钾素水平对CO2加富条件下黄瓜光合适应发生的影响
2018-12-25达布希拉图李珊珊何丽琼海梅荣
达布希拉图,李珊珊,何丽琼,海梅荣
(1.云南农业大学资源与环境学院,云南 昆明 650201;2.云南农业大学农学与生物技术学院,云南 昆明 650201)
CO2作为植物光合作用原料,多数C3植物的CO2饱和点为1 000 μmol·mol-1左右。当其它环境条件适宜时,现有大气CO2浓度可能是C3植物光合作用的主要限制因子。在封闭的设施环境中,光照充足时,CO2浓度会降至100 μmol·mol-1以下,作物处于严重的CO2缺乏状态[1],因而,CO2施肥在设施农业生产中增产效果明显。但植物长期处于高浓度CO2下时,一些叶片的光合速率会逐渐下降,最终会低于自然大气CO2浓度下生长的对照植物水平[2],这种现象被称为光合适应。光合适应的发生机制可以归纳为:光合产物超长累积引起的反馈抑制[3];核酮糖-1,5-二磷酸(RUBP)羧化限制和再生限制[4];气孔状态限制[5];以及呼吸作用限制[6]。植物叶片荧光反应对CO2加富的响应为:CO2浓度倍增使鼓粒期绿豆叶片初始叶绿素荧光值(Fo)增加,降低了最大叶绿素荧光值(Fm)、PSII原初光能转化效率(Fv/Fm)和潜在光化学效率(Fv/Fo),但对蕾期影响不显著[7];也有研究称CO2浓度倍增使PSII原初光能转化效率(Fv/Fm)和潜在活性(Fv/Fo)提高,提高了叶绿体捕获光能的效率,提高植物的光合速率[8];而Pospisilova等[9]对高CO2浓度下生长4周的烟草的研究显示,叶绿素含量及PSII活性均无明显变化,似乎与光合适应无关。
以上研究都是在自由CO2加富条件下(FACE)开展的。关于设施CO2施肥条件下(阶段性CO2加富,且浓度更高)植物光合适应是否发生,其光合特性变化规律是否会与FACE条件下相同,却少有报导。K素能促进光合产物的运输、调节气孔开放,增强植物体内、外CO2的交换,因此,K素水平的提高可能会对CO2加富后期的光合特性的变化有所影响,进而缓解光合适应。短期内CO2浓度和K肥对棉花的叶面积、光合作用、干物质累积及分布均存在交互作用[10],但缺乏长期处理对植物光合作用影响的研究。并且在CO2加富条件下K素对光合荧光反应及CO2响应曲线、光响应曲线由来的光合特性没有深入的报导。黄瓜是主要的设施栽培作物,关于设施黄瓜CO2加富的研究报道也较多,且遗传稳定性好。因此,本文研究设施条件下长期CO2阶段性加富与K素互作对黄瓜光合特征的影响,不仅能揭示黄瓜光合作用的变化规律,也为不同CO2加富条件下植物生产提供理论基础。
1 材料与方法
1.1 供试材料与栽培条件
供试黄瓜品种“中农20号”。盆栽试验于2015年3月至6月,在云南农业大学农场大棚内,搭建长5 m、宽1.5 m、高1.5 m的开顶式塑料薄膜培养室完成。供试土壤为高原红壤,土壤化学性质为碱解氮(N)27.53 mg·kg-1,有效磷(P2O5)4.57 mg·kg-1,速效钾(K2O)80 mg·kg-1,有机质16.24 g·kg-1,pH值4.94。施肥以N 200 mg·kg-1、P2O5120 mg·kg-1、K素水平(K2O 200、400 mg·kg-1)为标准,用尿素(N 46%)、重过磷酸钙(P2O546%)、硫酸钾(K2O 51%)随水冲施,氮、磷肥分3次(15 d一次)施完,钾肥随试验设计施用。
