七鳃鳗属3种类的亲缘关系分析
2018-12-20常玉梅黄晶孙博苏宝锋梁利群
常玉梅,黄晶,孙博,苏宝锋,梁利群
(中国水产科学研究院黑龙江水产研究所,中国水产科学研究院特殊生境鱼类种质特性与抗逆育种重点实验室,农业部淡水水产生物技术与遗传育种重点实验室,淡水鱼类育种国家地方联合工程实验室,黑龙江 哈尔滨 150070)
七鳃鳗(Lamprey)是现存的古老无颌类脊椎动物的代表种,是脊椎动物演化发育生物学研究的重要模式动物。全球有3科(七鳃鳗科Petromyzonidae、地蝗科Geotriidae和水螨科Mordaciidae),10属,超过40种。其中4种隶属于南半球的地蝗科和水螨科,而其余种都属于北半球的七鳃鳗科[1,2]。
七鳃鳗属Lethenteron隶属于七鳃鳗科,主要分布在欧亚大陆[3]。基于形态特征和分子数据,目前大多数学者认为该属至少有6个有效种:包括一个洄游寄生种北极七鳃鳗(Lethenteron camtschaticum,以前为Lampetra japonica)、两个淡水非寄生种溪七鳃鳗(Le.kessleri,以前为 Lampetra japonica kessleri和 Le.reissneri,以前为 Lampetra reissneri)[4,5]、两个日本学者发现的隐秘种Lethenteron sp.N和Lethenteron sp.S[6],以及一个朝鲜七鳃鳗(Le.morii,以前为 Lampetra morii&Eudontomyzon morii)[2,7]。但近期有学者发现,两个淡水溪七鳃鳗在表型、核DNA及mtDNA高度相似,将二者归为同一种Le.reissneri complex[6,8]。最新研究表明,这两个淡水溪七鳃鳗与北极七鳃鳗又无明显的种间差异,将其视为北极七鳃鳗的两种生态类型[8,9]。因此看出,七鳃鳗属虽被认定为一个有效属,但是其种类的归属仍存在分歧。
线粒体细胞色素c氧化酶亚基I(cytochrome coxidase subunit I,COI)在种间的遗传变异显著大于种内的遗传变异,即使两个亲缘关系非常相近的物种,COI基因的差异也会超过2%[10,11],是物种鉴定的有效基因。如笔者利用COI基因分析了绥芬河珠星三块鱼(southern forms ofTribolodon hakonensis)和日本珠星三块鱼(northern forms of Tribolodon hakonensis)的亲缘关系,发现二者之间遗传距离大于2%,表明这两个群体已出现明显分化,不能视为同一种[12]。
