长角天牛属部分种类DNA条形码及系统发育的研究
2019-01-09禹海鑫孙民琴郭骁驹杨晓军安榆林
禹海鑫, 蔡 波, 孙民琴, 郭骁驹, 杨晓军, 安榆林
(1.南通出入境检验检疫局,江苏南通 226004; 2.海南出入境检验检疫局热带植物隔离检疫中心,海南海口 570311;3.江苏出入境检验检疫局动植物与食品检测中心,江苏南京 210009)
沟胫天牛亚科(Laniinae)隶属于鞘翅目(Cleoptera)叶甲总科(Chrysomiloidea)天牛科(Cerambycidae),是天牛科中种类最多的一个亚科。其中,长角天牛属(Acanthocinus)天牛是该亚科内极为重要的一类林木蛀干害虫,可危害鱼鳞松、油松、马尾松、红松、山杨、核桃、栎属等多种针、阔叶树种。该属天牛主要以幼虫钻蛀树木的木质部、韧皮部和边材组织为害,常使树势衰弱,危害严重时还会导致树木整株死亡,给农林业生产造成极大的经济损失[1]。截至目前,该属昆虫在全世界范围内已被描述的种类约有14种,广泛分布于欧洲、非洲北部、俄罗斯、日本、朝鲜、中国台湾等国家和地区。我国已报道的长角天牛属昆虫种类约有5种,包括白带长角天牛(A.carinulatus)、大灰长角天牛(A.aedilis)、小灰长角天牛(A.griseus)、黑带长角天牛(A.stillatus)和台湾长角天牛(A.gundaiensis)[2-3]。其中,白带长角天牛被2007年公布的《进境植物检疫性有害生物名录》列为我国进境植物检疫性有害生物。
近年来,随着我国进出口贸易的迅猛发展,原木、板材及木质包装的进出口总量也与日俱增,这些货物中极易携带大量天牛、小蠹等有害生物,对我国口岸生物安全造成极大威胁。我国口岸曾多次在进境原木及木质包装中截获到长角天牛属昆虫。例如,2013年深圳口岸检疫人员从进境罗马尼亚云杉原木中截获了检疫性害虫——白带长角天牛[4];2016年,深圳蛇口口岸检疫人员在进境美国长叶松中首次截获到了结节长角天牛(A.nodosus)[5];2016年,江苏常州口岸检疫人员在进境美国白松原木中首次截获到了斜带长角天牛(A.oblisquus)[6]。
