鸡枞菌多糖对鲫的生长性能、抗氧化及免疫功能的影响
2019-01-22王思芦王红明赵艮勇肖文渊黄志秋
王思芦,王红明,赵艮勇,肖文渊,黄志秋
(西昌学院动物科学学院,四川西昌615013))
随着水产动物网箱养殖的出现和快速发展,高密度的养殖方式带来高效益的同时也伴随着许多问题,如造成鱼类的应激增强而引起免疫力下降,容易爆发传染性疾病,长期使用化学药物防治鱼病容易引起水体中病原耐药性增加和产生药物残留等生态问题。因此,开发利用具有多功能的天然无公害的药物,实现促生长、提高动物抗氧化和免疫能力,是现代养殖业迫切需求。研究表明,多糖对水产动物的免疫功能、抗氧化能力具有增强作用,如在异育银鲫饲料中添加酵母细胞壁多糖,可提高其血清抗体效价以及血液中白细胞的吞噬能力[1],在罗非鱼的饲料中添加黄芪多糖,可以显著提高血浆中的CAT、SOD的含量[2]。前期研究表明,鸡枞菌多糖具有显著的免疫增强活性[3-4]和体外抗氧化活性[5-7],但其对水产动物的体内免疫活性及抗氧化作用国内外尚未见报道。本研究旨在探讨鸡枞菌多糖(TAP)对鲫的肥满度、体内抗氧化能力及溶菌酶水平的影响,为研发新型安全高效的鱼用保健品提供参考。
1 材料与方法
1.1 材料
鲫和通威鲫饲料均购自市场、黄芪多糖(APS)和鸡枞多糖(TAP)均由西昌学院动物科学学院基础医学实验室制备,丙二醛(MDA)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)、溶菌酶(LZM)检测试剂盒均购自南京建成生物工程研究所。
1.2 方法
1.2.1 含药饲料的制备
将相应添加量的APS或TAP溶解于适量蒸馏水,对各组日粮进行充分浸泡,待药液被完全吸收后,于40 ℃烘干装入密封袋备用。
1.2.2 动物分组与处理
选取(8±0.2)g的健康鲫175尾,随机分成5组,每组35尾,分别为空白组、黄芪多糖对照组(APS添加量为日粮的1%)、TAP高、中、低剂量组(TAP添加量分别为日粮的1%、0.5%、0.1%),日粮按各组鲫总体重的3%分为早、晚两次投喂。分别在给药后的第7、14天各采样一次,每次各组随机选取15尾鱼,称量每尾鱼的体重、体长,肝胰脏和头肾组织重量,记录结果。