本研究通过分析七鳃鳗属种属分类存有争议的3个种,北极七鳃鳗、溪七鳃鳗及朝鲜七鳃鳗的COI基因,以明确这3个种的分类地位,为更好地保护和利用这些优良的种质资源提供科学依据。
1 材料与方法
1.1 材料
本研究共用到102尾样本,各样本的采集国别、地点、GenBank登录号及出处见表1。样品来源如下:(1)本研究在黑龙江抚远采集北极七鳃鳗46尾鳍条于95%乙醇中保存;(2)从NCBI GenBank数据库中下载来自俄罗斯26个采样点(包括黑龙江、萨哈林岛、国后岛、堪察加半岛、北冰洋、西伯利亚南部、乌特霍洛克河和克拉维耶姆河)及美国阿拉斯加育空河的北极七鳃鳗COI序列36条;(3)从NCBI GenBank数据库中下载来自俄罗斯乌特霍洛克河、克拉维耶姆河和米斯蒙特河的溪七鳃鳗Le.reissneri complex COI序列 17 条;(4)从 NCBI Gen-Bank数据库中下载来自中国辽宁丹东和黑龙江抚远的朝鲜七鳃鳗线粒体全基因组序列3条。
1.2 基因组DNA的提取
剪取少量95%乙醇保存的鳍条,滤纸吸干乙醇,放入2mL离心管中加入600μLDNA裂解液(成分:1mol/LTrispH8.0、5MNaCl、0.5mol/LEDTApH8.0、10%SDS和200μg/mL蛋白酶K)于55℃过夜消化,次日加入等体积的酚/氯仿混合液(1∶1)抽提1次,12 000r/min室温离心10min,吸取上清(~550μL)加入1mL无水乙醇沉淀,12 000r/min室温离心10min,70%乙醇洗涤1次,室温干燥10min,加入0.1×TE溶解。采用1%琼脂糖凝胶电泳检测DNA的完整性,Nanodrop 8000(Thermo Fisher Scientific,MA,USA)测定 DNA 浓度,稀释至 10ng/μL,4℃保存备用。
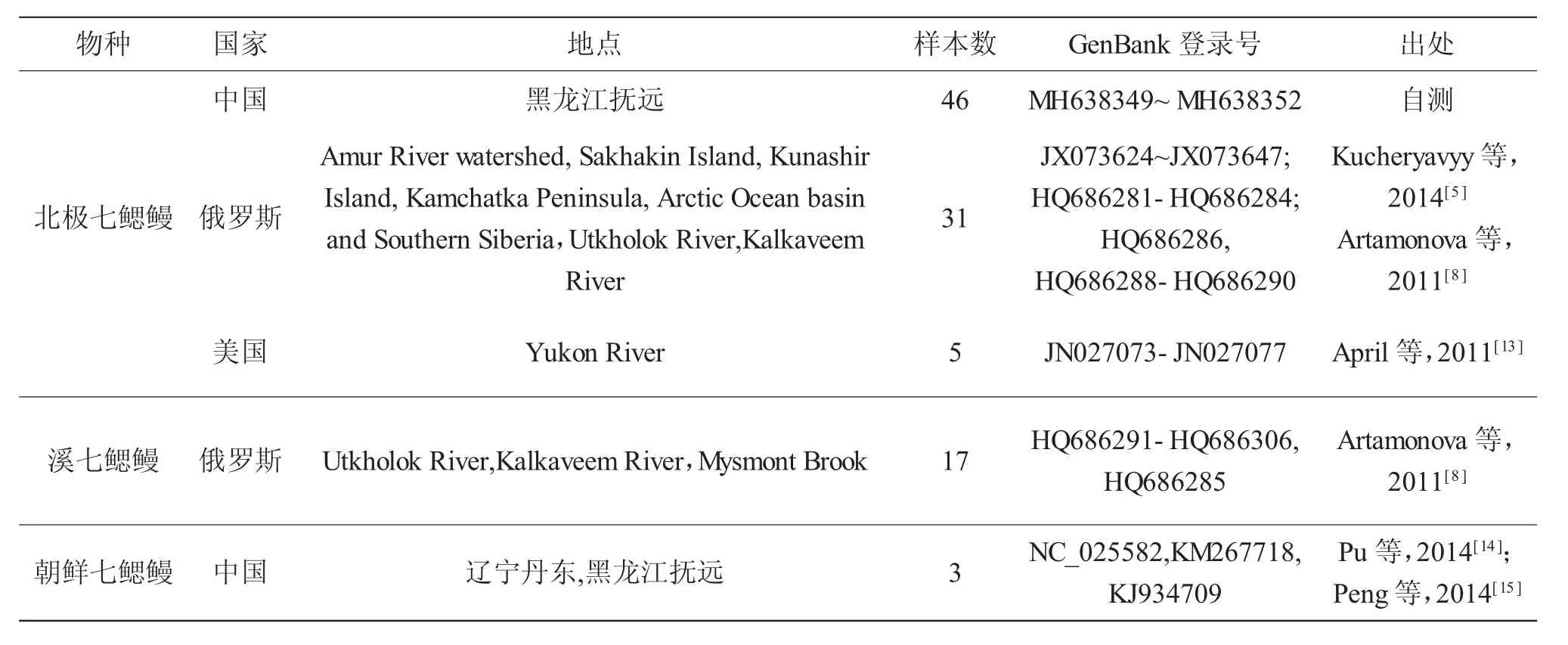