面对如此严峻的生物入侵形势,只有对有害生物进行快速、精确的鉴定才可以采取针对性的措施有效阻止其入侵我国。目前,各口岸主要采用传统的昆虫种类鉴定方法,即依据形态特征,同时参照昆虫分类检索表的方法对长角天牛属天牛进行分类鉴定。但在口岸检疫过程中,由于截获的天牛多以卵、幼虫、蛹或肢体残缺成虫的虫态出现,利用形态学特征进行鉴定相当困难,很多天牛标本就无法鉴定到种[7]。因此,迫切须要探索新的鉴定技术来弥补传统鉴定方法的不足。截至目前,利用分子生物学手段研究物种鉴定、系统发育已经成为昆虫学科的研究热点。其中,由于线粒体细胞色素C氧化酶亚基Ⅰ基因(mtDNACOⅠ)结构相对保守,且种间差异比较大,因此,利用基于COⅠ基因序列片段的DNA条形码技术进行近缘种间鉴定及系统发育的研究已经得到了更为成熟的应用[8]。Stauffer利用该DNA条形码技术对欧洲7种齿小蠹进行了快速准确的鉴定[9]。禹海鑫等应用该技术实现了白条天牛属下13个种类准确高效的鉴定[10]。邬颖等也通过该技术实现了光肩星天牛幼虫种类的快速鉴定[11]。郑丝竹深入研究了天牛科基因条形码的构建方法及分子快速鉴定技术,为探索天牛种类的分子鉴定方法提供了极有价值的参考[12]。
白带长角天牛作为重要的检疫性有害生物,对其及所在属内多个种类天牛进行快速鉴定具有重要意义。本研究收集了5种口岸截获的长角天牛样本,对其COⅠ片段进行扩增和测序,并与GenBank中下载的9条COⅠ序列进行比对,分析这些序列的碱基多样性、遗传距离和系统进化关系,以期获得1种快速鉴定长角天牛种类的分子方法,为今后农林部门及进出境植物检疫部门对长角天牛属昆虫的快速准确鉴定提供依据[13]。
1 材料与方法
1.1 试验样本
本试验所用标本由南通出入境检验检疫局有害生物检疫实验室提供,包括小灰长角天牛(A.griseus)、大灰长角天牛(A.aedilis)、斜带长角天牛、A.princeps、白带长角天牛5种长角天牛,所有天牛标本均由国内天牛专家安榆林研究员鉴定复核。标本来源与采集时间见表1。另外,从GenBank网站获得小灰长角天牛、大灰长角天牛、A.reticulatus、Acanthocinussp. CA14_3.01 4种长角天牛,及作为外群的棕榈象甲(Rhynchophoruspalmarum)的共9条COⅠ序列(表2),用于比对分析。其中,白带长角天牛是我国进境植物检疫性昆虫,其他长角天牛均是其同属近似种。
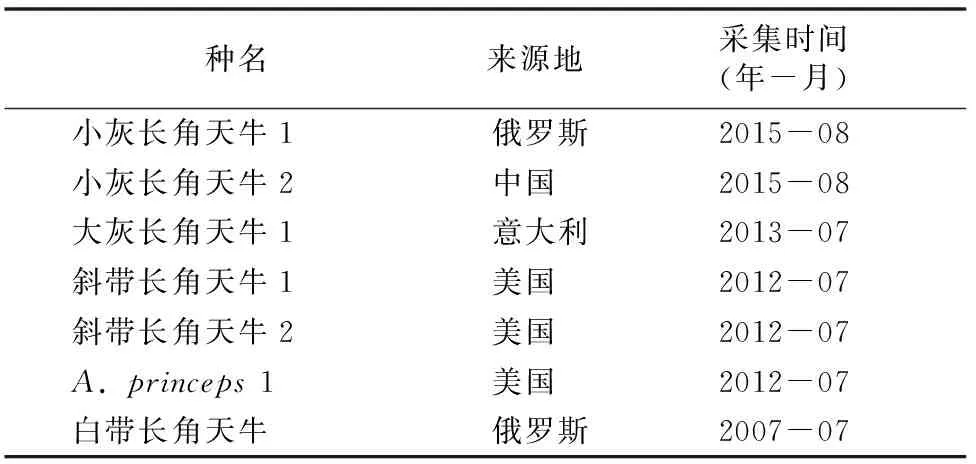
表1 供试标本的来源及采集时间

表2 本研究使用的GenBank下载的COⅠ基因序列信息
1.2 样品组织基因组DNA提取
