水稻二化螟热激蛋白的研究进展
2018-12-05陆明星李秋雨杜予州
陆明星 李秋雨 杜予州

摘要
水稻二化螟Chilo suppressalis (Walker)是水稻上的主要害虫之一,近年来在部分地区发生为害逐渐加重。热激蛋白是一类胁迫蛋白,在细胞生长、发育、分化、基因转录等功能方面具有重要的作用。本文综述了二化螟热激蛋白的研究进展。目前,已在二化螟体内鉴定出11种热激蛋白,它们分别属于热激蛋白90家族、热激蛋白70家族、热激蛋白60家族和小分子量热激蛋白家族。这些热激蛋白在二化螟生长发育中的调节作用也各不相同;同时,热激蛋白(Cshsp90,Cshsp70和Cshsp60)还与水稻二化螟的滞育有着紧密的联系。不同的热激蛋白对温度的响应大体可以分为三种类型:对高低温都有响应型、对低温响应型和对温度不响应型。最后,分析了二化螟热激蛋白研究过程中存在的问题,提出了该研究领域的展望。
关键词
水稻二化螟; 热激蛋白; 生长发育; 滞育; 温度
中图分类号:
S 435.112.1
文献标识码: A
DOI: 10.16688/j.zwbh.2018314
Research progresses in the heat shock proteins of Chilo suppressalis
LU Mingxing, LI Qiuyu, DU Yuzhou
(College of Horticulture and Plant Protection, Institute of Applied Entomology,
Yangzhou University, Yangzhou 225009, China)
Abstract
The rice stem borer, Chilo suppressalis (Walker), is one of most important rice pests. In recent years, C.suppressalis have caused more and more damages. Heat shock proteins belong to stress proteins, which play very important roles in the development, differentiation, gene transcription of cells. In this review, the progresses in the heat shock proteins of C.suppressalis were summarized. To date, 11 heat shock proteins have been identified from C.suppressalis, belonging to heat shock protein 90, heat shock protein 70, heat shock protein 60, and small heat shock protein family. They played different roles in the development of C.suppressalis. Moreover, heat shock proteins (Cshsp90, Cshsp70 and Cshsp60) are also closely related to the diapause of C.suppressalis. In general, the responses of heat shock proteins to temperature can be divided into three types: response to both heat and cold stress, response to cold stress only and no response to temperature. Last, the problems in the study of heat shock proteins of C.suppressalis were analyzed, and the prospects for the future study was exhibited.
