草地螟迁飞的原因、目标与对策
2018-12-05罗礼智程云霞唐继洪江幸福张蕾
罗礼智 程云霞 唐继洪 江幸福 张蕾
摘要
人们对36°~55°N地区的重大农业害虫草地螟Loxostege sticticalis L.(鳞翅目:草螟科)的迁飞规律已了解较多,但对其迁飞的原因、目标与对策所知甚少。为了弥补这些缺陷,本文根据草地螟生活史特征,结合迁出、迁入以及再迁飞种群,成虫生殖,后代存活,田间幼虫危害种群发生与温湿条件的关系分析,明确了:1)迁飞种群主要降落在21℃,RH>60%的地区,较高或较低的温湿度是导致成虫迁出或再迁飞的原因;2)最大的成虫生殖能力及后代存活率,以及田间幼虫为害或大发生种群主要在21℃,RH>60%条件下形成;3)成虫为到达21℃,RH>60%的地区所采取的对策:先迁飞、后生殖,以及在迁飞过程中表现出的各种适应性行为等。所得结果为深化草地螟迁飞规律的认识、研究与应用提供了重要依据。
关键词
草地螟; 迁飞; 生殖与存活; 危害种群; 温湿度
中图分类号:
S 433.4
文献标识码: A
DOI: 10.16688/j.zwbh.2018338
The causes, targets and strategies of migration in the beet webworm,
Loxostege sticticalis (Lepidoptera: Crambidae)
LUO Lizhi CHENG Yunxia TANG Jihong JIANG Xingfu ZHANG Lei1
(1. State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection,
Chinese Academy of Agricultural Sciences, Beijing 100193, China; 2. Environmental and Plant
Protection Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou 571101, China)
Abstract
The beet webworm, Loxostege sticticalis L. (Lepidoptera: Crambidae) is a well-known cosmopolitan insect pest of crop and fodder plants in many regions between 36°-55°N latitude, but the causes, targets and strategies of its migration were less understood. Based on analyses of the life cycle, immigration, emigration and re-emigration, reproduction, survival and the relationships between occurrence of larval population and temperature and humidity, this review indicates that: (i) migrants mostly land on the areas with the conditions of 21℃, >RH 60%, too high or too low temperature and humidity are the major causes for emigration and re-emigration; (ii) the maximum fecundity and offspring survivorship and the most severe damage or outbreak are observed under the conditions of 21℃, >RH 60%; (iii) the migratory strategies taken by adult migrants are: migration is followed by reproduction, and various adaptive behaviors during migration. These results enhance our recognition, research and application of migration in this species.