用GenMagBio动物细胞组织/细胞基因组DNA磁珠提取试剂盒(北京金麦格生物技术有限公司)提取上述各个样品组织的基因组DNA。提取步骤如下:剪取适当大小的经乙醇浸泡的各天牛样品肌肉组织(应少于30 mg),用双蒸水冲洗干净后,装入 1.5 mL 离心管中并置于MM400球磨仪(德国retsch公司提供)中振荡研磨60 s(30次/s),再将磨碎的组织离心10 min(转速为12 000 r/min)。离心后加入180 μL裂解缓冲液及20 μL蛋白酶K,漩涡振荡,重悬已经预处理的组织样品,然后放入55 ℃水浴锅中温浴1~3 h,直至组织样品完全裂解消失。加200 μL无水乙醇、200 μL Binding Buffer、20 μL磁珠,磁珠用于吸附组织中的基因组DNA。加入 500 μL Wash Buffer去除杂质后,再加入20 μL Elution Buffer,静置10 min后洗脱磁珠,便可获得样品组织的基因组DNA溶液[14-15]。
1.3 COⅠ片段扩增与测序
本试验PCR扩增反应采用巢氏PCR,在ProFlexTMPCR仪(美国ABI公司提供)上进行。反应采用25 μL体系,包括1.0 μL基因组DNA模板、2.0 μL MgCl2(25.0 mmol/L)、2.5 μL 10×buffer(不含Mg2+)、1.0 μL dNTPs(2.5 mmol/L)、0.4 μL rTaqDNA聚合酶(5.0 U/μL,购自TaKaRa公司),各0.5 μL上、下游引物(10.0 μmol/L,由南京金斯瑞生物科技有限公司合成),再加灭菌水补至总体积为25 μL。PCR扩增反应条件如下:94 ℃ 5 min;94 ℃ 40 s,55 ℃ 30 s,72 ℃ 1 min,36个循环;72 ℃ 10 min。反应完毕后将产物送至南京金斯瑞生物科技有限公司进行测序分析[10,12]。巢式PCR第1轮反应引物为F-1、R-1,第2轮PCR引物为F-2、R-2[12],各引物序列信息如表3所示。本研究采用巢式PCR,既降低了扩增出非目的基因条带的概率,又增强了该PCR检测的可信度和灵敏度[12]。

表3 巢式PCR所用引物序列
1.4 序列处理及分析
将各样品COⅠ序列测序结果导入DNAStar中的SeqMan分析软件中进行拼接与手工校正[16]。利用美国国立生物技术信息中心(National Center for Biotechnology Information,简称NCBI)的Blast工具进行序列相似性搜索,以确定序列的方向和可信度。再将所测序列以及从GenBank下载的8条长角天牛序列一同载入Clustal X 1.83软件进行比对,输出格式为FASTA[17]。