1.2.3 TAP对鲫肥满度和肝体比的影响
鱼肥满度是鱼体重量与鱼体体长立方数的比值,是反映鱼类肥瘦程度和生长情况的重要指标。分别将各组给药后的第7、14天称量鱼的体重、体长和肝胰脏重量,按以下公式计算肥满度(K)和肝体比(HIS)。
K=(W/L3)×100
HIS=G/W×100%
式中W为体重(g),L为体长(cm),G为肝胰脏重量(g)。
1.2.4 鲫肝胰脏匀浆的制备
使用球磨仪每秒振动150次,研磨30 s,按组别分别将每一尾鲫的0.1 g肝胰脏以4 ℃ 0.65%生理盐水制备成10%的匀浆液,将匀浆液以3 000 r/m离心10 min,取上清液于4 ℃保存待测。
1.2.5 TAP对鲫肝胰脏抗氧化性能的影响
取1.2.4制备的样品按照检测试剂盒使用说明分别进行各组MDA、CAT、SOD的检测,方法参照试剂盒说明书进行。
1.2.6 TAP对鲫免疫功能的影响
分别按以下公式计算给药后第7和第14天各组鲫的头肾体指数(HKBI)。HKBI=TS/W×100%,式中TS为头肾重(g),W为体重(g)。
取1.2.4制备的样品按照检测试剂盒使用说明进行各组LZM含量的检测。
1.2.7 数据统计与分析
各组试验数据以平均值±标准误(SE)表示,并采用SPSS 20.0软件中 ANOVA 模型进行单因素方差分析,采用 LSD 方法进行多重比较,P<0.05表示组间差异显著,P<0.01表示组间差异极显著。
2 结果
2.1 TAP对鲫肥满度和肝体比的影响
结果表明,APS及TAP各剂量组均可不同程度地提高鲫肥满度。给药7 d后,TAP高剂量组比空白组提高了69.2%;其余各组间差异不显著。给药14 d天后,各组间肥满度差异均不显著。结果反映了高剂量TAP在7 d内可能对鲫有促进生长的作用,效果优于相同剂量的APS,结果见表1 。
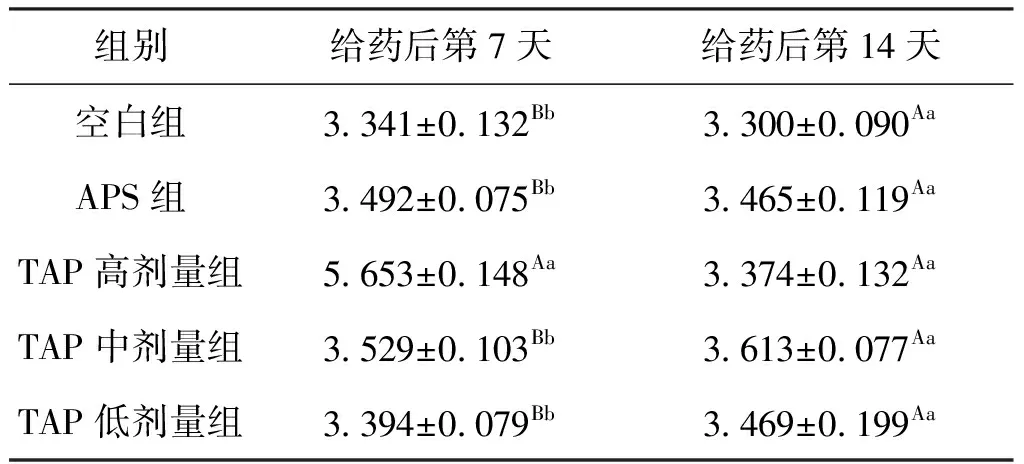
表1 TAP对鲫肥满度的影响Tab.1 Effect of TAP on fullness of C.auratus %