表1 3种七鳃鳗样本来源信息Tab.1 Samples information of three lamprey species
1.3 COI基因的引物来源及PCR扩增
线粒体COI引物参照Prasanna等[16]报道的通用引物(COI-F:5′-TCAACCAACCACAAAGACATTG GCAC-3′;COI-R:5′-TAGACTTCTGGGTGGCCAAA GAATCA-3′)。PCR 反应总体积体系为 25μL,其中包含自制 PCR buffer mix18μL(包括 50mmol/L KCl、10mmol/L Tris-HCl、0.10%Triton X-100、1.5mmol/L MgCl2、0.10%NP-40、0.01%明胶、200μmol/L 4 种 dNTP),浓度10μmol/L的上、下游引物各1μL,Taq DNA聚合酶 0.2μL(Fermentas,5U/L),模板 DNA 2μL,去离子灭菌水补至总体积25μL。反应程序为95℃预变性 3min,35 个循环包括 95℃15s、57.7℃30s、72℃30s,最后72℃延伸7min。PCR扩增产物用1.5%的琼脂糖凝胶电泳检测合格后,送至北京诺赛基因公司进行测序。
1.4 序列处理及分析
将测序得到的序列文件用DNASTAR.Lasergene 7.1中的EditSeq模块转换成fasta格式用于后续分析。采用ClustalX1.83对COI基因序列进行多重比对,获得对齐后的同源序列,采用NCBI网站提供的Bankit提交到GenBank数据库中。利用DNAsp 5.10.01鉴定单倍型;使用MEGA7.0计算所得单倍型的碱基组成、单倍型间的替换率和转换/颠换R值。将从NCBI GenBank数据库中下载的56条已知COI序列和mtDNA全基因组序列(北极七鳃鳗36条、溪七鳃鳗17条和朝鲜七鳃鳗3条)与本研究得到的46条序列合并后采用邻接法(Neighbor-Joining)构建基于Kimura双参数模型(Kimura 2-parameter,K2P)的单倍型进化树,并经1 000次自展检验(Bootstrap),选用黄鳝 Monopterus albus COI基因相应片段(GenBank登录号:MF122533)作为系统进化分析的外类群;用MEGA7.0计算基于K2P模型下各个类群内部的遗传距离及各类群间的平均遗传距离。