Key words
Chilo suppressalis; heat shock protein; development; diapause; temperature
水稻二化螟Chilo suppressalis (Walker)隸属于鳞翅目Lepidoptera,螟蛾科Pyralidae,俗名钻心虫、蛀心虫或蛀秆虫等,是世界水稻36种螟蛾科蛀茎害虫中分布最广,并且为害最为严重的害虫之一[1]。据报道,自2013年以来,江苏大丰东部沿海地区二化螟种群数量呈上升趋势,为害加重[2];在江西的余干县2016年全年总体偏重发生,重于2015年及近年,发生面积8.05万hm2次[3];在湘南地区的衡阳县、涟源市、衡南县、茶陵县的二化螟偏重发生[4];浙江象山县的水稻二化螟种群在2013—2017年间数量急剧回升[5]。同时,我们从近两年全国主要稻区的调查采样也发现,二化螟在局部地区有逐年加重为害的趋势,需要给予密切关注。水稻二化螟在我国从南到北广泛分布,该虫对温度胁迫有着很强的耐受能力[6-7]。但是,由于在多个地区已经发现水稻二化螟对三唑磷、杀虫单、阿维菌素、氟苯虫酰胺和氯虫苯甲酰胺等药剂都产生了抗性[8-10]。因此,为了阐明在全球变暖条件下水稻二化螟种群数量逐渐增加的原因,同时也为今后开辟新的防控技术提供理论参考,对水稻二化螟热激蛋白的相关研究进行了总结。
热激蛋白 (heat shock proteins, HSPs)先于1962年由Ritossa在研究果蝇唾腺染色体时偶然发现,后在1974年被Tissieres等人研究证实其是一组响应温度胁迫的特殊蛋白[11-12]。它在分子进化上相对保守,广泛分布于各种生物体内,在细胞生长、发育、分化、基因转录等功能方面具有重要的作用。基于分子量和同源性,热激蛋白家族通常分为HSP100,HSP90,HSP70,HSP60和小分子量热激蛋白亚族(small heat shock protein,sHSPs,12~43 kDa)[13-14]。目前,已经有多种热激蛋白从昆虫中鉴定发现,并初步研究了其功能。例如,通过不同高温处理果蝇成虫,发现果蝇耐热性与HSP70的表达成正相关,而且HSPs的表达可以使原本对高温敏感的黑腹果蝇Drosophila melanogaster初生精子细胞具备耐热性[15]。
对蝶蛹金小蜂Pteromalus puparum的研究发现,经过非致死高温锻炼明显提高其在极端温度下的羽化率,原因在于hsp70的迅速上调表达降低了高温带来的损伤[16]。同样,对一种弹尾目昆虫Orchesella cincta的研究发现,该虫耐热性的获得与其hsp70蛋白表达水平呈正相关,但与HSP70 mRNA的表达水平关系不大[17]。悬铃木方翅网蝽Corythucha ciliata的 hsp70响应高温,有助于提高自身的热耐受性[18]。高低温可以诱导中华蜜蜂的hsp24.2和hsp23.0上调表达;同时,RNAi技术沉默了中华蜜蜂的hsp22.6可以明显降低其高低温存活率[19-20]。在所有已研究的与滞育相关的基因中,热激蛋白基因被认为是与滞育关系最为密切的基因[21]。由此可见,昆虫体内的不同热激蛋白具有不同的作用,并在昆虫生命活动中起到重要的作用。本文就水稻二化螟热激蛋白的相关研究进展进行了综述,并提出了该研究领域的展望。
1 二化螟热激蛋白的鉴定与分类
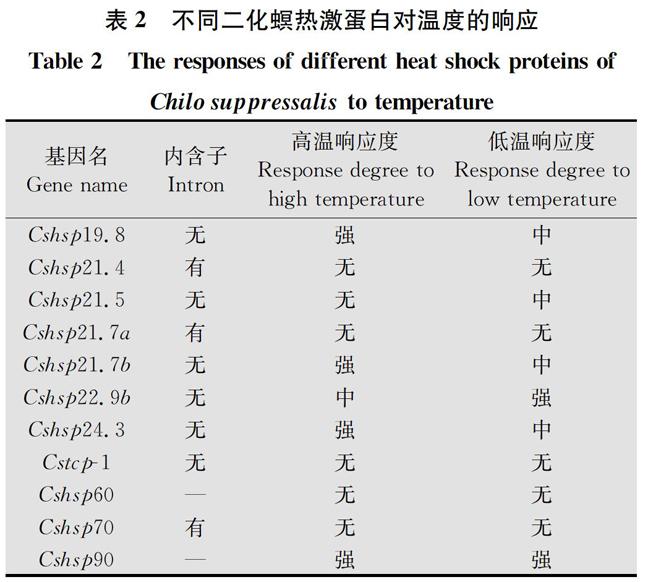