黄瓜种子置于装有湿润滤纸的培养皿中,在25℃培养箱中恒温催芽,48 h催芽至2~3 cm,播种于50穴的育苗盘(单孔大小4.8 cm)中育苗,育苗基质配方为泥炭土∶珍珠岩∶蛭石=3∶1∶1,每天浇1次水。长出1片真叶后选取长势一致的黄瓜苗,移栽至盆内(盆高30 cm,直径20 cm);移栽15 d长至3片真叶后,每个K素水平下随机选取6盆置于气候室,采用纯度99.9%的液态CO2钢瓶,经减压缓冲后向室内供应CO2,用Telaire7001红外二氧化碳检测仪定时监测棚内CO2浓度,调节气候室内CO2浓度为(1 000±200)μmol·mol-1。每天9:00~12:00施用CO2气体,即阶段性CO2施肥,共处理35 d。温室内环境条件:午间最大阳光辐照强度为(1 000±600)μmol·m-2·s-1,温度为白天(31±6)℃、夜晚(22±3)℃。
1.2 试验设计
试验设2个CO2浓度:自然大气CO2浓度(380 μmol·mol-1)与高CO2浓度(1 000 μmol·mol-1);每个CO2浓度下设2个K素水平处理(K2O 200、400 mg·kg-1),共4个处理,每个处理6个重复,每盆装土7 kg。K素200 mg·kg-1处理施肥时间与方法同氮、磷肥,400 mg·kg-1处理中其余的200 mg·kg-1与CO2处理开始时间相同。
试验处理:CF表示CO2浓度380 μmol·mol-1,K素水平200 mg·kg-1,为对照;T表示CO2浓度1 000 μmol·mol-1,K素水平200 mg·kg-1;TK表示CO2浓度1 000 μmol·mol-1,K素水平400 mg·kg-1;K表示CO2浓度380 μmol·mol-1,K素水平400 mg·kg-1。
1.3 测定项目及方法
1.3.1 光合参数测定
采用便携式LI-6400光合作用分析仪(Li-Cor,Lincoln,NE),张强等[11]的方法测定,于9:00~11:30测定自然条件下和高浓度CO2条件下黄瓜2、14、24、30、35 d时的叶片净光合速率(Pn)。测定条件:光强为800 μmol·m-2·s-1;温度为25℃;CO2浓度为各处理浓度(380、1 000 μmol·mol-1),测定部位按照每次都为第3片新功能叶的原则开展。
采用6400-40荧光叶室,测定初始叶绿素荧光参数。将待测叶片用夹子夹住暗适应1 h,然后测定并记录叶片Fm、Fo和Fv′/Fm′,测定部位按照每次都为第3片新功能叶的原则开展。
测定光响应曲线时,叶室温度控制在25℃,通过CO2注入系统控制参比室的相对浓度为380 μmol·mol-1,预热20 min后夹住叶片,并在光照强度600 μmol·m-2·s-1下诱导3~5 min;待稳定后启动光响应曲线自动测量程序。利用内置的红蓝光源设定光照强度梯度(即光合有效辐射,PAR)为:1 500、1 200、800、600、400、300、200、150、100、80、60、40、20、0 μmol·m-2·s-1,每个光强平衡5 min,光源由仪器自带的LED提供。测定部位按照每次都为第3片新功能叶的原则开展。计算最大光合速率(Amax)、光饱和点(LSP)、光补偿点(LCP)、表观量子产额(AQY)等光合响应指标。
测定CO2响应曲线时,叶室温度设置为25℃;光强设置为800 μmol·m-2·s-1;CO2浓度梯度通过CO2注入系统控制参比室的相对浓度为:1 500、1 200、1 000、800、600、400、360、300、250、200、150、100、50 μmol·mol-1。测定部位按照每次都为第3片新功能叶的原则开展。采用CO2响应曲线自动测量程序进行测定;在每个相对CO2浓度下均平衡5 min,并在稳定后同时记录叶片净光合速率,计算CO2羧化效率(CE)。