注:同一列上标不同大写字母表示差异极显著,P<0.01;上标不同小写字母表示差异显著,P<0.05 。下表同。
肝胰脏是鱼体最大的消化腺,具有分泌胆汁和消化酶功能。给药7 d后各组肝体比均高于空白组,其中APS组的肝体比提高了21.77%,TAP中剂量组的肝体比提高了21.29%,但各组间无显著差异;给药14 d后,各组肝体比仍然高于空白组,其中APS组的肝体比最高,比空白组提高了13.06%,而TAP中剂量组则提高了10.4%。结果表明,APS和TAP对鲫的肝胰脏的代谢有一定的影响,但这种影响在试验期间不显著,可能与给药时间较短有关,结果见表2。
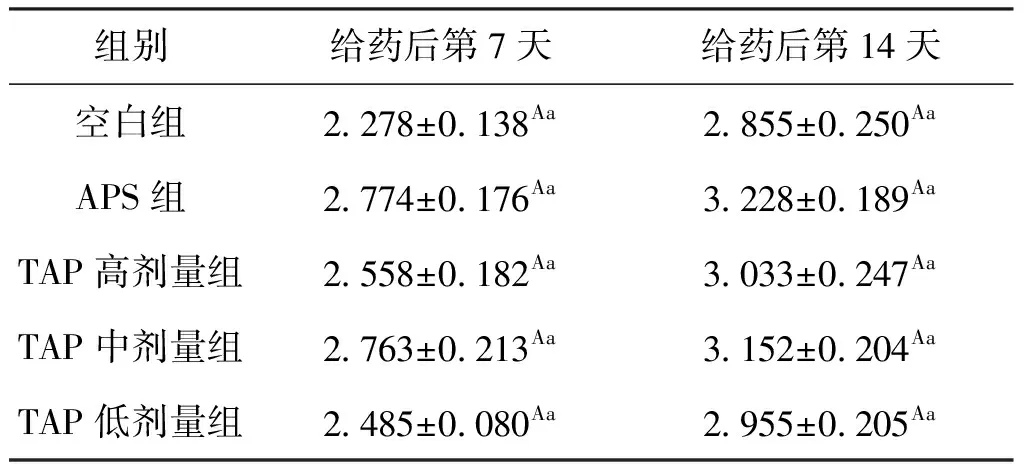
表2 TAP对鲫肝体比的影响Tab.2 Effect of TAP on hepatopancreas index of C.auratus %
2.2 TAP对鲫肝胰脏抗氧化功能的影响
给药7 d后,APS组鲫肝胰脏中的MDA含量比空白组降低了43.0%,TAP中剂量组MDA比空白组降低了35.9%,且二者相比差异不显著;给药14 d后,MDA含量在各组间差异均不显著,但TAP中剂量组的MDA比空白组降低了23.6%。结果表明,1% APS与0.5% TAP给药7 d后,可快速降低鲫肝胰脏内MDA水平,但随着继续给药,APS组MDA水平恢复正常水平,而TAP中剂量组MDA仅略有回升,仍然低于空白组,反映两种多糖的抗氧化机制可能存在差异,结果见表3。
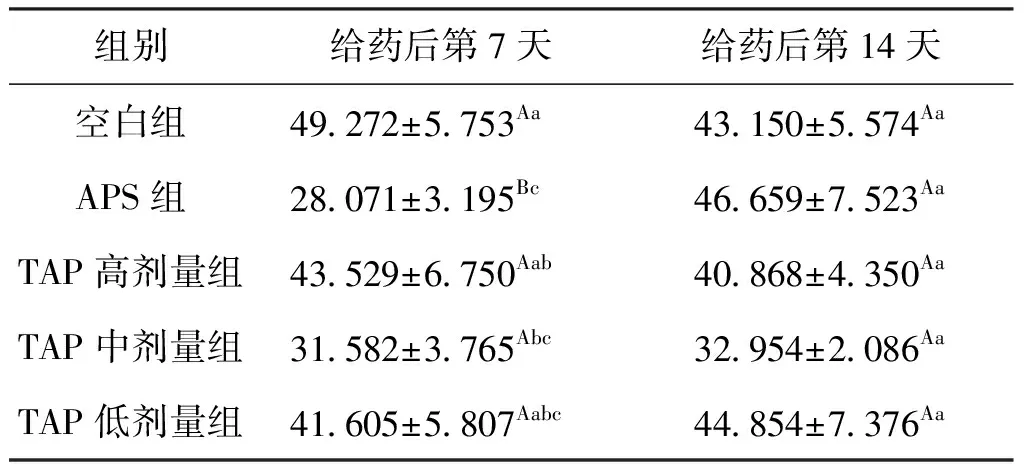
表3 TAP对鲫肝胰脏中MDA含量的影响Tab.3 Effect of TAP on MDA in hepatopancreas of C.auratus nmol/mg prot
用药7天后,各组与空白组相比CAT含量均有不同程度提高,APS使肝胰脏的CAT含量提高了14.93%,TAP高、中、低剂量组则分别提高了21.24%、26.31%(P>0.05)、5.74%;用药14 d后,APS组CAT含量提高了14.20%,TAP高、中剂量组则分别提高了34.61%、25.57%,高剂量TAP效果最好。结果表明,TAP的高、中浓度在给药期间能持续的提高鱼肝胰脏CAT水平,效果优于APS,结果见表4 。