2 结果与分析
2.1 单倍型分析
序列经比对处理后得到COI有效序列长度为451bp。102条有效序列共检测到16种单倍型(图1)。北极七鳃鳗共检测到14种单倍型,其中中国群体有4种单倍型(GenBank登录号:MH638349~MH638352),俄罗斯群体多样性较高,有11种单倍型,美国群体有3种单倍型,大部分个体共享2种单倍型Hap1和Hap5;溪七鳃鳗有3种单倍型,均与北极七鳃鳗共享;朝鲜七鳃鳗独享2种单倍型,分布在两个不同的地理群体(辽宁丹东和黑龙江抚远)(表2)。单倍型碱基组成分析显示,A、T、C、G 4种碱基所占比例恒定,平均含量分别为27.7%、30.3%、28%和14%,A+T含量(58%)明显高于C+G含量(42%)。平均碱基替换率为19.03,碱基交换/颠换R值为3.19。
2.2 单倍型序列分歧度分析

图1 COI基因在3种七鳃鳗群体中的16种单倍型Fig.1 Sixteen haplotypes of COI gene found in three lamprey species
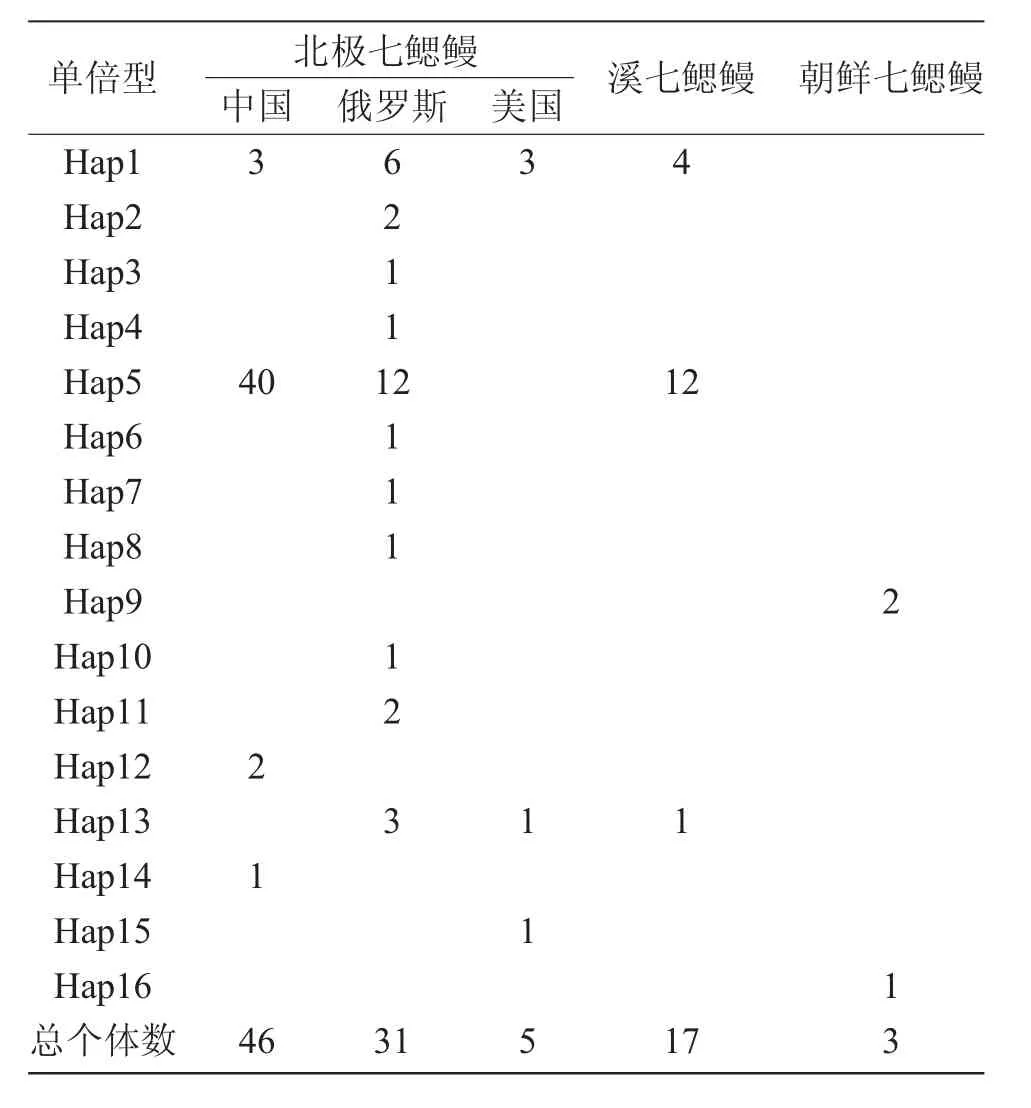
表2 COI基因的16种单倍型在3种七鳃鳗群体中的分布Tab.2 Distribution of 16 haplotypes of COI gene in threelamprey species
16种单倍型间的序列分歧度计算结果见表3。由表3可知,北极七鳃鳗14种单倍型间的序列分歧度为0.002~0.011,其中中国群体4种单倍型间的序列分歧度只有0.002~0.004,俄罗斯群体11种单倍型间的分歧度为0.002~0.011,美国群体3种单倍型分歧度为0.002~0.007;溪七鳃鳗和北极七鳃鳗共享3种单倍型,序列分歧也只有0.002~0.004;朝鲜七鳃鳗独享2种单倍型,Hap9和Hap16,序列分歧度达到0.020。辽宁丹东群体单倍型Hap9与北极七鳃鳗和溪七鳃鳗的单倍型序列分歧只有0.002~0.011,而黑龙江抚远群体单倍型Hap16与北极和溪七鳃鳗的单倍型序列分歧较大,为0.016~0.025。