目前已从水稻二化螟体内鉴定分离出11种热激蛋白,分别属于热激蛋白90家族、热激蛋白70家族、热激蛋白60家族和小分子量热激蛋白家族,它们都具有典型的家族特征[22-29]。例如,7种小分子量热激蛋白基因在序列长度上不一样,而且序列变异也较大,但是它们都拥有α-晶状体结构域;二化螟热激蛋白90拥有NKEIFLRVISNASDALDKIRY,LGTIAKSGTK,IGQFGVGFYSCYLVADR,IKLYVRRVPI和GVVDSEDLPLNISREM典型的家族特征。在这11种热激蛋白中,有一种特殊的热激蛋白——T-复合多肽-1(Cstcp-1),该类蛋白隶属于热激蛋白60家族成员。但是,它们的家族特征完全不同,热激蛋白60拥有C端的(GGM)4重复序列,而T-复合多肽-1具有RSAYGPNGMNKMI和QDAEVGDGT家族特征序列(表1)。对这些二化螟热激蛋白进行系统进化树分析表明,这些热激蛋白也明显地分为4个热激蛋白亚族;同时,不同种昆虫的同一家族的热激蛋白成员的亲缘关系也近于同一种昆虫不同热激蛋白家族成员。此外,研究结果进一步表明,二化螟7種小分子量热激蛋白成员实际上分为了3支,说明二化螟体内不同种小分子量热激蛋白变异性较大(图1)。
表1 已鉴定的二化螟热激蛋白及基本特征
Table 1 The identified heat shock proteins in Chilo suppressalis and their basic characteristics
图1 用邻接法构建的水稻二化螟不同热激蛋白的系统发育树
Fig.1 Neighbour-joining phylogenetic tree of different heat
shock proteins of Chilo suppressalis
2 热激蛋白在二化螟生长发育中的调节作用
水稻二化螟是完全变态昆虫,分为卵、幼虫、蛹和成虫4种形态完全不同的虫态。在二化螟不同的生长发育阶段,Cshsp90的表达水平没有显著差异,而Cshsp70在整个发育阶段变化显著。尽管其体内的不同种热激蛋白呈现不同的表达规律。但是Cshsp90,Cshsp60,Cshsp19.8,Cshsp21.4,Cshsp21.5和Cshsp21.7b在成虫的表达水平最高,Cshsp70和Cshsp21.7a在1龄幼虫表达水平达到最高。hsp90,hsp70和hsp60在雌性个体中的表达量高于雄性个体,无论是蛹期还是成虫期的Cshsp19.8,Cshsp21.4,Cshsp21.5,Cshsp21.7a和Cshsp21.7b的表达水平在雄性和雌性之间都存在差异[25, 30]。有研究表明,在昆虫的发育过程中热激蛋白的表达水平受到蜕皮激素的调节,在昆虫的身体重构和变态阶段会具有更高的热激蛋白表达水平[19, 31]。同时,在昆虫生长过程中,体内会有积累的活性氧,从而启动转录因子Foxo和JNK信号途径诱导热激蛋白保护昆虫正常生长[32]。
3 热激蛋白在二化螟滞育过程中的作用
水稻二化螟属兼性滞育昆虫,短光照是滞育诱导的关键因子,3龄幼虫是主要的感应光周期的敏感虫龄,它能以4~7龄幼虫滞育越冬[7, 33-34]。水稻二化螟在扬州地区进入滞育的时间在9月初,滞育解除的时间为1月底到2月初之间,最高的滞育强度是11月份,可以维持滞育状态长达5个月之久,在田间保持静止期长达3个月,并且热激蛋白(Cshsp90,Cshsp70和Cshsp60)的表达和水稻二化螟的滞育有着紧密的联系[7]。例如,水稻二化螟越冬幼虫体内的hsp90,hsp70和hsp60都在2011年1月上中旬达到最大值,并且这3种热激蛋白的含量在2011年1月中旬以后都呈下降趋势[7]。同时,不同种类的热激蛋白在不同的地理种群水稻二化螟滞育幼虫体内的表达量没有显著性差异[7]。热激蛋白可能在二化螟滞育过程中起到保护自己的结构、调节和酶类蛋白的作用,从而保证滞育解除后,这些蛋白能够保证二化螟的进一步发育。