Key words
Loxostege sticticalis; migration; reproduction and survival; outbreak population; temperature and humidity
草地螟Loxostege sticticalis L.(鱗翅目:草螟科)是一种温带昆虫,主要在北纬36°~55°N之间的地区,如我国华北、东北和西北发生为害。由于这些地区地貌复杂,不同海拔高度、季节和发生为害区间的温湿条件以及降雨量等变化很大[1-3]。另外,草地螟的繁殖、生长发育、存活等对温湿条件的要求十分严格[3]。但是,草地螟不仅能在这些地区生存和繁衍,而且能经常暴发成灾并造成重大经济损失[4-8]。那么,草地螟能在如此恶劣、多变的环境中生存繁殖或暴发成灾的对策是什么?已有的研究结果表明,是迁飞以及其他有效的生存对策。通过迁飞,草地螟可以逃离不利环境因素的影响而找到适宜的生境,可以使其生殖潜力得到最大程度的释放并使后代的生存成为可能,可以保持种群间的基因交流,而避免地方种群的产生[9-21]。的确,在成虫繁殖季节,草地螟迁飞现象无时不在。
经过长期的研究探索,草地螟迁飞规律的研究已取得了很大的进展:1)与95%的迁飞昆虫一样,草地螟属于兼性迁飞昆虫。即其迁飞行为的形成会受到温湿度[22]、幼虫密度[23-24]或滞育状态[25]、蛾龄[26-27]、生殖状态[28-29]等环境生理因子的调控,在不利的环境条件下形成的成虫迁飞倾向较大,反之,较小。2)迁飞行为的发生既与蛾龄、卵巢发育或生殖状态[9-10,24]有关,也与温湿度、
气流、降雨等气候因子密切相关[15-18,30-31]。另外,成虫在迁飞过程中会表现出各种主动行为来适应气候因子的变化[16-18,20]。3)成虫具有较强的迁飞能力:在室内,可以连续飞行24 h,飞行距离可超100 km[26];在田间,成虫的迁飞距离大多为数百公里,最远的可达1 200 km [9-10,13,32]。4)在我国,华北越冬成虫主要向东北地区迁飞[15,18-20,32],而东北一代成虫则可向华北地区迁飞[6, 21,33-34]。此外,草地螟还可进行垂直迁飞[13]。5)除新疆以外,我国草地螟各个发生为害区的虫源关系可互为虫源地[12,19, 35],即在此地发生的成虫可以迁飞到他地产卵为害,反之亦然。主要证据是:在东西6个省区,最大直线距离为1 582 km之内的11个越冬代成虫和一代成虫种群的176头成虫之间没有明显的遗传分化[19,35]。
值得重视的是,虽然草地螟迁飞研究已取得了很大进展,但由于人们对草地螟迁飞的原因、目的及对策关注甚少,困惑人们的问题依然不少。主要的有:1)草地螟可以滞育幼虫在其发生为害区平安越冬[2,12-13],那么成虫为什么还要迁飞?另外,在华北越冬虫源区或东北一代为害区,成虫羽化后通常会迁离本地,不在当地产卵,而是迁入到其他地区产卵为害[12-13,15, 36-37,39]。更有一些地区成虫迁入后很快又迁出[21,28,37-38],那么,促使成虫迁离羽化场所或再迁飞的原因又是什么?成虫对迁入场所的选择是否有要求,如果有,条件是什么?2)虽然都知道幼虫发生为害程度与成虫迁入(发生)量密切相关[13-14,36,38],但为什么在一些成虫迁入区幼虫为害很重甚至大发生,而在另外的迁入区,即使成虫迁入数量很大也没有幼虫为害[36,38],这些现象产生的原因或条件是什么?成虫对迁入场所的选择与成虫生殖及后代存活有没有关系? 3)草地螟的迁飞过程漫长、艰难而复杂[13,15,32,40]。那么,成虫是怎样或采取那些对策来实现其迁飞目标的,这些对策与成虫生殖状态有无关系,是否会影响成虫的生殖能力,以及成虫在迁飞过程中表现出的各种行为[16-18,20]是否有助于其迁飞目标的实现同样不清楚。