2.3 3种七鳃鳗的系统进化关系
采用MEGA7.0软件中的邻接法构建3种七鳃鳗16种单倍型间的系统发育进化树(图2)。由图2可知,除了朝鲜七鳃鳗黑龙江群体Hap16单独聚类外,其余15种单倍型无明显分化,聚为一支。各类群间和类群内平均遗传距离显示,北极七鳃鳗和溪七鳃鳗亲缘关系较近,而朝鲜七鳃鳗与二者较远;北极和溪七鳃鳗种内不同群体无明显分化,而朝鲜七鳃鳗不同群体存在一定分化(表4)。
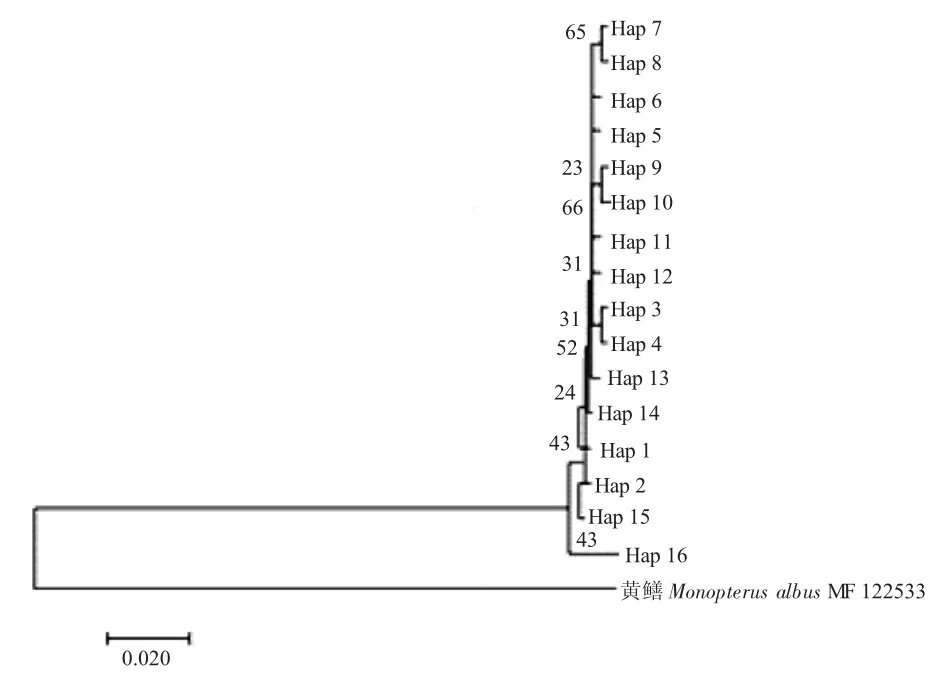
图2 七鳃鳗属鱼类COI基因单倍型的系统发育进化NJ树Fig.2 Neighbour-Joining tree of COI gene in genus Lethenteron

表3 16种单倍型间的序列分歧度Tab.3 Sequence divergenceamong 16 haplotypes

表4 3种七鳃鳗各类群间和各类群内平均遗传距离Tab.4 Mean genetic distance between and within each group in three lamprey species
3 讨论
3.1 北极七鳃鳗和溪七鳃鳗的系统进化关系
北极七鳃鳗主要分布在欧亚大陆的东部、日本列岛,朝鲜半岛的北部及北美大陆的西北部,有溯河寄生型(anadromous-parasitic form)和河流非寄生型(fluvial-nonparasitic form,也叫定居型)两种生活史类型[17-19]。然而,一些学者认为这两种生活史类型的命名和分类也有问题。Yamazaki等[19]利用微卫星标记研究了溯河寄生型、河流定居型和陆封型北极七鳃鳗的群体遗传结构,结果发现,这些同水域的七鳃鳗能够一起产卵,而且存在紧密的基因流,推测这种大小不匹配的交配和不同生活史的可塑性是导致不同类型群体基因流产生的重要原因[20];随后Artamonova等[8]利用COI基因分析俄罗斯北极七鳃鳗洄游型和定居型群体发现,二者具有相同的单倍型,认为不同的生活史类型是由北极七鳃鳗表型的可塑性进化而来。