1.3.2 植株生物量的测定
CO2处理第35 d,摘取黄瓜果实,测量鲜重。
1.3.3 数据处理
采用SigmaPlot 10.0计算光响应曲线和CO2响应曲线相关参数值。根据方程y=a-c·e-bx拟合光响应曲线和CO2响应曲线并计算相关参数[12],根据光响应曲线的拟合结果计算出最大光合速率(Amax为参数a)、光补偿点[LCP,为ln(a/c)·(-1/b)]、光饱和点[LSP,为ln(0.1 a/c)·(-1/b)]、暗呼吸速率(Rd,为a-c)以及表观量子产额[AQY,为b·c·eb·(-cp)],并根据CO2响应曲线的拟合结果计算出RUBP羧化效率[CE,为b·c·eb·(-cp)]。
所有数据3次重复,采用Excel 2007和SPSS 17.0软件进行数据的处理和统计分析。采用独立样本t检验进行显著性分析,P≤0.05时,处理间差异显著。
2 结果与分析
2.1 K对CO2加富条件下黄瓜鲜果重的影响
由图1可知,CO2加富第35 d,CO2加富处理(T)显著提高了黄瓜鲜果重(较CF),在CO2加富的同时提高K素水平(TK),能够更显著地增加黄瓜鲜果重,且高于T处理。在自然CO2浓度(380 μmol·mol-1)的基础上增加K素水平(K),也能显著提高黄瓜鲜果重,且增重显著高于单CO2加富处理。CO2加富与K素水平对黄瓜鲜果重具有显著交互作用(表1)。

图1 CO2加富与K素水平对黄瓜鲜果重的影响注:图中不同小写字母表示处理间差异显著(P<0.05)。下同。

表1 CO2加富与K素水平对黄瓜鲜果重交互作用
2.2 K对CO2加富条件下黄瓜叶片光合速率的影响
在CO2加富的第2、14、24、30、35 d进行了黄瓜叶片光合作用的测定(图2)。结果显示:在CO2加富前期(14、24 d)黄瓜光合作用随CO2加富(T)而增加,在第24 d CO2加富并增加K素水平(TK)黄瓜光合作用较单独CO2加富(T)更高。CO2加富30 d发现,4个处理均处于同一光合速率水平,处理间无显著差异;第35 d的光合速率数据显示,CO2加富处理(T)显著低于环境CO2浓度(CF)下,CO2加富并增加K素水平(TK)高于单独CO2加富(T),其中环境CO2浓度下增加K素水平(K)光合速率最高。
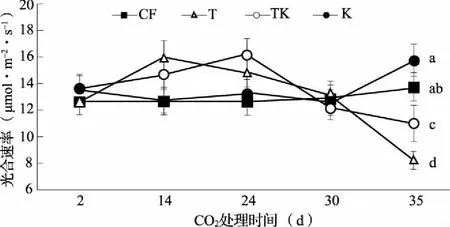
图2 CO2加富与K素水平对黄瓜光合速率的影响
2.3 K对CO2加富条件下黄瓜叶片叶绿素荧光参数的影响
针对CO2加富后光合作用升高(14 d)和降低(35 d)时的黄瓜叶片叶绿素荧光参数进行了分析(图3、4,表2)。在CO2加富后光合作用升高的第14 d,两个CO2加富处理(T、TK)与单独增加K素水平处理(K)的Fo均显著高于环境CO2浓度处理(CF)。其中,单独CO2加富处理(T)Fo最高。在CO2加富后光合作用降低的第35 d,环境CO2浓度处理(CF)较第14 d Fo有增高的趋势,但在35 d两个CO2加富处理(T、TK)较环境CO2浓度处理(CF)Fo有降低的趋势(图3)。

图3 CO2加富与K素水平对黄瓜初始叶绿素荧光值的影响
在CO2加富后光合作用升高的第14 d,两个CO2加富处理(T、TK)与单独增加K素水平处理(K)的Fm均显著高于环境CO2浓度处理(CF)。其中,单独CO2加富处理(T)与单独增加K素水平处理(K)的Fm最高。在CO2加富后光合作用降低的第35 d,环境CO2浓度处理较第14 d Fm有增高的趋势,但在35 d的处理间无显著差异(图4)。

图4 CO2加富与K素水平对黄瓜最大叶绿素荧光值的影响