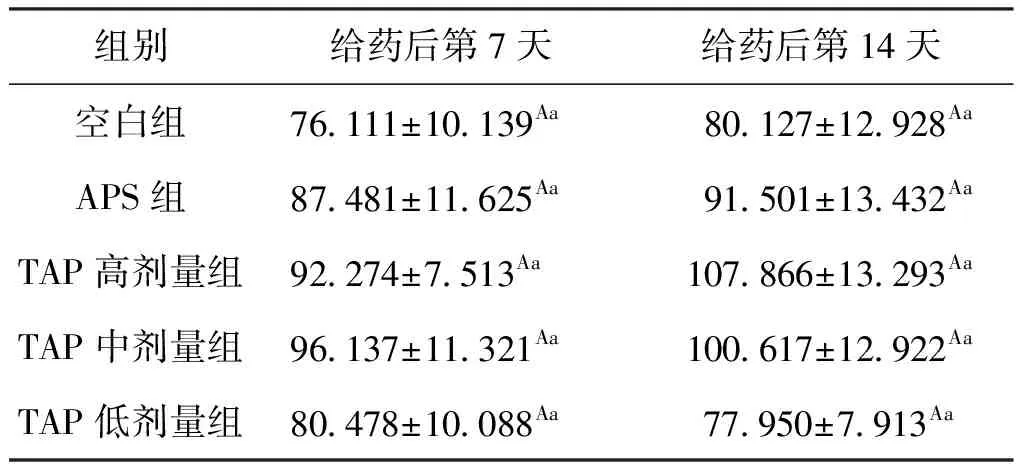
表4 TAP对鲫肝胰脏中CAT含量的影响Tab.4 Effect of TAP on CAT in hepatopancreas of C.auratus U/mgprot
给药7 d后,各组与空白组相比,SOD含量均无显著差异;TAP高剂量组的SOD含量比空白组提高了32.6%,同时比APS组提高了43.4%。给药14 d后,APS组的SOD含量高于第7天时的水平,提高了33.3%;与空白组相比,APS组SOD含量提高了26.81%,而TAP高剂量组提高了10.05%。结果表明,APS与TAP对肝胰脏的SOD水平均有促进其升高的趋势,但出现浓度高峰的时间不一致,提示二者对SOD的影响机制可能不同,结果见表5。
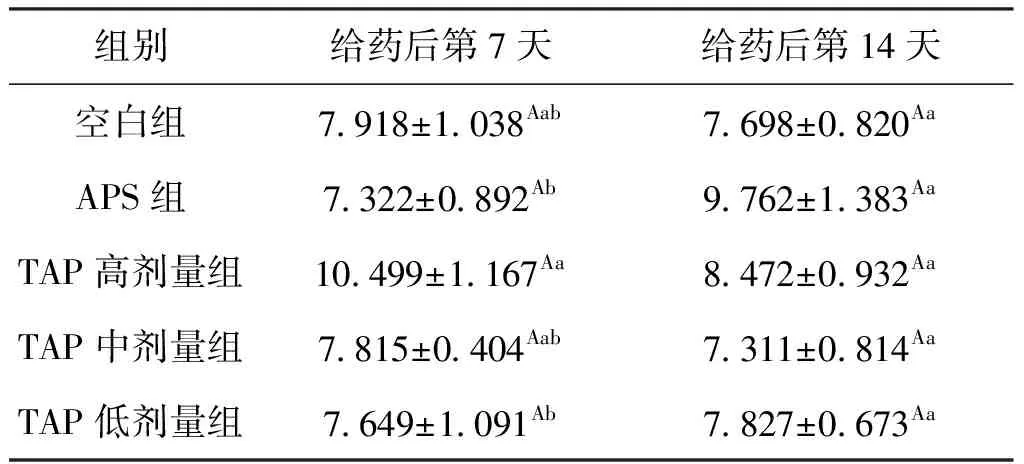
表5 TAP对鲫肝胰脏中SOD含量的影响Tab.5 Effect of TAP on SOD in hepatopancreas of C.auratus U/mgprot
2.3 TAP对鲫免疫功能的影响
鱼类的头肾既是造血器官也是免疫器官。给药7 d后,各组HKBI均有不同程度的升高;与空白组相比,APS组HKBI升高了18.1%,而TAP中剂量组的HKBI升高了27.3%。给药14 d后,APS组的HKBI比空白组升高了44.2%;TAP高、中、低剂量组HKBI则比空白组分别升高了19.4%、32.5%、38.8%。结果表明,连续给药14 d后1% APS和0.5%、0.1% TAP均可显著提高鲫的免疫器官指数,且TAP的免疫增强效果优于相同剂量APS,结果见表6。
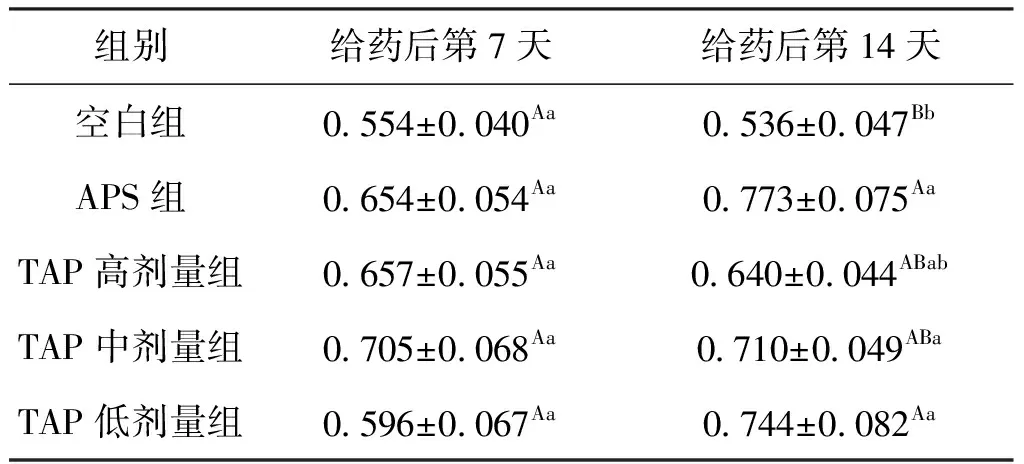
表6 TAP对鲫HKBI的影响Tab.6 Effect of TAP on HKBI of C.auratus %