为了完善对草地螟迁飞规律的认识,并为其研究与应用提供更多的依据,本文以草地螟生活史特征为基础,以对其种群动态规律影响较大的温湿条件[1-3]为主线,在探明羽化迁出、再迁飞以及降落场所的温湿条件(本文以21℃,RH>60%为标准,在提到“较低”时,指温湿度均低于这个标准;“较高”时,仅指温度)并明确迁飞目的地的同时,进一步比较成虫生殖、后代存活以及田间种群形成与温湿度的关系,从而确定成虫迁飞的最终目的。最后,就成虫为实现迁飞目标所采取的对策进行了探索。现将主要结果报告如下。
1 成虫迁飞的原因与目标
虽然已知草地螟幼虫所经历的种群密度[23-24]和温湿条件[22]均可诱导成虫迁飞,但成虫羽化后所处的温湿条件是否会导致其迁飞并不十分明确。这里,我们尝试比较成虫羽化、再迁飞以及成虫迁入场所的温湿条件差异,以探明草地螟迁飞的原因与目标。
1.1 羽化迁出种群
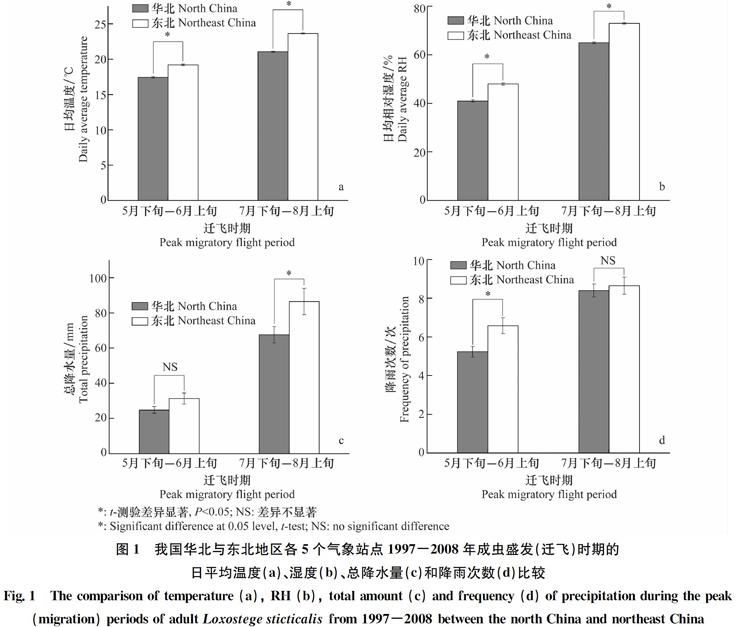
草地螟在我国主要发生为害区每年发生2代[2],大规模迁飞活动有2次。依次为发生于5月下旬到6月上旬,以及7月下旬至8月上旬的越冬代和一代成虫迁飞[6, 18-21,33-34],尽管在其他时期也存在着迁飞现象[4,13,38]。由于季节和地理条件(经纬度和海拔高度)的差异,在大多数地区和季节,成虫羽化时的温湿条件不是过低,就是过高(图1),因而成虫羽化后,大都迁离羽化场所,而不在本地产卵。
1.1.1 越冬成虫
在我国,草地螟主要在38°~ 43°N,108°~118°E的地区,如山西雁北、内蒙古乌兰察布盟和河北張家口等地越冬[12]。蛹和成虫的发育起点温度均为12℃[2],即在土表温度达到12℃若干时间段后,成虫即可羽化。但由于海拔较高(平均约1 300 m),气温上升较慢,再加上降雨较少,因此,成虫羽化时的温湿度通常较低(图1)。例如,在河北康保和内蒙古四子王旗,越冬成虫的始见期为5月13日前后7 d(罗礼智未发表),但康保此时的气温平均仅有10.3℃,湿度仅有44%。另据一些相关的报道,越冬成虫出现时的温度较低:如12.5~15℃[10]和15~16℃[16];陈阳等[32]在内蒙古察右中旗和武川县进行成虫标记释放试验期间的温度分别为14.8℃和16.6℃。而更重要的是,在这些条件下羽化的成虫会突然消失或减少,不在本地产卵[13,15,30,39]。而在其他温湿条件较高的地区,如东北等,通常在华北越冬成虫高峰期出现的2~4 d后成虫数量突然增加[15,30,39]。这似乎与此时东北地区的温湿度和降雨次数较高(图1)有关。
图1 我国华北与东北地区各5个气象站点1997-2008年成虫盛发(迁飞)时期的
日平均温度(a)、湿度(b)、总降水量(c)和降雨次数(d)比较
Fig.1 The comparison of temperature (a), RH (b), total amount (c) and frequency (d) of precipitation during the peak
(migration) periods of adult Loxostege sticticalis from 1997-2008 between the north China and northeast China
1.1.2 一代成虫
