不同水分条件对3种蕨类植物配子体形态发育的影响
2019-01-14高晓雯周施雨卞竹箐王清华戴锡玲
高晓雯, 周施雨, 卞竹箐, 王清华, 戴锡玲
(上海师范大学 生命科学学院 植物种质资源开发协同创新中心,上海 200234)
0 引 言
我国蕨类植物资源丰富,约有2600种,占全世界蕨类总数的1/5.STOKEY[1-2]研究了多种蕨类植物配子体的发育过程,证实了其中的孢子萌发类型、配子体发育类型及成熟原叶体形态[3-5]在探讨蕨类科、属分类和系统演化方面具有一定的价值.但是,相对于种子植物,有些蕨类植物配子体性状表现不稳定,易受环境条件影响而变化,例如,环境光的颜色、光照强度和温度对蕨类植物孢子萌发、配子体生长、假根生长及两性配子体的形成具有一定的影响[6-10].而水分条件对配子体形态发育的影响未见报道.
利用液体培养基培养乌毛蕨(Blechnumorientale)孢子[11],其孢子萌发时间较混合土培的明显缩短,丝状体长度更长,假根细长[12].用含水量较大的Knop′s琼脂培养基和混合土对普通针毛蕨(Macrothelypteristorresiana)孢子进行培养[13],土培配子体在接种后35 d可观察到精子器,而在含水量较大的Knop′s琼脂培养基上的配子体产生精子器则需要60 d左右.采用MS(Murashige and Skoog)培养基、混合土和吸水滤纸分别培养傅氏凤尾蕨(Pterisfaurieisp.)孢子[14],发现其配子体的形态与培养方式有关,MS培养基与土培的配子体假根粗短.以上研究结果表明,不同的水分条件可能是影响蕨类植物配子体形态发育的重要因素之一,对蕨类植物配子体形态中丝状体形态、假根长短及性器官分化等存在一定影响.
本文依据PPG-1(2016)蕨类植物分类系统[15],选取薄囊蕨类(Polypodiidae)中莎草蕨目(Schizaeales)海金沙科(Lygodiaceae)海金沙[Lygodiumjaponicum(Thunb.) Sw.],水龙骨目(Polypodiales)中真水龙骨类(Eupolypods):真水龙骨类I(Eupolypods I)凤尾蕨科(Pteridaceae)水蕨[Ceratopteristhalictroides (L.) Brongn],真水龙骨类II(Eupolypods II)乌毛蕨科(Blechnaceae)狗脊[Woodwardiajaponica(L.f.) Sm.]3种不同演化地位的蕨类植物,其中海金沙、水蕨配子体的形态发育已有报道[16-17].利用改良Knop′s琼脂培养基,在其余环境条件完全相同的条件下,设置3组不同的水分条件,对3种蕨类植物配子体进行培养;利用光学显微镜对3种蕨类的孢子萌发、丝状体、片状体、原叶体、性器官和假根等形态特征进行详细观察,旨在进一步明确蕨类植物配子体形态发育过程中不受水分条件影响的稳定性状,为蕨类植物配子体发育和分类演化研究提供基础资料.
1 材料和方法
1.1 材 料
海金沙(L.japonicum)于2013年7月采自浙江宁波;水蕨(C.thalictroides)于2013年9月采自上海师范大学奉贤校区植物园;狗脊(W.japonica)于2013年8月采自浙江西天目山.将带有成熟孢子的3种蕨类植物叶片置于洁净纸袋中,放于干燥通风处,使孢子自然散落,约1周后将孢子去杂并收集于硫酸纸袋中.