最后将所有比对结果导入MEGA 5.20软件中[18]计算各种类间的遗传距离、转换和颠换值及其比值(R)、变异位点(variable sites,简称V)及保守位点(conserved sites,简称C)等数值[13],同时,利用邻接法(neighbor-joining,简称NJ),选取Kimura 2-parameter遗传距离模型,建立系统发育树。
2 结果与分析
2.1 DNA凝胶电泳结果
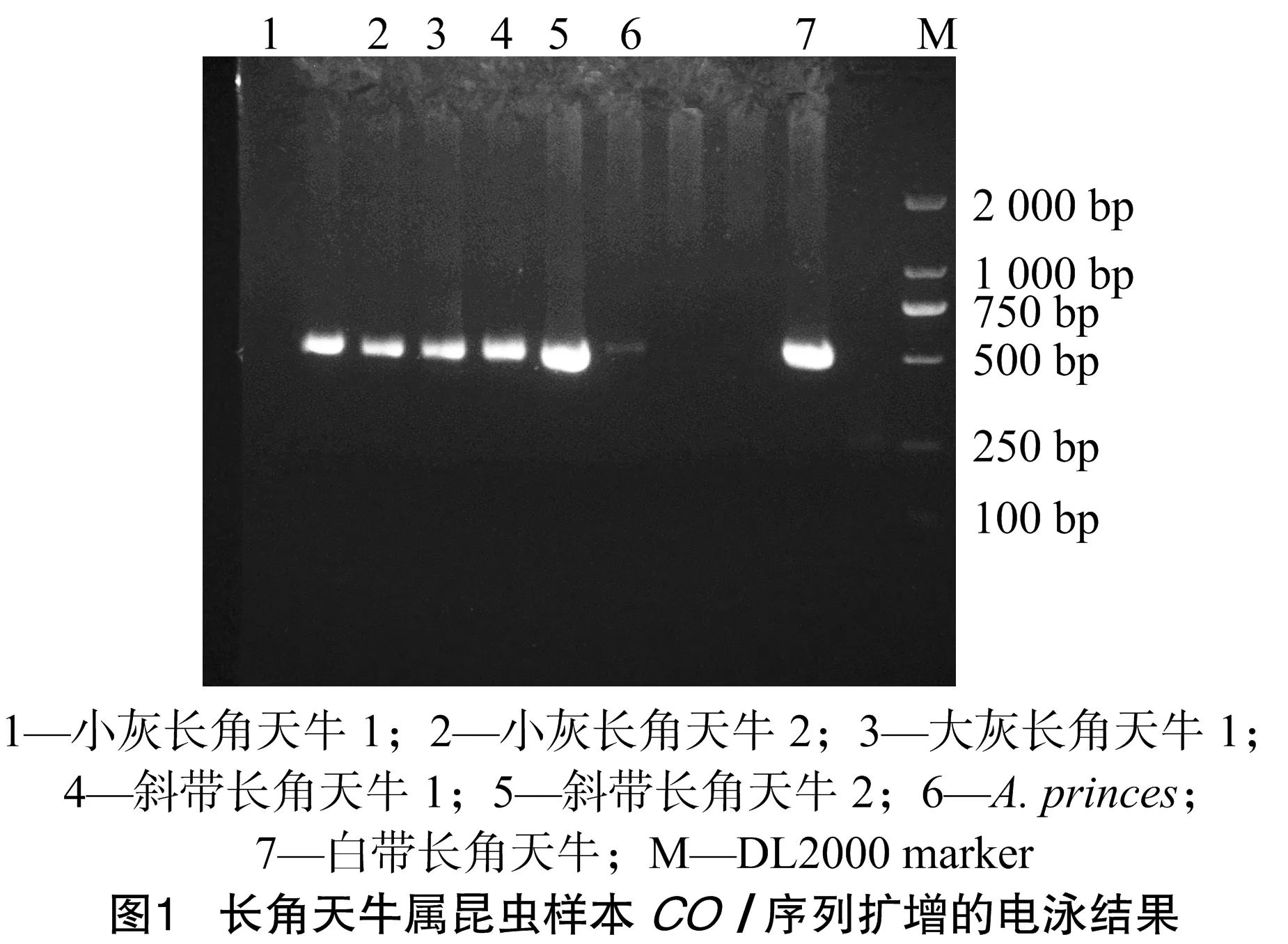
本试验对5种长角天牛共7个样本的基因组DNA进行COⅠ基因片段巢式PCR扩增,由图1的电泳结果可以看出,7个样本在525 bp处均有清晰且特异性良好的目的条带,可满足后续基因测序的需要。
2.2 长角天牛COⅠ序列解析
2.2.1COⅠ基因序列组成和变异特征 将测序和下载的各条序列导入MEGA 5.20软件中,剪切成同等长度片段(434 bp)[19]。结果发现,保守位点(C)、变异位点(V)、自裔位点(S)和简约信息(Pi)位点分别有199、235、56、179个。另外,统计所有位点的碱基平均含量发现,A平均含量为 29.8%,T为34.1%,G为14.4%,C为21.7%[12]。其中,A、T的含量相近,且A+T含量高达63.9%,明显高于G+C含量(36.1%),表现出明显的A+T碱基偏嗜,这也基本符合昆虫线粒体基因碱基组成的基本规律[19-20]。
2.2.2 碱基替换规律分析 用MEGA 5. 20软件分析序列各位点碱基替换规律[21]。由表4可以看出,整体位点的转换主要出现在C与T之间,颠换则主要出现在T与A之间,转换/颠换比值(R值)为0.83。对密码子各位点分析发现,转换和颠换主要在密码子第2位点上发生,且转换与颠换值相当(R=0.74)。此外,第1、3位点R值分别为0.59、1.29。无论整体或是密码子各位点,R值均小于2,表明该序列转换与颠换未达到饱和,在构建系统发育树时要考虑转换与颠换的发生概率。
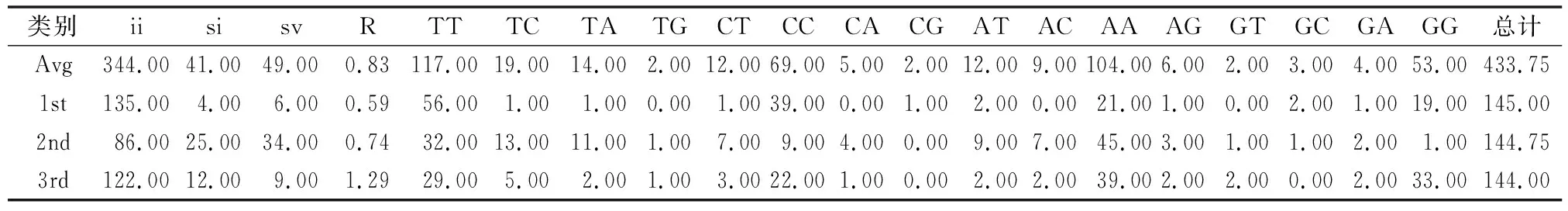
表4 核甘酸碱基替换结果
注:Avg表示平均频率,1st表示第1位点,2nd表示第2位点,3rd表示第3位点,ii表示相同碱基对,si表示转换碱基对,sv表示颠换碱基对,R表示转换碱基对/颠换基对。