鲫肝胰脏中的LZM是其重要的非特异性免疫指标。用药7 d后,各组LZM水平均高于空白组;与空白组相比,APS组LZM提高了19.8%;而TAP高剂量组LZM水平提高58.1%;中剂量TAP组提高了34.9%;低剂量TAP组提高了11.1%。给药14 d后,APS组的LZM含量与空白组仍然无显著差异,TAP高剂量组则比空白组提高了27.5%;TAP的中、低剂量组LZM水平均高于空白组和APS组,但组间差异均不显著。结果表明,1% APS对鲫肝胰脏的LZM含量没有显著影响,而0.5%~1% TAP对鲫非特异性免疫功能有持续的增强作用,结果见表7 。
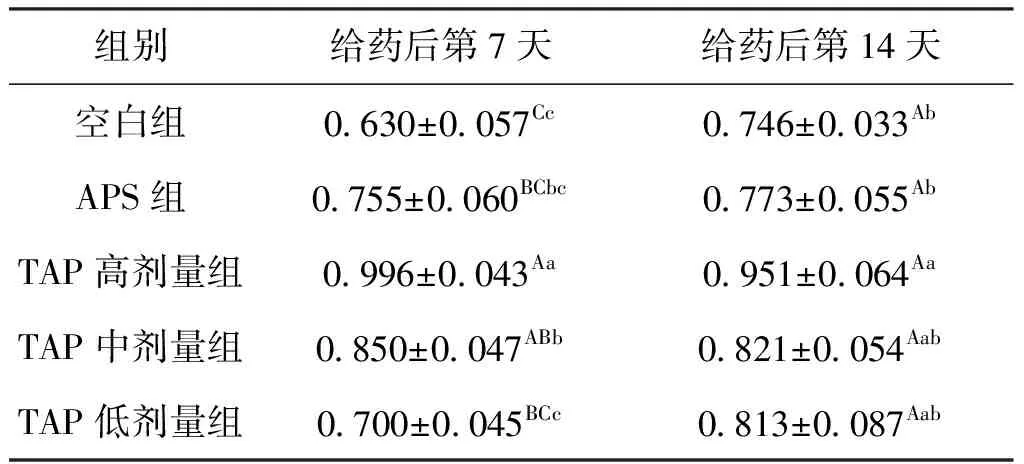
表7 TAP对鲫肝胰脏中LZM含量的影响Tab.7 Effect of TAP on LZM in hepatopancreas of C.auratus μg/mL
3 讨论
3.1 两种多糖对鲫生长性能的影响
据报道,APS能够改善齐口裂腹鱼[8]、罗非鱼的生长性能[2];APS在低剂量时可延缓斑马鱼细胞的凋亡,而高剂量有抑制生长的作用[9];但也有研究表明,APS对黄颡鱼的增重率、增长率、肌肉蛋白含量、饵料系数方面均无显著影响[10]。本试验结果也表明,1% 饲料添加量的APS对鲫的肥满度无显著影响,与上述结论相似;但相同剂量的TAP对鲫的肥满度有短期提高作用,揭示TAP短期内可能通过调节鲫的饲料利用率对鲫生长有一定促进作用。
肝胰脏是鱼类的可调节性贮脂器官,其重量反映出脂肪含量及营养状态,当饲料中的营养因子和抗氧化因子的影响了脂肪的代谢,其肝体比就会产生变化。本试验结果表明,两种多糖均对鲫的肝体比有提高的趋势,这可能与其能增强机体抗氧化作用有关。
3.2 多糖对鲫体内抗氧化能力的影响
鱼类在新陈代谢的过程中,体内活性氧自由基的产生和消除保持着一种动态平衡,过多的自由基会引起脂质过氧化反应,而MDA是脂过氧化的主要产物,具有细胞毒性。CAT和SOD组成的抗氧化酶系统可清除过多的自由基,减少脂质过氧化造成的细胞损伤。据报道,在齐口裂腹鱼的饲料中添加0.06%~0.08%的黄芪多糖,可使其血清中的 LZM、SOD活力显著高于未添加组[8]。