氮沉降增加情景下植物-土壤-微生物交互对自然生态系统土壤有机碳的调控研究进展
2019-01-18程淑兰方华军于光夏曹子铖
程淑兰,方华军,*,徐 梦,耿 静,何 舜,于光夏,曹子铖
1 中国科学院地理科学与资源研究所/生态系统观测与模拟重点实验室,北京 100101 2 中国科学院大学资源与环境学院,北京 100049
当前全球碳循环研究中的一个关键科学问题是“失踪碳汇”(missing C sink)的分布与驱动机制[1]。过去150年,人类活动导致大气氮沉降增加了3倍[2],显著提高了受氮限制的陆地生态系统碳储量,是正确解释“失踪碳汇”的重要途径[3]。然而,目前有关氮沉降驱动陆地生态系统固碳的效率以及时空格局还存在很大的不确定性,不同学者估计的“氮促碳汇”变化范围为16—400 kgC/kgN[4- 11]。相对于植被碳汇,有关氮沉降增加情景下土壤碳储量的演变方向也存在分歧,包括增加[12]、降低[13]和不变[14]等3种结论。增氮引起土壤碳汇增量变化范围为0—70 kgC/kgN[7, 15- 17],同样存在很大的不确定性。
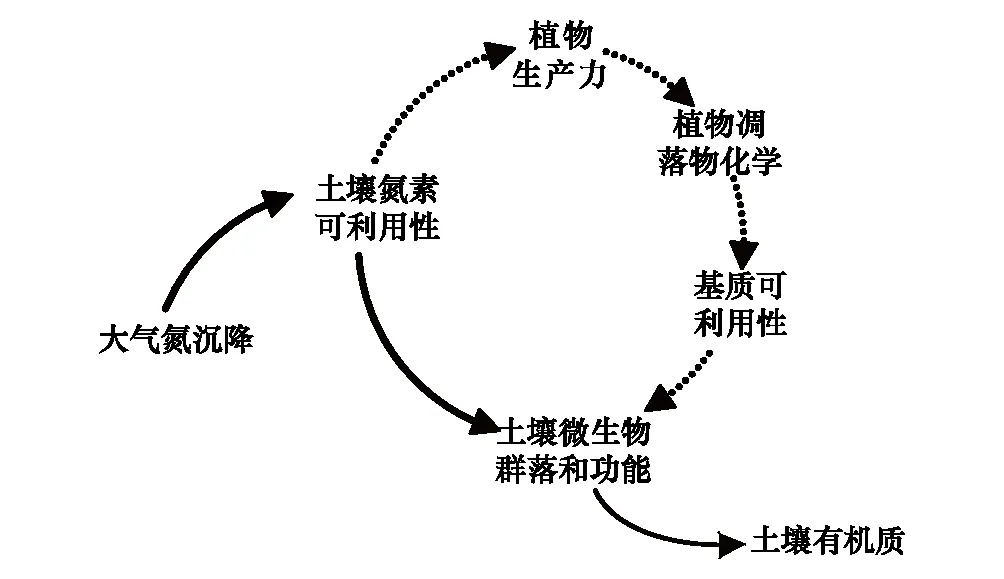
图1 氮沉降对自然生态系统植物和微生物群落的影响 Fig.1 Effects of nitrogen deposition on plant and microbial communities in natural terrestrial ecosystems实线为直接影响,虚线为间接影响
大气氮沉降输入提高生态系统氮素的可利用性,直接改变土壤微生物群落的数量和组成,抑制根系和凋落物分解的速率和程度[18],进而增加或降低土壤有机质(SOM)的储量和稳定性[19];另外,大气氮沉降输入提高植被生产力,改变凋落物产量和生化属性[20],通过底物可利用性来调节土壤微生物群落的结构和功能[21],间接影响SOM的累积速率(图1)。土壤有机碳(SOC)库对增氮的响应取决于生态系统类型[22]、土壤深度[23]、SOC组成[24]、施氮类型和剂量[25]等要素。就植物-土壤-微生物群落系统而言,增氮会自上而下改变SOC的周转过程与稳定性,理论上会产生以下4种影响:①外源性氮输入会改变植物同化碳的再分配,根际沉积碳的数量和质量的变化进而影响活跃微生物群落组成[26];②氮素有效性增加会抑制微生物胞外酶活性,降低微生物群落获取碳源的能力,提高其碳素利用效率[27];③氮素富集会改变微生物群落之间的交互作用和竞争关系,进而改变分解微生物群落的组成[28];④氮素富集会降低凋落物的分解速率和程度,增加类木质素化合物的氧化程度和稳定性,进而促进SOM的积累[29]。然而,上述4个科学假设缺乏系统的实验验证,其内在的微生物分子生态学机理未得到很好的理解,也没有被融入到当前主流的生物地球化学模型之中[30]。