1.2 方 法
在超净工作台上将5%(质量分数)次氯酸钠溶液加入放有适量孢子的离心管中充分振荡摇匀,浸泡5 min后离心(2000 r·s-1,3 min).去上清液,加入无菌水,振荡离心.重复3次.在离心管中加入无菌水,配制孢子悬浮液.用胶头滴管吸取1 mL孢子悬浮液,均匀涂布接种在平板培养基上(涂布密度小于100 mL-1),再按照无水、加入1管水(约2 mL)和2管水(约4 mL)分为3组,分别标记为组1、组2、组3,置于ZGX-300C型人工培养箱(浙江钱江仪器设备有限公司制造)中培养,全天光照,光照强度约为2000 lx,温度为(25±2) ℃,湿度为60%~75%.
定期利用Nikon SMZ1500型光学显微镜观察并记录、照相.实验重复3次,文中数据为3次观察的平均值,用SPSS17.0软件进行单因素方差分析.
2 结果与分析
2.1 不同水分条件下3种蕨类植物配子体的形态发育
2.1.1 海金沙配子体的形态发育
海金沙孢子接种后,组1、组2、组3的孢子萌发时间分别为16,14,11 d左右.海金沙孢子萌发类型为密穗蕨型(Anemia type)[18].
孢子萌发后形成丝状体.比较了不同水分条件下海金沙配子体的发育形态,如图1所示.由图1可知:组1的丝状体大多为单列细胞,长度为50~65 μm;组2和组3的丝状体则具有单列细胞和双列细胞两种形态,且丝状体长度为50~100 μm,明显大于组1的长度.丝状体顶端细胞经过多次纵裂,向两侧扩展,呈分散生长形成片状体,片状体进一步发育成心脏型原叶体,海金沙原叶体发育类型为铁线蕨型(Adiantum type)[18].
当原叶体上产生性器官,表明原叶体已成熟.3个组都出现了雌性原叶体和雄性原叶体,雄性原叶体上精子器出现的时刻都早于雌性原叶体上颈卵器出现的时刻,两种原叶体形态具有较大差异.海金沙接种约4周后,组3配子体首先出现精子器,约2 d后其余两组也相继出现精子器.精子器混生于假根丛中、两翼下部及近中肋,近球形,直径约为50~60 μm.3组产生的精子器的形态、大小无差异,且雄性原叶体的数量比3组雌性原叶体产生的数量多.海金沙接种9周左右后,组3最先出现颈卵器,颈卵器顶面观为铜钱状,且组3中雌性原叶体的数量多于组2和组1的.
海金沙配子体发育过程中,组1次生假根,始终粗短;组3次生假根细长;组2次生假根的长度介于组1和组3的长度之间.每组选取10个样本进行测量,计算长度平均值,对组2和组1、组3和组1的次生假根长度进行显著性分析比较,结果如表1所示,组3、组2的次生假根长度与组1的具有极显著差异性(p<0.01).
2.1.2 水蕨配子体的形态发育
水蕨孢子接种后,组1、组2和组3的孢子萌发时间分别为7,6,5 d左右,水蕨孢子萌发类型为书带蕨型(Vittaria type)[18].
接种9 d后,3个组丝状体发育大小不同.比较了不同水分条件下水蕨配子体的发育形态,如图2所示.由图2可知,组2和组3的丝状体长度明显大于组1的,组1的丝状体长度为(65.6±8.3) μm,组2的丝状体长度为(114.7±38.6) μm,组3的丝状体长度为(170.9±11.2) μm,丝状体形状均呈长条形.对组2和组1、组3和组1的丝状体长度进行了显著性分析比较,如上述结果所示,组3、组2丝状体长度与组1具有极显著差异性(p<0.01).
接种约12 d后,水蕨配子体进入片状体阶段,随后发育成略不对称的心脏型原叶体.水蕨原叶体发育类型为水蕨型(Ceratopteris type)[18].组1大多为雄性配子体,呈长条形;组2、组3大多为两性配子体或雌性配子体.
水蕨接种约4周后,组3首先出现精子器,约2 d后其余两组也相继出现精子器.精子器的出现同样早于颈卵器.精子器大多着生于原叶体边缘;接种5周后,组3首先出现颈卵器,多发生于原叶体生长点下方的中脉范围内.