与越冬成虫相反,一代成虫大多于7月下旬-8月上旬羽化。因此,成虫羽化时所处的气温通常较高,尤其在低海拔(如东北)地区(图1a)。例如, 1982年8月1-4日,内蒙古扎赉特旗成虫羽化期间平均气温28.2~30.0℃,RH 61%~72%[21]。由于从高温条件下发育而成的成虫迁飞能力较强[22], 另外,这样的温濕条件也不利于成虫生殖(见2),因此,东北一代成虫羽化后迁出或不在本地产卵便成了我国二代幼虫为害通常轻于一代以及在当地很少找到越冬虫源的主要原因[12,14]。与此相对应的是,在华北越冬虫源区,此时的平均温度为21.1℃,RH 65%(图1a,b),从而较适合成虫生殖和后代存活(见2)。这也是我国越冬虫源区主要在华北地区形成的原因。在2003和2008年,一代成虫大量迁入到华北地区并形成大发生种群[5-7,33] 便是最好的例证。据称2008年的成虫是源自与东北相邻的蒙古国和俄罗斯 [6-7]。原因是成虫羽化后温湿条件不适宜[21]。
1.2 迁入种群对温湿条件的选择
与迁出种群不同的是,迁飞成虫通常降落在21℃左右, RH>60%的场所。例如,在1979-1983年,山西雁北地区越冬成虫产卵高峰期的平均温度20.5℃,RH 70%左右[13]。在河北康保1997年越冬成虫迁入高峰期(6月20-28日),平均气温20.8℃、RH 55%,两晚有小雨[4,38]。新疆阿勒泰地区2008年5月15-20日7县市有成虫降落,期间平均气温20.2℃,4县市有降雨(罗礼智未发表);在陈阳等[32]回收到的10头标记释放成虫中,回收地点平均温度18.4℃,其中6头回收地点温度在18.2~21.8℃之间;平均RH 65%;9头在回收前的12 h当地有降雨[40]。2012年6月17-18日,成虫迁入河北康保时的平均气温为20.9℃,但因湿度较低(42%)而迁出[28]。
一代成虫迁入的温湿条件与越冬成虫相一致。2003年大量一代成虫迁入晋北、冀北、蒙中西和陕北。在成虫迁入大同市期间(8月上旬),平均温度为21.9℃, RH 71%,降雨量28.2 mm,在迁入集宁、四子王旗和呼和浩特期间(7月27日-8月7日)的平均温度20.7℃,RH 62%,三地平均降雨量56.7 mm(罗礼智待发表),造成了这些地区二代幼虫大发生[5]。2006年8月1-10日,一代成虫迁入内蒙古四子王旗,期间平均温度20.8℃,RH 63%,降雨2次,共7.5 mm,二代幼虫发生面积约3.5万hm2(罗礼智待发表)。对我国2008年一代成虫迁入量与温湿条件进行分析的结果表明,成虫在21℃左右,RH 60%~80%条件下降落的频次最高,数量最大[33]。其中河北康保、内蒙古呼和浩特成虫迁入高峰期(8月上旬)的平均温度分别为20.6℃和21.9℃[7],内蒙古四子王旗7月31日降雨36.3mm,8月1-3日单灯诱蛾量超10万头,期间平均温度20.9℃、RH 63%(罗礼智未发表数据)。
成虫迁入与降雨也密切相关:1979-1985年,山西雁北地区10个测报站出现的70次蛾峰日中,有降雨的占88.4%(62/70)[13];内蒙古扎兰屯1980-1982年越冬成虫迁入高峰日均有降雨[41];1982-1983年6月,我国越冬成虫数量突增的15个点中,11个有降雨[15]。
1.3 再迁出种群
虽然国外学者[9-10]认为,草地螟迁飞降落后,一般不会再迁飞。但在我国,成虫再迁飞现象较常见[21,28,37-38]。在河北康保,已确认的再迁飞现象有3次,依次发生在1999年6月4日,2004年5月24日和2012年6月19日。第一和第三例迁入成虫滞留的时间为2 d(6月2-4日,6月16-18日),第二次成虫滞留的时间是1 d。成虫再迁出的原因是成虫迁入后温湿度较低,如第一次的温度为15.7~17.3℃,RH 26%~43%[38],第二次的温度为18.8℃,RH 39%,第三次则是温度合适,但湿度过低[28]。最后,前两个再迁飞种群的迁飞距离和命运不清,而第三个再迁飞种群降落在该县有灌溉条件的王红礼村,距离约10 km[28]。这些证据表明,成虫迁入后如果温湿度较低,成虫便会再次迁出并寻找适宜的温湿条件。
2 成虫生殖、后代存活以及危害种群形成的温湿条件
上述结果表明,迁飞成虫主要选择在21℃、RH>60%的地区降落,而迁出或再迁出种群所在的温湿条件不是过高,就是过低。那么,成虫历尽千辛万苦寻找这些条件的原因是什么?这些条件对于成虫生殖和后代存活到底有多重要?