4 二化螟热激蛋白对温度的响应
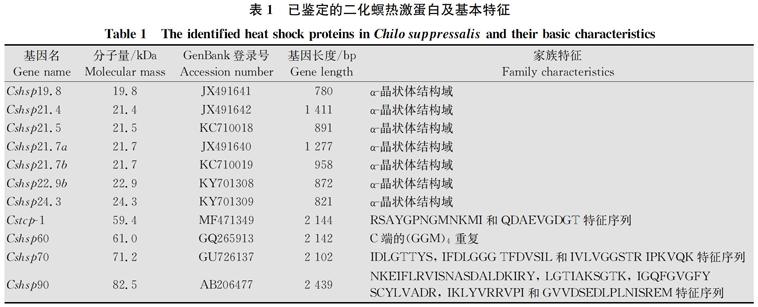
提及滞育,总是要和昆虫的耐寒性联系在一起。昆虫的耐寒性与滞育的关系大体上分为三种类型:滞育与其耐寒性完全独立、滞育与耐寒性部分独立、滞育与其耐寒性紧密联系。研究表明,越冬期间的水稻二化螟可以耐受-21℃的低温[35],且扬州地区水稻二化螟的耐寒能力在1月份最强,此时二化螟已开始逐渐解除滞育[7],说明扬州二化螟种群的耐寒性不是完全依赖于滞育,它部分独立于其滞育[35]。此外,田间水稻二化螟卵块经过42℃,8 h处理后,仍然有75%的存活率;二化螟幼虫高温处理2 h的LT50(引起种群50%个体死亡的温度)是45.4℃;水稻二化螟雄成虫和雌成虫高温处理2 h的LT50分别为43.8℃和43.6℃[36]。二化螟如此高的温度耐受能力和其体内热激蛋白的表达也有着密切的联系。日本的研究者发现:Cshsp90可以被低温诱导,但是,Cshsp70和Cshsp19.8不能够被低温诱导[22-23]。我们对11种二化螟热激蛋白对温度的响应规律做了系统的分析,表明二化螟不同种类的热激蛋白的响应规律不尽相同,其中Cshsp19.8,Cshsp21.7b,Cshsp22.9b,Cshsp24.3,Cshsp90对高低温都有响应,但对高低温的响应程度不一致。Cshsp19.8,Cshsp21.7b和Cshsp24.3对高温有很强的响应,而对低温仅有中等程度的响应;Cshsp22.9b对高温有中等程度的响应,而对低温具有很强的响应;Cshsp21.5仅对低温有中等的响应,而对高温没有响应。Cshsp21.4,Cshsp21.7a,Cstcp-1,Cshsp60,Cshsp70对高低温都没有响应。总之,在这11种热激蛋白中,多数为高温响应型[25-27, 30]。此外,对这些热激蛋白基因组结构特征分析表明,一般具有内含子的热激蛋白基因对温度不响应或者响应不敏感,例如Cshsp21.4具有内含子,对高低温都不响应。温度诱导型的热激蛋白在二化螟应对不良温度时,彼此相互配合,共同发挥作用保护二化螟免受高低温伤害。
表2 不同二化螟热激蛋白对温度的响应
Table 2 The responses of different heat shock proteins of
Chilo suppressalis to temperature
5 问题与展望
尽管已经对二化螟的热激蛋白进行了较为系统的研究,但依据转录组数据分析表明,水稻二化螟体内仍然还有其他热激蛋白的存在,因此还需要进一步分析鉴定其他的热激蛋白基因。同时,作为分子伴侣,水稻二化螟的许多生命活动都需要热激蛋白的参与,那么,每一种热激蛋白在不同的生命活动中其所起的作用是什么?例如,在生长发育中到底哪些热激蛋白起到主要的作用;在温度响应中哪一些热激蛋白起到主要的作用。针对其他的不良环境,如高湿、干燥、缺氧、饥饿等[37],水稻二化螟又是如何调节热激蛋白进行响应的?这些都是我们需要进一步研究的重要科学问题。由于研究昆虫热激蛋白还有很多技术障碍,目前水稻二化螟的热激蛋白研究还主要集中在基因水平。因此,解决技术难题,将水稻二化螟热激蛋白的研究逐步深入到蛋白質水平,以及研究二化螟热激蛋白表达的基因网络调控路径,是今后的主要研究方向。随着研究的不断深入,水稻二化螟的热激蛋白功能也会逐渐被揭示,这将为未来利用新的分子生物学技术防治该虫提供基础。
参考文献
[1] KHAN Z R, LITSINGER J A, BARRION A T, et al. World bibliography of rice stem borers 1794-1990 [M]. International Rice Research Institute, 1991.