过去30年,尽管自然陆地生态系统对大气氮沉降增加的响应研究得到了充分的重视,但是主要侧重于生态系统碳氮转化过程与土壤碳储量动态,缺乏植物-土壤-微生物群落的交互作用及其调控机制的研究,对SOC积累与损耗的微生物学机理研究也不够系统深入。探讨植物-土壤-微生物群落三者交互作用及其对土壤碳储量动态的调控作用,研究有助于完善陆地生态系统碳-氮循环耦合模型,深入认识SOC截存与损耗的机理,有效降低区域陆地“氮促碳汇”评估的不确定性,并可为陆地生态系统应对全球变化提供科学依据。鉴于此,本文以地下碳循环过程为主线,分别综述氮富集对植物地下碳分配、SOC激发效应、微生物群落碳代谢过程的影响,深入分析SOM化学稳定性与微生物群落动态的关系。论述过程中我们试图标注各个领域的薄弱环节,明确未来可能的研究重点,期望起到抛砖引玉的效果。
1 增氮对植物地下碳分配的影响
2 氮素有效性对SOC激发效应的影响
增加凋落物的输入会降低土壤碳库,该现象称为“激发效应”[47]。激发效应是由于外源底物的添加激活土壤中的微生物,促进它们对SOM的分解。根据激发方向可分为正激发和负激发效应,根据有无有机质参与可分为表观激发和真实激发[48]。当添加的外源底物远低于微生物量时,由于微生物自身内部代谢和周转导致CO2释放的过程,并不涉及SOM周转,因此表观激发对土壤碳库没有影响。但是当添加的底物剂量远高于微生物量时,微生物优先利用外源底物的能量,降低对SOM的分解,因此会出现负激发。随后,当外源底物的能量耗尽之后,促使被激活的微生物分解SOM来获取所需的养分和能量,从而产生正激发。基于全球培养实验的集成分析,发现正激发效应可促使有机质分解增加 3.8倍,而负激发效应导致SOM分解减少50%[49]。激发效应的产生机制主要包括:①共代谢理论[50];②底物偏好理论[51];③氮挖掘理论[52];④化学计量学理论[53]等。正激发产生机理以微生物激活作用、微生物竞争与演替、养分挖掘和化学计量学为主,而负激发产生机理以底物偏好利用和毒害作用为主。微生物的种类和大小[51]、植物类型和物候[54]、土壤动物对微生物的取食[55- 56],均会影响激发效应的强度。非生物因素如底物质量[57]、数量[58]和土壤属性(温度、水分、土层、养分状态)也会影响微生物群落对底物的利用,进而影响激发效应的强度和方向[59-62]。
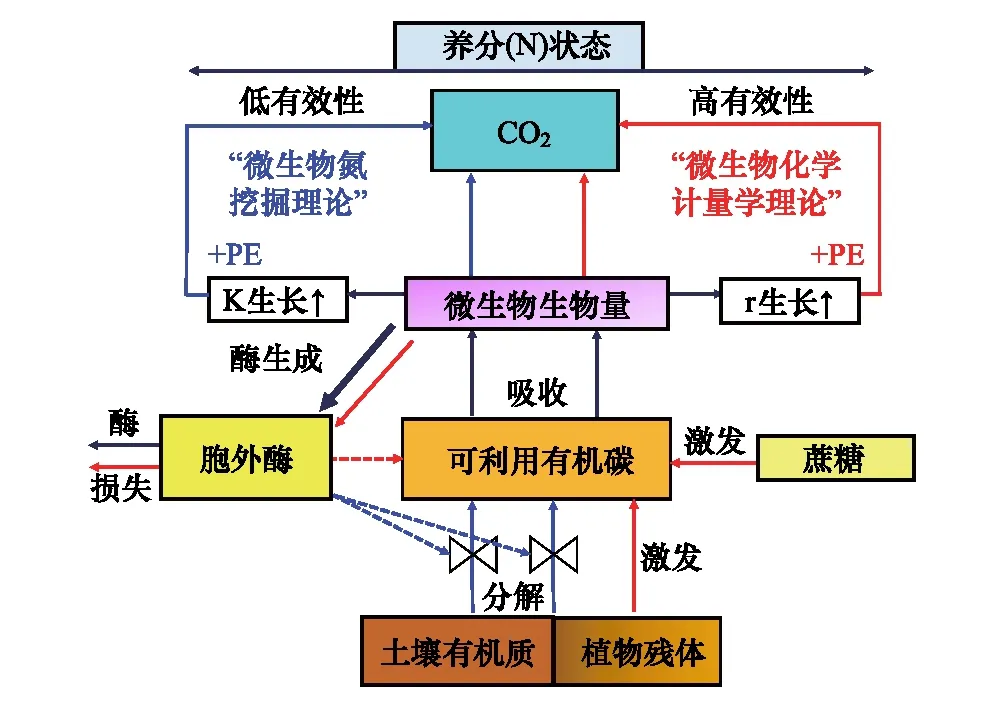
图2 养分挖掘理论与化学计量学理论关系[53] Fig.2 The relationship between nutrient mining theory and stoichiometry theory[53]PE: 激发效应priming effect

3 增氮对微生物群落碳代谢的影响
微生物碳利用效率(CUE)是指分配给微生物生长所用有机碳与微生物群落组成所吸收的有机碳之比,它是微生物群落碳代谢的一个重要的综合指标[67]。理论上,微生物CUE被热力学限制在0.88[68],即微生物吸收的碳用于生长的部分不会超过88%,因为微生物至少需要12%的碳用于呼吸产生能量以维持其自身的生物量。然而,由于环境条件的限制,微生物生长和生物量的维持需要超过其最小的能量投入,土壤微生物CUE未达到理论上的最大值[69]。土壤生源要素化学计量学、底物质量、微生物群落组成均会影响微生物CUE。在养分限制条件下(高碳/养分比),微生物被迫分配较多的碳和能量以获取所需养分,土壤呼吸CO2排放较高,导致微生物CUE较低[67, 69]。