2.1 成虫生殖
交配能力:草地螟行两性生殖,即雌蛾需要成功交配后方能产卵。因此,交配率和交配次数是影响成虫产卵和孵化的关键因子:在14~34℃和RH 70%条件下,交配率和交配次数均以22~26℃条件下的最高,当温度高于或低于这个范围时,这些参数均明显下降[42]。另外,成虫开始交配以及交配持续时间也会受到温度的影响[42]。在不同温湿条件下,成虫交配次数以22~26℃,RH 60%~80%条件下的较高,在其他温湿条件下的较低[22]。
产卵量:成虫产卵量与温湿度也有密切关系。湿度相同、温度不同时,产卵量以22℃条件下的最大,在温度超出这个数值时,产卵量便会下降[14,42-43];在温度相同、湿度不同时,产卵量以RH 60%~80%条件下的最高。同样,当湿度偏离这个范围时,产卵量便会下降[22,43-44]。在田间,如果在成虫盛发期遇到干旱、炎热或多雨天气,雌蛾的卵巢管会黏合在一起,卵子不能正常排出而引起卵巢管大小不一,卵巢管腐烂,卵巢退化而使雌蛾不孕并死亡。例如,在30℃、RH40%的条件下,就会破坏卵黄蛋白的形成,继而使卵子产生自溶现象[45]。另外,成虫的抱卵量与环境湿度关系密切。当RH>60%时,成虫的抱卵量在80%~90%之间,而在RH<43%时,卵巢不能发育[46]。
2.2 后代存活
孵化率:孵化率与温湿度密切相关。在10~14℃条件下,孵化率随温度的升高而增加,其中12℃和14℃条件下的存活率分别为10℃的39.5倍和47.6倍[2];在30℃条件下,孵化率会显著低于18~26℃的[47]。虽然有人认为湿度对孵化率没有影响[44],但不同温湿组合条件下的研究结果表明,在22℃、RH 60%~80%条件下的孵化率最高,在30℃、RH 20%條件下的最低[47]。
幼虫存活:与其他鳞翅类昆虫[48]一样,草地螟低龄幼虫的存活受温湿度[2,47]和食物[49]的影响最大。但由于藜等适宜寄主遍布于草地螟发生危害区。再加上成虫和幼虫均具有主动选择藜产卵和取食的能力,因此,只要温湿度适宜,食物不是影响幼虫存活的关键[1]:在10~14℃条件下,1龄幼虫存活率极低[2]。在18~30℃,RH 20%~100%条件下,1~2龄的存活率以22℃、RH 60%~80%条件的最高,30℃、RH 20%条件的最低。但3~4龄存活不受温湿度影响,5龄存活仅受湿度影响[47]。因此,温湿度是影响卵和低龄幼虫存活的关键因子。
2.3 田间危害种群形成
温湿度条件对草地螟生殖和存活的影响作用不仅体现在室内,而且更体现在田间。在我国,一代幼虫的发生为害程度主要取决于成虫发生量和盛发期间的温湿度条件。当成虫发生量大,盛发期的温度在21℃左右,RH≥60%时,幼虫就会大发生,如张家口坝上地区[4,38,46,50]、山西雁北地区[13]、东北(内蒙古东部、吉林和黑龙江中西部)地区[14,36-37,41,51-53]。另外, 二代幼虫发生为害的条件也与一代幼虫一样。如2003年[5]和2008年的二代[7,33]。但是,若成虫迁入后温湿度条件不适宜,即使成虫数量很大,也很难看到幼虫为害。如1983年6月上旬大量越冬成虫迁入东北地区,其中吉林省梨树县6月2日晚一台黑光灯诱蛾量超12万头。但是,由于成虫迁入后降雨量大,温度较低(6月均温19℃,比大发生的1982年同期低了3℃),因而极少有幼虫为害[36]。同样的情况也出现在东北其他地区[14,41,53]。2003和2008年,一代成虫在7月底和8月上旬大量迁入到晋北、冀北和蒙中西地区,但是,二代幼虫的大发生仅局限在21℃,RH>60%的地区[5,7,33],而在其他温湿条件不适宜的地区并没有幼虫危害种群发生。例如,2003年8月2-8日河北康保单灯诱蛾量达2.3万头,但由于期间的温度平均仅19℃,虽然RH 67%,二代幼虫很难发现(康爱国未发表)。又如2008年8月上旬,迁入到北京、河北南部、天津等地的成虫并没能形成幼虫危害种群,主要是由于气温较高(≥27℃)[7,33-34]。因此, 21℃、RH≥60%的地区,既是成虫迁飞的目的地(见1.2),更是幼虫为害或大发生种群形成的场所。
3 实现迁飞目标的对策