2.2.3 遗传距离分析 根据Kimure 2-parameter模型分析7种15头长角天牛和1个外种群之间的遗传距离,将转换和颠换考虑在内,利用Bootstrap值(1 000次)进行检验[21]。由表5可以看出,长角天牛属内相同物种内的遗传距离数值介于0.000~0.016内,平均遗传距离为0.01,其中小灰长角天牛种内距离最小,为0~0.014;斜带长角天牛种内距离最大,为0.016;长角天牛属不同种间的遗传距离介于0.055~0.629之间,平均遗传距离为0.350,其中相似种A.princeps与白带长角天牛之间的遗传距离最小,为0.055;斜带长角天牛1与A.reticulatus之间的遗传距离最大,为0.629。可见,同一物种内的遗传距离较近,基本不受其地理分布的影响;同属不同种类间的遗传距离较远,呈现出较为明显的遗传差异性,可考虑将此段序列作为鉴别不同物种的依据。
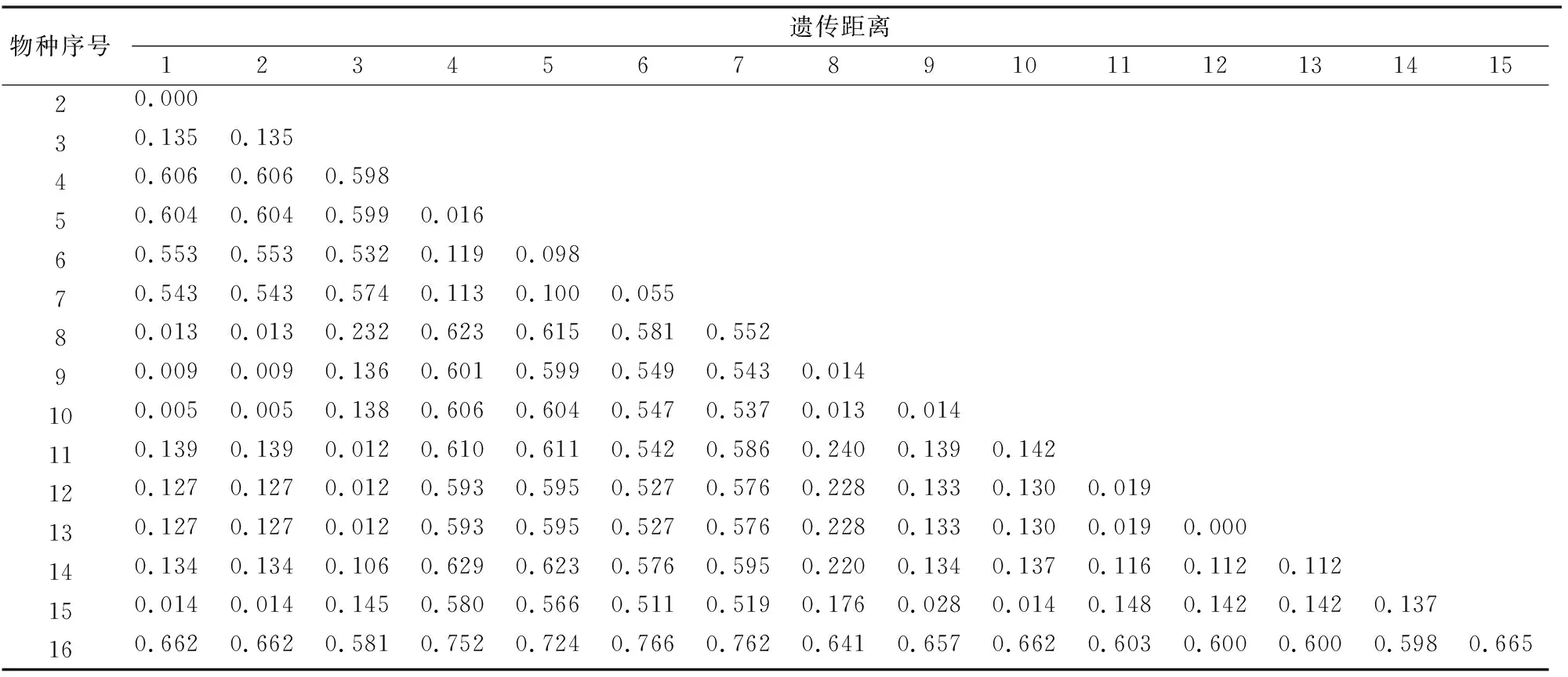
表5 基于Kimure 2-parameter模型长角天牛属的种内、种间遗传距离
注:1表示小灰长角天牛1;2表示小灰长角天牛2;3表示大灰长角天牛1;4表示斜带长角天牛1;5表示斜带长角天牛2;6表示A.princeps;7表示白带长角天牛;8表示小灰长角天牛5;9表示小灰长角天牛3;10表示小灰长角天牛4;11表示大灰长角天牛2;12表示大灰长角天牛3;13表示大灰长角天牛4;14表示A.reticulatus;15表示Acanthocinussp. CA14_3.01;16表示棕榈象甲。
2.2.4 系统发育树的建立 采用MEGA 5.20分析软件,选择棕榈象甲为外群,以长角天牛属7种昆虫共15条COⅠ序列为靶标,使用邻接法建立系统发育树。从图2可以看出,从整体上看,长角天牛属各近似种聚为一大支,而外群棕榈象甲单独成1支,此聚类结果与形态学鉴定结果一致。在长角天牛属所聚的一大支中,7种长角天牛又分聚为2支,其中小灰长角天牛、大灰长角天牛、A.reticulatus和Acanthocinussp. CA14_3.01聚为1支,说明这4种长角天牛的遗传进化关系较为接近,互为近缘种;同时,又与聚为另1支的斜带长角天牛、A.princeps及白带长角天牛遗传进化关系较远,也说明彼此之间的亲缘关系较远,2支互为姊妹群。从局部来看,同一种类不同个体之间各聚为一小支,且置信度很高,这样就很容易和其他种类的长角天牛区分开来。上述结果表明,长角天牛属不同种类之间的系统进化差异明显,因此可将此DNA条形码用作长角天牛属不同种类分子鉴定的依据。

3 结论与讨论