当外源性底物的C∶N∶P比与微生物生物量C∶N∶P比(60∶7∶1—42∶6∶1)差异较大时,微生物必须分配更多能量生产胞外酶来获取缺失元素以维持其生物量;同时更多同化碳被用于微生物呼吸,导致微生物CUE下降[67, 69]。因此,养分富集农田土壤的微生物(尤其是细菌)生长效率普遍高于受氮限制的森林和草地土壤[70]。此外,碳和养分的可利用性也会影响微生物生物量的周转[71]。微生物CUE和土壤微生物生物量周转时间可能受有机碳浓度和质量的影响。降解复杂化合物需要多种胞外酶,而酶的合成需要消耗大量的能量和氮,所以降解复杂底物的微生物CUE较低[72]。由于微生物通过不同的代谢路径来同化不同类型底物,导致单位质量的同化碳具有不同的呼吸速率[73]。因此,植物类型、微生物群落组成以及不同来源的凋落物也会间接地影响微生物CUE,真菌的CUE一般高于细菌[74]。目前测定微生物CUE的方法主要是基于13C标记底物,然而不同底物的微生物CUE差异巨大,该方法混淆了微生物碳利用效率与特定底物的利用效率[75]。最近,Spohn等[76]建立了基于18O-H2O标记技术计算微生物CUE的新方法,根据18O-DNA的增加量来计算土壤微生物的生长速率,再结合基础呼吸速率和微生物数量,可计算出微生物CUE和周转时间。
野外观测和模型模拟结果表明,施氮降低微生物活性、SOM分解和CO2排放[77- 80]。施氮可能通过改变微生物群落活性和组成[81- 82]以及底物的化学性质(如可分解性)来抑制土壤微生物呼吸,与微生物CUE、胞外酶活性以及微生物生物量的变化有关[27]。其理论假设是:(1) 氮限制条件下,增氮会导致更多的碳向微生物分配,而不是通过呼吸和胞外酶损失,导致微生物CUE增加,呼吸降低[69],这可能与微生物群落组成向高CUE转变有关[83]。(2)增氮抑制了降解复杂底物(如木质素)的氧化酶活性。例如,增氮降低了北美温带森林土壤酚氧化酶活性[22]、木质素降解相关的功能基因丰度和基因表达[84]。(3)增氮通过减少分解微生物的生物量而降低呼吸[33]。增氮导致土壤酸化,增加Ca2+、Mg2+等阳离子的溶解与流失,土壤溶液中Al3+浓度增加,对微生物产生毒害作用[85]。此外,土壤酸化也会抑制微生物胞外酶活性,金属离子的键合增加有机质中碳的稳定性,降低微生物获取碳的能力,进而降低微生物生物量和分解速率[86- 87]。尽管已经对氮素富集条件下土壤微生物碳的代谢过程和响应格局做了一些探讨,但是氮素类型和剂量如何差异性地影响微生物碳代谢功能还不得而知;由于缺乏合适的测定方法,氮沉降增加情景下自然陆地生态系统土壤微生物碳利用效率及微生物生物量的周转时间尚未得到准确的量化。
4 SOM化学稳定性与微生物群落动态的关系

有机质化学结构的变化也会影响土壤微生物的底物利用,进而影响微生物群落的活性和组成,反之亦然。长期施氮会显著降低森林、草地等自然生态系统土壤微生物生物量[34],抑制腐生菌分泌木质纤维素水解酶,降低腐生微生物群落获取碳源(如纤维素、半纤维素)的能力[93];此外,氮沉降输入还会抑制白腐菌产生酚氧化酶,进而抑制木质素溶解酶生成,促进耐分解有机质的积累[94]。