由于草地螟的迁飞是一个漫长而艰难的过程[13,21,32,40],从起飞、运转到降落的各个阶段都会受到气候因素的影响。为此,草地螟必须采取一些对策来提升本身的迁飞能力并适应天气条件的变化,从而增加到达目的地的可能性。从现有的结果来看,主要的对策有以下几方面。
3.1 生殖前迁飞
由于较长的产卵前期是成虫在产卵前找到适宜生境的保证,因此,迁飞蛾类通常都具有较长的产卵前期[11, 54-55]。草地螟在16~19℃条件下,产卵前期均>10 d, 最长为26 d;在22~28℃ 时, 为7 d左右[42-43]。较长的产卵前期对于其迁飞目标的实现意义重大:首先,草地螟的迁飞是一个漫长而艰难的过程[13,21,32,40],因此,成虫必须具备较强的飞行能力才能提升到达目的地的可能性。然而,成虫产卵后,体内飞行能源物质如甘油酯和糖类[22,29]含量显著下降,飞行肌已经降解、飞行能力显著下降[29]。其次,如若成虫产卵后再行迁飞,不仅其所产的后代会因温湿条件不适难以存活(见2.3),而且还会导致本身迁飞能力的丧失,因此,产卵后的迁飞既无可能,也没有意义。最后,其他相关结果也表明:成虫卵巢成熟后,群体迁飞活动不可能进行或显著下降[9-10];迁出或迁飞途中的雌蛾卵巢发育大多属于1~2级,交配率低,而在迁入区通常是3~4级,交配率高[12,17-20]:交配或开始产卵的田间种群虽然可以飞行,但飞行距离仅有10 km[28]。
3.2 适应性迁飞行为
起飞与迁飞高度:成虫羽化后便有一定的飞行能力,但要到3日龄才会迁飞[26-27],这可能与能源储备或飞行肌发育有关[11]。另外,成虫通常是在气温>15℃时起飞[9-10,16]。起飞时间主要在18:00左右或日落前后[9,16,26],主要是因为此时上升气流较强[9],有助于成虫用较少的能量上升到平流层;第三,成虫迁飞高度因季节、地区或风速的不同而发生变化[16-18,20],这也可能与节省飞行能源物质或提高飞行能源物质利用效益有关[22]。
定向:虽然草地螟迁飞方向主要取决于气流,但依然有一定的自主定向能力:在6月前后的迁飞通常是东北定向, 8月的则是西南定向,而在7月中旬则为随机定向[16-18]。由于我国主要越冬区5-6月上旬的温湿度较低,东北迁入区温湿度较高,而7-8月华北气温低于东北平原,较东北地区更接近草地螟适生温度条件(图1)。另外,越冬虫源区和东北迁入区5-6月份盛行的是西南风[15,20,40],而7-8月东北地区盛行的大多是西北风[6,21, 34],因而存在着由华北向东北往返迁飞的条件。因此,草地螟的这种定向机制使得迁飞种群较易找到适宜的温湿条件。
降落:无论是在室内还是室外,草地螟的降落活动通常是发生在凌晨4:00-5:00[9-10,17-18,20,26],尽管也有例外[21]。导致成虫凌晨降落的因素有低温(≤12℃)、阳光的出现、降雨、下沉气流和环境湿度的综合影响等[9-10]。成虫的降落大多发生在多云,阴雨的低压后部[10-11]。如当出现辐合带时;在锋面天气中,有下沉气流并有雨时;“槽前锋后”等[15,40]。因此,“随雨降落”可使迁飞种群较易到达湿度较高的生境(见1.2)。
3.3 成虫迁飞后生殖能力不受影响
虽然产卵后的成虫不能迁飞,但迁飞后的成虫生殖能力不受影响。Cheng等[27]模拟实际迁飞情况,对飞行与没有飞行过的成虫生殖能力进行研究的结果表明,飞行后的产卵前期、交配率、产卵量及孵化率与未飞行过的没有显著差异;飞行后成虫开始产卵整齐度显著提高。这些结果为后来的研究[56]所确认。这些结果表明:草地螟先迁飞、后生殖的对策是正确的,因为其迁飞的最终目的是寻找适于成虫生殖和后代存活的生境;成虫迁飞后的产卵前期不变和产卵整齐度显著提高也有重要的适应意义。前者可在成虫降落后遇到不适宜的温湿条件时,可再次迁飞(见1.3);而后者可使田间卵和幼虫种群密度瞬时增加,从而会提升幼虫的生存和取食为害能力并使为害加剧[24,27,57]。这或许就是东北迁入区和华北虫源区分别把单灯每晚诱捕到5 000头[14,36]和10 000头[13]作为当地草地螟大发生预测标准的原因。
4 讨论