[2] 刘勇,沈田辉,王风良,等.2013-2017年大丰东部沿海地区二化螟重发原因及绿色防控技术[J].现代农业科技,2018(7):146-147.
[3] 潘战胜,乐丽红, 江红梅. 余干县水稻二化螟发生特点及回升原因[J]. 江西农业, 2017(5): 102-104.
[4] 周斌, 刘洋, 黄思娣, 等. 湘南水稻二化螟发生现状及防治对策[J]. 湖南农业科学, 2017(12): 81-84.
[5] 孙莲, 赖朝晖, 蒋天梅, 等. 近13年象山县水稻二化螟发生情况及防控对策[J]. 浙江农业科学, 2018, 59(3): 494-495.
[6] 林炜, 韩永强, 刘玉娣, 等. 二化螟越冬生物学研究进展[J]. 湖南农业科学, 2008(3): 116-118.
[7] LU Mingxing, CAO Shuangshuang, DU Yuzhou, et al. Diapause, signal and molecular characteristics of overwintering Chilo suppressalis (Insecta: Lepidoptera: Pyralidae)[J]. Scientific Reports, 2013, 3(11):3211.
[8] 叶波, 李生智, 丁文兵, 等. 湖南不同二化螟种群对几种杀虫剂的抗性监测[J]. 湖南农业科学, 2017(9): 59-62.
[9] 赵丹丹,周丽琪,张帅,等.二化螟对双酰胺类杀虫剂的抗药性监测和交互抗性研究[J].中国水稻科学,2017,31(3):307-314.
[10] HUANG Shuijin, CHEN Qiong, QIN Wenjing, et al. Resistance monitoring of four insecticides and a description of an artificial diet incorporation method for Chilo suppressalis (Lepidoptera: Crambidae)[J].Journal of Economic Entomology, 2017, 110(6).
[11] RITOSSA F. A new puffing pattern induced by temperature shock and DNP in drosophila [J]. Experientia, 1962, 18(12): 571-573.
[12] TISSIRES A, MITCHELL H K, TRACY U M. Protein synthesis in salivary glands of Drosophila melanogaster: Relation to chromosome puffs [J]. Journal of Molecular Biology, 1974, 84(3): 389-398.
[13] KIM K K, KIM R, KIM S H. Crystal structure of a small heat-shock protein [J]. Nature, 1998, 394(6693): 595.
[14] HOFFMANN A A, SRENSEN J G, LOESCHCKE V. Adaptation of Drosophila, to temperature extremes: bringing together quantitative and molecular approaches[J]. Journal of Thermal Biology, 2003, 28(3): 175-216.
[15] DAHLGAARD J, LOESCHCKE V, MICHALAK P, et al. Induced thermotolerance and associated expression of the heat-shock protein Hsp70 in adult Drosophila melanogaster[J]. Functional Ecology, 1998, 12(5): 786-793.
[16] WANG H, DONG S Z, LI K, et al. A heat shock cognate 70 gene in the endoparasitoid, Pteromalus puparum, and its expression in relation to thermal stress [J]. BMB Reports, 2008, 41(5): 388.
[17] BAHRNDORFF S, MARIN J, LOESCHCKE V, et al. Dynamics of heat-induced thermal stress resistance and Hsp 70 expression in the springtail, Orchesella cincta[J]. Functional Ecology, 2009, 23(2): 233-239.
[18] JU R T, LUO Q Q, GAO L, et al. Identification of HSP70 gene in Corythucha ciliata and its expression profiles under laboratory and field thermal conditions [J]. Cell Stress & Chaperones, 2017, 23(2): 1-7.
[19] LIU Z, YAO P, GUO X, et al. Two small heat shock protein genes in Apis cerana cerana: characterization, regulation, and developmental expression [J].Gene,2014,545(2):205-214.
[20] ZHANG Yuanying, LIU Yaling, GUO Xulei, et al. sHsp22.6, an intronless small heat shock protein gene, is involved in stress defence and development in Apis cerana cerana[J]. Insect Biochemistry & Molecular Biology, 2014, 53(2): 1-12.