组1的次生假根总体较短;组2的次生假根较长,组3的次生假根最长.对组2和组1、组3和组1的次生假根长度进行显著性分析比较,由表1可知:组3、组2的次生假根长度与组1的具有极显著差异性(p<0.01).

图1 不同水分条件下海金沙配子体发育形态比较
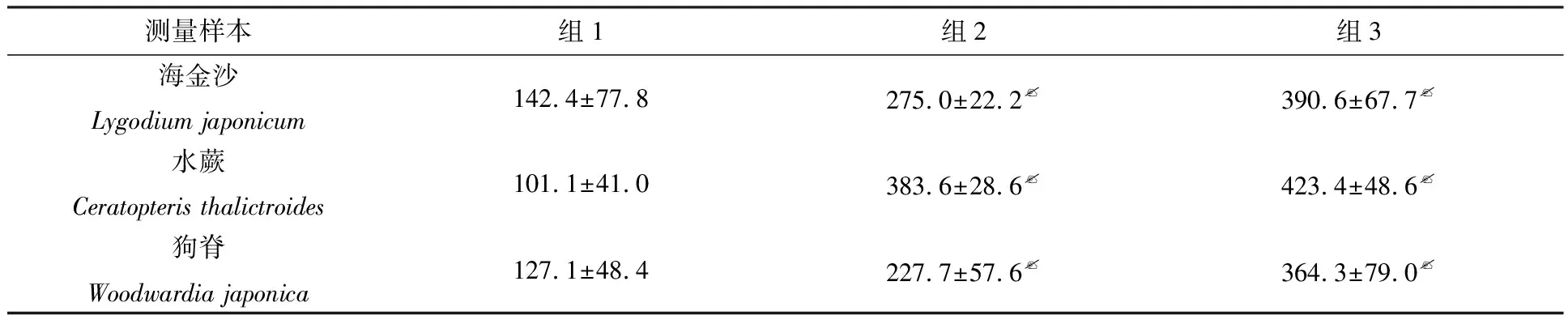
表1 不同水分条件下海金沙、水蕨和狗脊配子体次生假根长度比较 μm
注:p<0.01代表具有极显著差异性

图2 不同水分条件下水蕨配子体形态发育比较
2.1.3 狗脊配子体的形态发育
狗脊孢子接种后组1、组2、组3的孢子萌发时间分别为18,14,10 d左右,孢子壁从裂缝处裂开,长出初生假根以及含有叶绿素的原叶体原始细胞.狗脊孢子萌发类型为书带蕨型(Vittaria type)[18].
原叶体原始细胞横裂均形成单列丝状体.比较了不同水分条件下狗脊配子体的发育形态,如图3所示.由图3可知,组1、组2丝状体长度约为50~80 μm;组3丝状体最长,为85~110 μm.丝状体继续发育,大多形成倒梯形片状体.片状体发育形成的原叶体基部微收,边缘不平滑.在组3中明显可观察到:相隔几个细胞便有一个细胞凸出,单细胞棒状毛状体长在突出细胞的顶端.组1大部分成熟原叶体展开时宽约800~1200 μm,组2的宽约1000~1500 μm,组3的宽约1000~1800 μm.狗脊原叶体发育类型为三叉蕨型(Aspidium type)[18].
狗脊配子体为两性配子体.接种约6周后,组3首先出现精子器,约4 d后其余2组也相继出现精子器.接种7周左右,组3首先出现颈卵器.
在狗脊整个配子体发育过程中,组1的次生假根粗短,组3的细长,组2的介于两者之间.对组2和组1、组3和组1的次生假根长度进行了显著性分析比较,由表1可知:组3、组2的次生假根长度与组1的都具有极显著差异性(p<0.01).当配子体生长到成熟原叶体时期,组1的次生假根直径约为20 μm,组2的约为17 μm,组3的与组2的较为接近.
