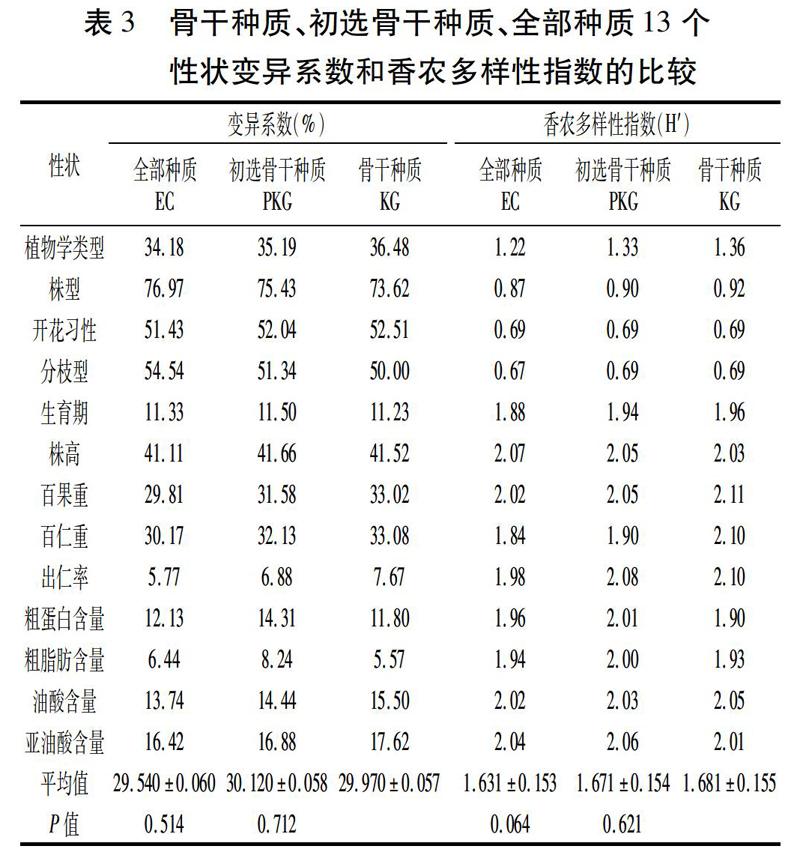
由表3可以看出,骨干种质、初选骨干种质、全部种质13个性状的香农多样性指数(H′)和变异系数非常相似:骨干种质的香农多样性指数平均值为1.681,变异系数为29.970%;初选骨干种质这两个指标的值分别为1.671和30.120%;全部种质则为1.631和29.540%。可以看出,骨干种质和初选骨干种质这两个指标的值略高于全部种质。成对双样本t测验表明,骨干种质H′与初选骨干种质、全部种质的差异均不显著(P=0.621,P=0.064),变异系数也无显著差异(P=0.712,P=0.514),说明骨干种质的样本足够大,且有效去除了总资源中的冗余,保留了全部种质的遗传多样性。
2.4 分布频率和表型保留比例的比较
由表4可以看出,全部种质、初选骨干种质13个性状的102个表型分级均保留在骨干种质中。对13个性状的分布频率进行卡方测验,差异均不显著,表明三个样本的性状分布是一致的,骨干种质可代表全部种质的变异。植物学类型、百仁重和粗脂肪含量的表型保留比例较大,是补充了一些特殊种质和极值材料所致;其余10个性状的表型保留比例均稍高于全部种质,表明骨干种质保留了全部种质丰富的变异,且丰度更高。
2.5 表型相关性分析
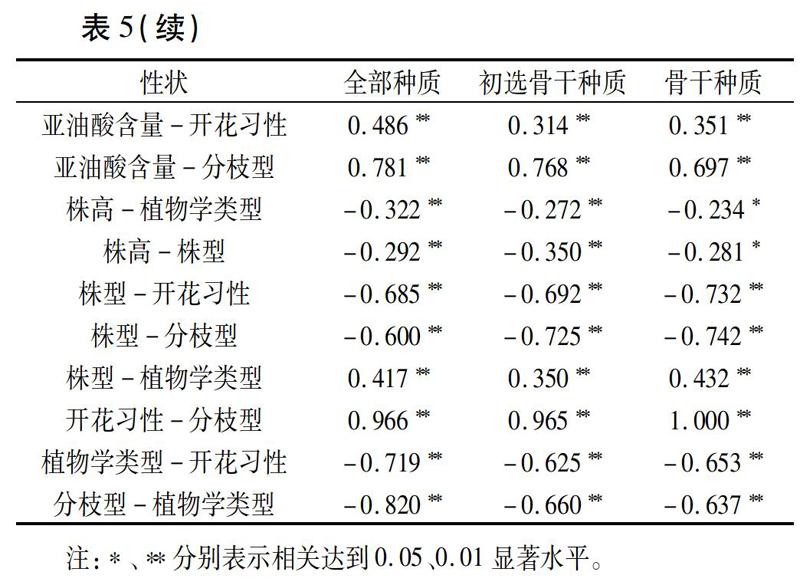
骨干种质除应具有较小的遗传冗余外,还应保留原群体固有的性状间的遗传关联。对13个性状的表型相关性进行分析,38对性状在全部种质中呈极显著正相关,其中有35对与初选骨干种质中的相关性是一致的,32对与骨干种质中的相关性一致(表5),即骨干种质对初选骨干种质和全部种质极显著关联性状的保持率分别为91.4%和84.2%。因此,骨干种质很好地保持了全部种质的表型相关性。不同性状间的极显著相关性意味着在育种实践中,可以不直接筛选难测量的性状,而优先筛选与其极显著相关的易测量性状,从而较快地实现育种目的。
3 讨论
3.1 骨干种质的构建策略