[21] RINEHART J P, LI A, YOCUM G D, et al. Up-regulation of heat shock proteins is essential for cold survival during insect diapause [J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(27): 11130.
[22] SONODA S, FUKUMOTO K, IZUMI Y, et al. A small HSP gene is not responsible for diapause and cold tolerance acquisition in Chilo suppressalis [J]. Journal of Applied Entomology, 2006, 130(5): 309-313.
[23] SONODA S, FUKUMOTO K, IZUMI Y, et al. Cloning of heat shock protein genes (hsp90 and hsc70) and their expression during larval diapause and cold tolerance acquisition in the rice stem borer, Chilo suppressalis Walker [J]. Archives of Insect Biochemistry & Physiology, 2006, 63(1):36-47.
[24] CUI Yadong, DU Yuzhou, LU Mingxing, et al. Cloning of the heat shock protein 60 gene from the stem borer,Chilo suppressalis, and analysis of expression characteristics under heat stress [J]. Journal of Insect Science, 2010, 10(10): 100.
[25] LU Mingxing, HUA Jin, CUI Yadong, et al. Five small heat shock protein genes from Chilo suppressalis: characteristics of gene, genomic organization, structural analysis, and transcription profiles [J]. Cell Stress & Chaperones, 2014, 19(1): 91-104.
[26] PAN D D, LU M X, LI Q Y, et al. Characteristics and expression of genes encoding two small heat shock protein genes lacking introns from Chilo suppressalis[J]. Cell Stress & Chaperones, 2018, 23(1):55-64.
[27] YU Tongying, LU Mingxing, CUI Yadong. Characterization of T-complex polypeptide 1 (TCP-1) from the Chilo suppressalis, HSP60 family and its expression in response to temperature stress [J].Journal of Integrative Agriculture,2018,17(5):1032-1039.
[28] 崔亚东, 杜予州, 陆明星, 等. 热胁迫对二化螟幼虫血淋巴细胞内活性氧、HSP90及细胞凋亡的影响[J]. 昆虫学报, 2010, 53(7): 721-726.
[29] 崔亞东,陆明星,杜予州.二化螟热休克蛋白70基因的克隆及热胁迫下的表达分析[J].昆虫学报,2010,53(8): 841-848.
[30] LU Mingxing, LIU Zhengxian, CUI Yadong, et al. Expression patterns of three heat shock proteins in Chilo suppressalis (Lepidoptera: Pyralidae)[J]. Annals of the Entomological Society of America, 2014, 107(3): 667-673.
[31] LUO Y, AMIN J, VOELLMY R. Ecdysterone receptor is a sequence-specific transcription factor involved in the developmental regulation of heat shock genes [J].Molecular & Cellular Biology, 1991, 11(7): 3660-3675.
[32] TOWER J. Heat shock proteins and Drosophila aging[J]. Experimental Gerontology, 2011, 46(5): 355-362.
[33] 汪信庚, 程家安. 二化螟滞育的研究[J]. 浙江大学学报(农业与生命科学版), 1993, 19(2): 170-174.
[34] XIAO Haijun, MOU Fengchen, ZHU Xingfen, et al. Diapause induction, maintenance and termination in the rice stem borer Chilo suppressalis (Walker)[J].Journal of Insect Physiology,2010,56(11): 1558-1564.
[35] LU Mingxing, LIU Zhongxian, WANG Xun, et al. Seasonal cold tolerance of Chilo suppressalis (Lepidoptera: Pyralidae)[J]. Annals of the Entomological Society of America, 2012, 105(3): 479-483.
[36] LU M X,CAO S S, LIU Z X, et al. Heat tolerance of developmental and seasonal stages of Chilo suppressalis [J]. Entomologia Experimentalis et Applicata, 2014, 152(2): 91-99.
[37] KING A M, MACRAE T H. Insect heat shock proteins during stress and diapause [J]. Annual Review of Entomology, 2015, 60(1): 59.
(责任编辑: 田 喆)