利用功能基因定量PCR和高通量测序技术,Hassett等[95]和Edwards等[84]研究发现,施氮倾向于降低美国密歇根糖枫林土壤担子菌漆酶基因lcc的拷贝数(lcc编码酚氧化酶),改变真菌群落组成(担子菌OTUs/子囊菌OTUs增加),并且随着施氮时间的延长其抑制效应更加显著。同时,施氮倾向于降低凋落物分解速率和程度,增加类木质素化合物的氧化度和稳定性,进而促进SOM的积累[29]。潜在的生物化学机理是:高剂量的无机氮会抑制某些担子菌木质素水解酶的合成,以及编码酚氧化酶、锰过氧化物酶和木质素过氧化物酶的真菌基因转录下调,导致木质素降解下降[96]。木质纤维素水解酶基因转录下调会导致凋落物分解下降和SOM积累[97]。总体上,过去对氮沉降增加情景下森林土壤真菌功能基因表达和氧化酶活性演变研究较为深入,但是有关草地土壤细菌和真菌群落组成的变化如何代谢有机底物的机制仍不清楚,土壤微生物群落组成、SOM化学结构的变化与SOC积累之间的耦联关系尚不清晰。
5 存在问题与展望
目前关于外源性氮输入对自然生态系统土壤碳动态的影响已开展了一些探索,但总体上还不够深入,存在许多问题亟待解决。首先,植物根系碳输入对增氮的响应呈现抛物线变化,取决于生态系统氮素有效性、施氮的类型与持续时间。总的来说,施氮降低根系的生长与周转,但是根际沉积物对增氮的响应及其固碳效应十分复杂,与植被-菌根类型、生态化学计量学等要素有关,未形成统一的研究结论。其次,关于激发效应的3个阶段、4种驱动机制及影响因素分析较为深入,认识到养分尤其是氮素强烈影响激发效应的方向及强度,但是氮素类型、土层深度及其交互作用对激发效应的贡献并不清楚。再次,微生物CUE是微生物群落碳代谢的关键表征,受底物生态化学计量学驱动,能够很好地解释氮沉降增加情景下土壤碳的积累与损耗过程;由于缺乏适宜的测定方法,目前还难以准确量化土壤微生物CUE及微生物生物量的周转时间。最后,SOM化学质量与土壤微生物群落活性、组成之间存在复杂的交互作用,增氮显著抑制贫氮生态系统土壤真菌群落及其胞外酶活性,对细菌群落组成的影响结论多样,有关微生物群落代谢底物、SOC积累的生物化学机制尚不清楚。

图3 氮沉降增加情景下植物-土壤-微生物交互作用对土壤有机碳动态的调控机制Fig.3 Mechanisms responsible for the regulation of plant-soil-microbe interactions to soil organic carbon dynamics under elevated nitrogen deposition
6 结论
外源性氮输入通过影响微生物群落活性和组成直接影响SOM分解,或通过改变底物数量和质量间接影响SOC的分解和累积过程。总体上,增氮倾向于促进地上生物量,抑制地下碳分配和根系周转。氮素有效性强烈影响SOM激发效应的方向和强度,取决于微生物生长的r-K策略和生态化学计量学特征。微生物CUE是衡量微生物群落碳代谢的重要表征,能够很好地解释土壤碳的积累与损耗过程,目前缺乏准确测定CUE的方法,稳定性碳氧同位素示踪是解决该问题较为理想的手段。SOM化学结构与微生物群落活性、组成紧密耦合在一起,增氮抑制真菌群落及氧化酶活性,对细菌群落组成的影响尚未形成定论,需要将有机质化学与分子生物学、宏基因组学方法紧密结合起来,深入地揭示增氮引起土壤碳积累与损耗的生物化学与微生物学机理。