对于大部分遗传资源非常丰富的农作物而言,只有建立尽可能小的骨干群体,才能实现资源利用的最大化,但需保留原始群体至少70%的遗传变异[15]。“循环系统聚类+优先取样法”是构建核心(骨干)种质的常用策略,该方法已用于葡萄[16]、甘蔗[17]、水稻[18]、大豆[19]和小麦[20]的核心种质构建。为使构建的骨干种质能以最小的样本量最大限度地代表原始群体的遗传多样性,就要不断地评估筛选种质的遗传代表性。本研究在每一轮聚类后,都要计算入选种质的遗传代表性估计值,使其不低于70%,再适当补充极值材料和特殊性狀种质,最终构建起代表全部种质82.4%遗传变异、代表初选骨干种质99.2%遗传变异的中国花生地方品种骨干种质。
3.2 骨干种质的评价与确认
骨干种质的评价就是检验其代表性和有效性。李自超等[21]认为香农多样性指数、表型方差、表型分布频率、变异系数、表型保留比率等是衡量骨干种质代表性的重要参数。本研究通过t测验、F测验、卡方测验对骨干种质与初选骨干种质、全部种质进行了比较,结果显示:大多数性状的均值、方差、极差、变异系数、香农多样性指数、分布频率等无显著差异;三者在植物学组成、生态分布上也有很好的一致性;同时,骨干种质具有更高的表型保留比例,表明与初选骨干种质和全部种质相比,骨干种质的丰度更高。此外,性状间的相关是作物的一个内在特性,是控制性状的基因存在某种关联的外在表现,抽样不应改变性状间的相关性,因此可通过比较骨干种质与全部种质性状间的相关系数来确认其代表性[22]。本研究构建的骨干种质对初选骨干种质和全部种质极显著关联性状的保持率分别为91.4%和84.2%,表明骨干种质很好地代表了初选骨干种质和全部种质各性状间固有的遗传相关。
4 结论
利用“循环系统聚类+优先取样法”策略,构建了171份中国花生地方品种骨干种质,占全部种质的6.24%,遗传代表性达82.4%。骨干种质大多数性状的均值、方差、极差、变异系数、香农多样性指数、表型保留比例等与全部种质无显著差异,植物学类型组成和生态分布一致,且保持了全部种质的表型分布频率和表型相关性。本研究建立的骨干种质具有很好的代表性。
参 考 文 献:
[1] Zhang H,Zhang D,Wang M,et al. A core collection and mini core collection of Oryza sativa L. in China[J]. Theoretical and Applied Genetics,2011,122(1):49-61.