通过对成虫迁出、迁入以及再迁飞场所(地区)温湿度条件的比较分析,明确了草地螟迁飞的诱因与目的:1)成虫羽化时的温湿度过低或过高是成虫迁飞的诱因。主要证据是:在华北越冬虫源区越冬成虫羽化时的温湿度通常较低(图1)。更重要的是,越冬成虫羽化后通常会迁飞到其他温湿度较高的地区如东北产卵为害[12-13,20,32];一代成虫主要在7月底8月初羽化,此时正是一年中温湿度较高的季节,尤其是在东北等低海拔地区(图1)。由于温度过高,成虫羽化后便在这些地区消失,从而构成了东北地区一代幼虫为害很重,二代幼虫为害很轻或越冬虫源很少的现象[12,36,51-52]。2)寻找21℃,RH>60%的地区是成虫迁飞的目的。无论是越冬还是一代成虫,迁飞成虫主要选择21℃,RH>60%的地区降落[4,7,33];另外,在成虫迁入场所的温度偏离21℃或RH<60%太多时,成虫还会再度迁飞[21,28,38]。需要强调的是,成虫在21℃,RH>60%条件下降落指的是迁入峰期的平均温湿度,单日(次)降落的温湿条件有时与上述指标相差较大,尤其是在有降雨或被迫降落的情况下。
为了阐明成虫主要选择21℃、RH>60%条件降落的原因,本文就温湿度条件对成虫生殖、后代存活以及田间幼虫危害种群形成的影响作用进行了分析。所得的结果惊人地相似:成虫生殖能力以及卵和1~2龄的存活均以在21℃、RH>60%条件下最大[2,14,42-43,47],田间幼虫危害或大发生种群似乎也只有在成虫盛发(迁入)期的温湿度条件处于21℃、RH>60%时才会形成[4,7,13,33]。为此,寻找21℃,RH>60%的生境既是成虫迁飞的目标,更是成虫生殖、后代存活以及田间幼虫危害种群形成的主要场所。如果这个结论正确的话,那么田间危害种群形成比室内种群所要求的温湿度条件更为苛刻。因此,加强对成虫迁入(盛发)期成虫发生量及温湿度条件的监测,就可预测当地幼虫的为害程度。另外,自新中国第一次猖獗为害周期以来,草地螟为害区的温度湿度[7,58-60]已经发生了较大的变化。因此,草地螟发生为害区有向纬度或海拔较高地区发展的趋势[59],这是今后草地螟虫情监测中应当注意的。值得提及的是,笔者也发现有幼虫危害种群是在成虫盛发期间温度高于21℃,或RH<60%的条件下形成的,如唐红艳等[53]的报道。这可能是由气象站获得的数据与田间为害场所的温湿度条件差异较大引起的,尤其是在田间为害场所与气象站距离或位置相差较大的情况下。因此,继续研究证实幼虫危害种群能否在成虫盛发期间温湿度条件较高或较低的情况下形成依然重要。
根据草地螟迁飞的目标,本文对其实现目标所采取的对策进行了探讨。所得的结果主要表现在两方面:1)先迁飞、后生殖:由于草地螟的迁飞是一个艰难而漫长的过程,而生殖后的成虫飞行肌降解,飞行能源物质及飞行能力显著下降[29]。因此,生殖后的成虫不可能完成迁飞。这也是成虫产卵前期较长[30,42]的主要原因。更重要的是,成虫迁飞后的生殖能力没有下降[27,51],因此,这个对策既避免了因生殖而造成的迁飞能力丧失,又可使到达目的地的成虫生殖能力和后代存活率显著提升,再加上产卵整齐度的提升,从而促进了种群的发展。2)在迁飞过程中表现出的各种行为也有很大的适应意义:如主动起飞、迁飞高度的变化,以及随风飞行等行为可能与节省飞行能源或提高飞行能源利用效益有关,季节性迁飞方向的改变可有助于成虫更好地利用盛行的气流找到适宜的温度条件,而随雨降落则有助于找到湿度较高的条件等。然而,由于影响草地螟迁飞的天气系统较多[15,40],草地螟迁飞过程中是否还有其他的适应对策尚待研究。
综上所述,通过本文的探索,虽然在草地螟迁飞原因、目标与对策等都取得了一些较有意义的结果,但是,由于笔者能力及其他条件的限制,笔者在讨论这些问题时,仅聚焦于现象分析,而对其产生的机理涉及较少;在探索迁飞原因与目的时,仅局限于温湿度的讨论,而对种群密度等其他环境因子的作用则未能探讨;在探索迁飞对策时,“目的”性也较强(见上段);没能就文中结果与其他相关研究结果进行比较,从而降低了本文应有的价值。因此,如何研究和解决这些问题,依然是草地螟迁飞研究所面临的最大挑战。
参考文献
[1] PEPPER J H. The effect of certain climatic factors on the distribution of the beet webworm (Loxostege sticticalis L.) in North America [J]. Ecology, 1938, 19: 565-571.