各组成熟原叶体形状没有差别,但在水分充足时,原叶体边缘毛状体下方细胞分裂,毛状体更突出,组1产生毛状体数量最少,长度为(8.9±0.5) μm,一个原叶体约有5~15个毛状体.组2的毛状体长度为(18.3±0.9) μm,一个原叶体约有10~23个毛状体.组3产生毛状体数量最多,毛状体长度为(20.3±1.1) μm,一个原叶体约有15~30个毛状体,发育最发达.对3个组所测数据进行显著性分析,结果显示,组3、组2的毛状体长度与组1的具有极显著差异性(p<0.01),说明水分条件对毛状体长度有影响.

图3 不同水分条件下狗脊配子体形态发育比较
2.2 不同水分条件对3种蕨类植物配子体形态发育的影响
通过以上对比观察,发现不同水分条件对3种蕨类植物配子体形态发育的影响表现在以下几个方面:
1) 对孢子萌发速率的影响:海金沙、水蕨和狗脊的孢子萌发速率不尽相同,但均在水分最为充足时(组3)萌发最早,而在没有水分时(组1)萌发最慢.
2) 对丝状体长度的影响:在水分充足的条件下,3种蕨类植物形成的丝状体长度最长,且海金沙在水分充足时易出现多列细胞的丝状体.
3) 对配子体性别表现的影响:在缺水或水分不足条件下,3种蕨类容易形成雄性配子体,而在水分充足条件下,形成雌配子体的概率较大.
4) 对假根和毛状体的影响:水分充足时,3种蕨类的配子体上次生假根更长,毛状体更发达且数量更多;缺水时配子体上次生假根较短,毛状体数量较少.
3 讨 论
3种蕨类的孢子萌发类型及配子体发育类型不受水分条件的影响[16-17].在水分充足时,海金沙、狗脊和水蕨孢子萌发速率明显较快,这与乌蕨(Stenolomachusanum)的情况相同[11-12];狗脊和水蕨的丝状体长度更长,这与金毛裸蕨(Clematischrysocoma)、中华桫椤(Alsophilacostularis)和扇蕨(Neocheiropterispalmatopedata)的情况相同[7-8,10,16-17,19].另外,在海金沙组中还出现了进一步发育的双列细胞丝状体.在水分充足时,狗脊配子体产生毛状体数量更多、发育更好,即毛状体数量及发育程度会受水分条件影响[20],但毛状体的形态、结构相对稳定[21].3种蕨类植物配子体的次生假根生长受水分条件影响较大[22],在水分不足的情况下,蕨类配子体次生假根基部膨大、末端分叉,而本实验结果表明:蕨类配子体假根在水分充足时较长,在缺水时较短.观察海金沙和水蕨的配子体发育发现:经历不适条件或环境胁迫时,蕨类植物多发育为雄配子体[19,23];而在适宜条件下易发育形成雌性配子体或两性配子体[24].这样使蕨类植物在逆境条件下趋于产生大量精子,提高受精率,有利于物种的生存[8,24].
4 结 论
通过设置3个不同的水分梯度并控制其他环境影响条件,研究了海金沙、水蕨和狗脊3种不同演化地位的蕨类植物的配子体形态发育过程.结果表明:不同水分条件对不同演化地位的蕨类植物配子体发育形态都具有一定影响.在水分充足时,孢子萌发速率更快,丝状体长度更长,性器官分化形成雌性配子体的概率更大,次生假根更长,毛状体更发达且数量更多;在水分不足时,孢子萌发速率较慢,丝状体较短,形成雄性配子体的概率更大,次生假根较短,毛状体数量较少.若将上述特征作为蕨类植物分类鉴定的依据,需要严格控制其外部环境.孢子萌发类型、配子体发育类型、成熟原叶体形态及毛状体形态结构不受水分条件的影响,是稳定的配子体特征性状,可以作为蕨类植物分类演化研究的依据.