一般而言,使用DNA条形码对物种进行快速准确的鉴定须要满足2个必要条件:首先每个物种都要具有独特且又相对保守的DNA条形码序列;其次是该条形码序列种间差异必须远大于其种内的差异[8]。本研究所采用的线粒体COⅠ基因序列就满足上述条件。首先,地球上几乎所有真核生物都含有线粒体COⅠ基因,且该基因含有很多相对保守的遗传信息位点。其次,对7种15头长角天牛基于线粒体COⅠ基因的种内及种间遗传距离进行计算,发现种内遗传距离为0.000~0.016,平均为0.01;种间遗传距离介于0.055~0.629之间,平均为0.35;种间遗传距离是种内遗传距离的35倍,完全符合种间差异应大于种内差异10倍以上的物种鉴定原则。事实上,线粒体COⅠ基因常被作为DNA条形码,用于昆虫种类鉴定及系统发育学的研究,类似的基因还有Cytb、18S、28S、5.8S、ITS、rbcL、matK等[22]。王颖等利用COⅠ基因作为DNA条形码,实现了对山东口岸进境原木截获蚊虫种类的快速鉴定[23]。董昆应用COⅠ基因分析了苹果蠹蛾不同地理种群间的遗传差异,并实现了对其幼虫的快速鉴定[24]。赵文静等利用COⅠ及ITS序列作DNA条形码深入研究了环带库蚊的分类地位和杂鳞库蚊复组内各亲缘种的系统发育关系[25]。
目前我国口岸昆虫种类鉴定主要依赖于完整成虫的形态特征,而实际上口岸截获到的多是昆虫的卵、幼虫、蛹及肢体残破的成虫。卵、幼虫和蛹须要花费较长时间培养至成虫阶段才能进行种类鉴定,而肢体残破的成虫则失去了形态学鉴定的价值。用DNA条形码技术就能很容易突破上述形态学鉴定方法遇到的瓶颈[26]。但是目前该技术[27]在长角天牛属种类鉴定中的应用还鲜有报道。本研究通过对5种长角天牛COⅠ基因所测序列与GenBank部分长角天牛COⅠ序列进行比对分析,发现该段序列既能高效区分部分长角天牛种类,又能提供丰富的物种亲缘关系信息。此外,本研究还首次对斜带长角天牛、白带长角天牛和A.princeps的COⅠ基因序列进行测序,不仅补充了长角天牛属COⅠ基因数据库,还将为基于COⅠ基因快速鉴定其他种类天牛的研究提供有益参考。值得注意的是,由于本研究未收集到全部的长角天牛属种类,仅能针对部分种类进行分析,因此本研究采用的DNA条形码尚不能确定可以完全鉴别所有的长角天牛种类,只能说明DNA条形码技术[27]在长角天牛属昆虫分子鉴定上具有可行性。后续研究应围绕收集长角天牛属其他种类,完善COⅠ基因数据库,结合多个基因共同分析等方向努力,以期获得更高效的DNA条形码,进而得到更精确的系统发育进化树。