[2] Li Y,Shi Y S,Cao Y S,et al. Establishment of a core collection for maize germplasm preserved in Chinese National Genebank using geographic distribution and characterization data[J]. Genetic Resources and Crop Evolution,2005,51(8):845-852.
[3] Oliveira M F,Nelson R L,Geraldi I O,et al. Establishing a soybean germplasm core collection[J]. Field Crops Research,2010,119(2/3):277-289.
[4] 关正君. 小麦骨干种质HMW-GS与面包品质关系的研究[D].保定:河北农业大学,2003.
[5] 董俊丽,王海岗,陈凌,等. 糜子骨干种质遗传多样性和遗传结构分析[J]. 中国农业科学,2015,48(16):3121-3131.
[6] 张小娟,何团结,陆徐忠,等. 陆地棉SSR核心引物筛选及95份骨干种质的遗传多样性分析[J]. 棉花学报,2011,23(6):529-536.
[7] 贾继增,高丽锋,赵光耀,等. 作物基因组学与作物科学革命[J]. 中国农业科学,2015,48(17): 3316-3332.
[8] Upadhyaya H D,Ortiz R,Bramel P J,et al. Development of a groundnut core collection using taxonomical,geographical and morphological descriptors[J]. Genet. Resour. Crop Evol.,2003,50(2):139-148.
[9] Holbrook C C,Anderson W F,Pittman R N. Selection of a core collection from the U.S. germplasm collection of peanut[J]. Crop Sci.,1993,33(4):859-861.
[10]姜慧芳,任小平,廖伯寿,等. 中国花生核心种质的建立[J]. 武汉植物学研究,2007,25(3): 289-293.
[11]Upadhyaya H D,Bramel P J,Ortiz R,et al. Developing a mini core of peanut for utilization of genetic resources[J]. Crop Sci.,2002,42(6):2150-2156.
[12]Hao C,Dong Y,Wang L,et al. Genetic diversity and construction of core collection in Chinese wheat genetic resources[J]. Chinese Science Bulletin,2008,53(10):1518-1526.
[13]邱丽娟,李英慧,关荣霞,等. 大豆核心种质和微核心种质的构建、验证与研究进展[J]. 作物学报, 2009,35(4):571-579.
[14]姜慧芳,任小平,黄家权,等. 中国花生小核心种质的建立及高油酸基因源的发掘[J]. 中国油料作物学报,2008,30(3):294-299.
[15]Brown A H D. Core collections:a practical approach to genetic resources management[J]. Genome,1989,31(2):818-824.
[16]郭大龙,刘崇怀,张君玉,等. 葡萄核心种质的构建[J]. 中国农业科学,2012,45(6):1135-1143.
[17]刘新龙,刘洪博,马丽,等. 利用分子标记数据逐步聚类取样构建甘蔗杂交品种核心种质库[J]. 作物学报,2014,40(11):1885-1894.
[18]李自超,张洪亮,曾亚文,等. 云南地方稻种资源核心种质取样方案研究[J]. 中国农业科学,2000, 33(5):1-7.
[19]赵丽梅,董英山,刘宝,等. 中国一年生野生大豆(Glycine soja)核心资源的构建[J]. 科学通报,2005,50(10):992-999.
[20]董玉琛,曹永生,张学勇,等. 中国普通小麦初选核心种质的产生[J]. 植物遗传资源学报,2003, 4(1):1-8.
[21]李自超,张洪亮,曹永生,等. 中国地方稻种资源初级核心种质取样策略研究[J]. 作物学报,2003, 29(1):20-24.
[22]劉遵春,张春雨,张艳敏,等. 利用数量性状构建新疆野苹果核心种质的方法[J]. 中国农业科学, 2010,43(2):358-370.