CO2加富第14 d,在相同CO2浓度的两个K素水平间实际光化学效率(Fv′/Fm′)无显著差异;在相同K素水平时CO2加富可显著提高Fv′/Fm′值。CO2加富第35 d,在相同CO2浓度的两个K素水平间Fv′/Fm′也无显著差异;但与第14 d相反,在相同K素水平时CO2加富显著降低Fv′/Fm′值(表2)。另外,Fv/Fm值均在0.81~0.82之间,且处理间及处理天数间无差异。

表2 CO2加富与K素水平对PSⅡ实际光化学效率的影响
注:不同小写字母表示在同一天相同CO2浓度下不同K素水平间的差异显著,*表示同一天相同K素水平下不同CO2浓度间的差异显著,n=3,P<0.05。下同。
2.4 K对CO2加富条件下黄瓜光响应、CO2响应曲线相关光合参数的影响
针对CO2加富后光合作用升高(14 d)和降低(35 d)时的黄瓜叶片光响应曲线拟合方程计算获得的相关光合参数进行了分析(表3)。CO2加富第14 d在自然CO2浓度下,增加钾素水平可增高Amax,在此基础上分别增加CO2浓度也能显著增高Amax、LSP、LCP;在第35 d发现与第14 d相反的结果,即在自然CO2浓度下,增加钾素水平可降低Amax,在此基础上分别增加CO2浓度也能显著降低Amax、LSP,在第35 d LCP表现为在低K素水平随CO2浓度升高而显著增高,但此时增加K水平没有增高LCP。AQY在本试验各处理中无显著差异。CO2加富第14 d在自然CO2浓度下,增加钾素水平,CO2加富,CO2加富的同时增加钾素水平均显著增加了CE,在第35 d各处理间无显著差异。
3 讨论与结论
CO2加富在一段时间内能加快许多植物的生长,随后出现生长缓慢甚至停滞现象,导致生产力的降低[13]。CO2加富的方法有开顶式气室、自由CO2加富系统、温室等,不同方法得到的试验结果往往有较大的差异,FACE条件下的研究主要用来模拟未来的大气CO2浓度升高对植物的影响,其加富时间是连续的,得到的结果能够预测未来环境条件变化对农业生产的影响,而OTC与FACE系统相比,能实现较高浓度的CO2加富,可能更有利于研究植物对CO2的响应及其机理,被广泛应用于环境CO2加富对植物影响的研究中。关于在OTC内进行CO2加富,使作物增产的研究报告很多,其中何丽琼等[14]研究发现CO2加富显著增加了黄瓜植株干重,高CO2浓度条件下增加K素水平可继续增加黄瓜生物量,在自然CO2浓度下增加K素水平黄瓜生物量则无显著变化。本试验对这一研究发现,CO2加富与K素水平增高均可以显著提高黄瓜鲜果重,且CO2加富与K素水平增高有交互作用,即在试验各处理中CO2加富条件下增高K素水平,黄瓜鲜果重最高。

表3 CO2加富与K素水平对光响应、CO2响应曲线相关光合参数的影响
FACE条件下,短期CO2加富(几分钟到几天)内可使植物光合速率显著提高,长期CO2加富条件下(数周到数月),光合速率则逐渐降低,最终与自然大气CO2浓度下的无差异或低于自然条件下生长的,出现光合适应现象[15-16]。本试验根据前期研究经验,在CO2加富第2、14、24、30、35 d分别测定了黄瓜第3功能叶片光合速率。加富14~24 d时光合速率升高,加富14 d时升高幅度为23.8%,加富24 d时升高14.1%,35 d时则与自然CO2浓度相比显著降低,出现光合适应,发生的时间在24~30 d之间。这与孙玉文等[17]、单国雷等[18]在甜瓜、西瓜的研究中得到的结论相符,即CO2加富时间以25 d为最优。因此可以推断,瓜果类蔬菜在OTC内高浓度的、阶段性的CO2加富也会产生光合适应。如果要持续地在采摘期提高产量,可能需要优化其他生长因子,如其他矿质养分供应。本研究中CO2加富第35 d,即黄瓜光合适应发生时期,在两个CO2浓度下增加K素水平,均提高了黄瓜光合速率,尤其在CO2加富处理也表现出显著差异,Reddy等[10]对玉米的研究也发现,CO2加富与K素水平在光合速率上存在交互作用,因此可以推断增加K素水平有缓解光合适应的潜力。