研究表明,长期投喂1.2 g/kg黄芪多糖可显著提高黄颡鱼肝胰脏中 LZM 和 CAT 的活力,显著降低肝胰脏中NO的含量,但对肝胰脏中SOD的活力无显著影响[11]。可见APS对不同鱼类抗氧化酶的活性存在一定影响,这可能与APS的给药剂量与时间有关。本试验结果表明,APS可降低鲫肝胰脏中的MDA水平,这可能与APS能提高其CAT、SOD活性有关,且TAP的体内抗氧化效果优于相同剂量APS。
3.3 多糖对鲫免疫功能的影响
免疫器官指数可代表免疫器官的发育程度,与免疫力成正比关系,是反映免疫功能的重要指标。鱼类主要的免疫器官是由淋巴样组织构成的头肾,是免疫细胞及抗体产生的主要场所,因此常以头肾体指数来反映鱼类的免疫器官指数。本试验结果表明两种多糖均能显著提高鲫的头肾体指数,因而对其免疫功能有促进作用。
溶菌酶具有杀菌作用,是鱼类非特异性免疫系统的重要组成部分。据报道,在罗非鱼的基础饲料中连续56 d添加0.1%的云芝多糖时,其血清LZM的活性最高(P>0.05),添加量为0.25% 时则显著低于对照组[12]。同样的云芝多糖给药56 d,添加0.1% 到银鲫基础饲料后仅在14~21 d时对血液白细胞吞噬能力、LZM活性均有显著促进作用[13]。以上研究表明多糖的免疫调节作用与给药剂量和时间有关,揭示多糖对健康鱼类的非特异性免疫功能可能具有双相调节作用或容易引起耐受。在本试验中,APS与TAP均可提高HKBI,但APS对LZM的影响不显著,这可能与APS的给药剂量和时间有关;而1% TAP可使鲫肝胰脏中LZM水平在试验期间均显著高于空白组,且LZM的浓度随着给药时间延长也出现下降的趋势。此外,有研究表明,免疫活性多糖的主要成分是β-葡聚糖,而从担子菌中提取的β-葡聚糖大多是β-1,3葡聚糖结合而成,其侧链和残基的分布和数量可能影响其生物学[14]。TAP是从担子菌门的鸡枞菌提取出的多糖成分,其结构与APS不同,这可能是两种多糖对鲫肝脏LZM的影响存在较大差异的原因。
本研究结果表明,以TAP连续给药14 d对鲫肥满度和肝体比均无显著影响,而对鲫抗氧化和免疫功能有显著的促进作用,效果优于相同剂量的APS,具有开发成鱼用保健品的潜力。