溪七鳃鳗分布也很广泛,从欧亚东部河流到日本北部,包括阿纳德尔、堪察加半岛、鄂霍次克海、黑龙江、萨哈林岛和日本北部[6,17]。早期学者根据地理分布和生活史,认为溪七鳃鳗Le.kessleri和Le.reissneri是两个独立种[2]。而 Makhrov等[18]分析了来自俄罗斯北部70个河流七鳃鳗的形态、分布、生态、资源量等认为栖息在欧亚大陆北方的七鳃鳗应为北极七鳃鳗的不同生态类型;随后Artamonova等[9]分析了欧亚大陆七鳃鳗属COI部分序列单倍型的分布,同样认为地理分布和生活史均不能成为鉴定这些种类的标准,支持将Le.kessleri和Le.reissneri划分为北极七鳃鳗的两种生态型而非两个独立种。
分子聚类研究发现,北极七鳃鳗和溪七鳃鳗组成一个单独的分支,没有种间差异[6,8]。从本研究的系统聚类也可以看出,来自中国、俄罗斯和美国不同水域的北极七鳃鳗,虽然有14种单倍型,多样性较高,但种内遗传距离只有0.002,14种单倍型聚为一支(图2)。而溪七鳃鳗的3种单倍型均为北极七鳃鳗的主效单倍型(Hap1,Hap5和Hap13),种间遗传距离最小,只有0.001(表4)。由此推测,溪七鳃鳗起源于北极七鳃鳗,支持二者为同一种的不同生态类型,这与日本和俄罗斯学者的观点一致。
3.2 朝鲜七鳃鳗的分类地位
朝鲜七鳃鳗主要分布在朝鲜半岛的中部和北部河流,我国辽宁和黑龙江也有少量分布[14,15,17]。早期命名为Lampetra morii,后来根据形态特征建议为Eudontomyzon morii[7],目前根据分子数据归为Lethenteron属[2,17]。Goto[21]利用 COI基因重建了东亚地区七鳃鳗属的系统进化树,同样支持朝鲜七鳃鳗是七鳃鳗属的一个有效种。本研究结果表明,虽然我国辽宁丹东和黑龙江抚远地区的朝鲜七鳃鳗独享两种单倍型,但是只有黑龙江抚远群体独享的Hap16与北极七鳃鳗的14种单倍型分歧较大,从0.016~0.025不等,平均0.02,达到不同种的分化水平(~2%),认为朝鲜七鳃鳗黑龙江群体为七鳃鳗属的一个有效种。而辽宁丹东群体独享的Hap9与北极七鳃鳗序列分歧较低0.002~0.011,且与黑龙江群体单倍型分歧达到种级分化水平。因此,这种差异是否采样误差导致,需待进一步验证。
综上所述,本研究分析的七鳃鳗属的3个种,实际只有两个有效种,北极七鳃鳗和朝鲜七鳃鳗。其中北极七鳃鳗由于生活环境、大小及表型特征的多样性存在多种生态类型。