荧光参数反映了植物叶片PSII功能、活性的变化,本试验中Fv/Fm值在整个生长期变化幅度<1%。张其德等[19]与刘家尧等[20]的研究表明,在FACE条件下CO2加富使得Fv/Fm显著增加;郝兴宇等[7]的研究则表明长期CO2加富使绿豆Fv/Fm降低6%,即相同的CO2加富在不同作物及处理条件下结果不尽相同。本试验Fv/Fm较稳定,只有彭长连等[21]的研究发现,CO2浓度倍增对整个生育期水稻Fv/Fm无显著影响,与本试验结果相近。因此本研究认为无论CO2加富时间长短,黄瓜叶片PSII没有感知CO2加富为环境胁迫,并且K素水平也没有被感知为胁迫。郝兴宇等[7]对绿豆的研究显示,CO2浓度倍增使鼓粒期绿豆叶片Fo増加;张其德等[22]认为,CO2浓度倍增提高了光合色素含量,提高了PSII活性、光能转化效率,促进了PSII反应中心的开放;而Pospisilova等[9]对高CO2浓度下生长4周的烟草的研究显示,叶绿素含量及PSII活性均无明显变化;Hogan等[23]也指出CO2加富能促使植物捕获更多光能,减少高光强对PSII反应中心的破坏。本试验发现,在短期(14 d)CO2加富时随着Pn的增加,Fo、Fm与Fv′/Fm′值均显著增加,在长期(35 d)CO2加富时Fo、Fm无显著变化,Fv′/Fm′值降低,且K素水平的增加没有改变CO2加富带来的变化,说明黄瓜光合适应的起因不是发生在PSII这个过程中。
在FACE条件下,长期CO2加富使植物净光合速率的降低伴随着RUBP羧化酶含量和活性的降低,Rubisco基因的转录及表达受到抑制,RUBP羧化限制可能是引起光合适应现象的原因[4]。Koch等[24]的研究说明短时间CO2浓度上升可以使RUBP羧化酶活性提高30%左右。本研究中,CO2加富14 d时,高CO2浓度条件下黄瓜CE升高,RUBP羧化酶含量和活性可能提高,CO2加富35 d时,CE无显著变化,RUBP羧化酶可能与光合下调无关。本试验中,其他光响应参数和CO2响应参数的变化亦显示出相同的趋势,CO2加富14 d时,Amax、LSP、LCP高于自然条件下,而加富35 d时,高CO2浓度条件下黄瓜Amax、LSP均低于自然条件下,这些指标的变化与后期光合适应现象的出现相符;AQY无显著变化。郑炳松等[25]的研究表明钾离子可提高光合关键酶RUBP羧化酶含量和活性,低钾会导致RUBP羧化酶含量降低。本试验中,CO2加富14 d时,自然条件下增加K素水平,Amax、LSP、LCP、CE显著增加,CO2加富35 d时,却均降低;并且在高CO2浓度下增加K素水平也为降低,即在光合适应发生的第35 d没有发现K素水平能够缓解这些光合参数降低的迹象,因此推断RUBP羧化限制不是引起光合适应的直接原因,而是光合适应发生后的结果。
在本研究条件下,CO2加富可使黄瓜鲜果重显著增加,并与钾素有显著交互作用。黄瓜在OTC条件下也会产生光合适应,发生在CO2加富第24~30 d间,增加K素水平有缓解光和适应的潜力。关于光合适应发生的原因,本文通过对荧光参数及光、CO2响应曲线相关参数的研究发现,黄瓜光合适应的发生可能不是直接作用在PSII及CO2羧化阶段,可能将研究重点放在光合产物反馈抑制方面更有意义。