[2] 羅礼智,李光博.草地螟的有效积温及世代区的划分[J].昆虫学报,1993,36(3):332-339.
[3] 罗礼智,程云霞,唐继洪,等.温湿度是影响草地螟发生危害规律的关键因子[J].植物保护,2016,42(4):1-8.
[4] 羅礼智,张红杰,康爱国.张家口1997年一代草地螟幼虫大发生原因分析[J].自然灾害学报,1998,7(3):158-164.
[5] 罗礼智.我国2004年一代草地螟将暴发成灾[J].植物保护,2004,30(3):86-88.
[6] CHEN Xiao, ZENG Juan, ZHAI Baoping. A series of abnormal climatic conditions caused the most severe outbreak of first-generation adults of the meadow moth (Loxostege sticticalis L.) in China[J].International Journal of Biometeorology,2016,60:789-800.
[7] 罗礼智,黄绍哲,江幸福,等.我国2008年草地螟大发生特征及成因分析[J].植物保护,2009,35(1):27-33.
[8] 罗礼智,程云霞.草地螟[M]∥中国农业科学院植物保护研究所,中国植物保护学会.中国农作物病虫害(第三版中册).北京:中国农业出版社,2014:1441-1448.
[9] MELNICHENKO A N. Regularities of mass flying of the adult Loxostege sticticalis L. and the prognosis of their flight migrations [R]. Bulletin of Plant Protection, Leningrad Series 1, Entomology, 1935, No. 17:1-54.
[10] STRELNIKOV I D. The migrations of Loxostege sticticalis L.[J]. Bull. Inst. Sci. Leshaft, 1935, 19: 77-120.
[11] JOHNSON C G. Migration and dispersal of insects by flight[M]. London: Methuen, 1969.
[12] 全国草地草地螟科研协作组.草地螟(Loxostege sticticalis Linnaeus)发生及预测和防治的研究[J]. 病虫测报,1987(S1): 1-9.
[13] 张树坤,刘梅凤,李齐仁,等. 山西省草地螟发生规律、预测预报及其综合治理的研究[J]. 病虫测报,1987 (S1):82-97.
[14] 魏倩, 崔万里, 杜俊岭, 等. 黑龙江省草地螟(Loxostege sticticalis)发生规律、预测预报及综合防治研究(1981-1985年)[J]. 病虫测报,1987(S1): 98-107.
[15] 杨素钦,马桂椿.草地螟迁飞路径的探讨[J]. 病虫测报,1987(S1): 122-128.
[16] 陈瑞鹿,暴祥政,王素云, 等.草地螟迁飞活动的雷达观测[J]. 植物保护学报, 1992, 19(2): 171-174.
[17] FENG Hongqiang, WU Kongming, CHENG Dengfa. Spring migration and summer dispersal of Loxostege sticticalis (Lepidoptera: Pyralidae) and other insects with radar in northern China [J]. Environmental Entomology, 2004, 33(5): 1253-1265.
[18] 程登发,封洪强,吴孔明.扫描昆虫雷达与昆虫迁飞监测[M].北京:科学出版社,2005.
[19] 曹卫菊,罗礼智,徐建华.我国草地螟的迁飞规律及途径[J].昆虫知识,2006(3):279-283.
[20] 张云慧,陈林,程登发,等.草地螟2007年越冬代成虫迁飞行为研究与虫源分析[J].昆虫学报,2008,51(7):720-727.
[21] 陈晓.草地螟迁飞、越冬规律及暴发机制研究[D].南京:南京农业大学,2010.
[22] 唐继洪.草地螟对温湿度变异的适应与反应[D].北京:中国农业科学院,2016.
[23] KONG H L, LUO L Z, JIANG X F, et al. Effects of larval density on flight potential of the beet webworm, Loxostege sticticalis (Lepidoptera: Pyralidae)[J]. Environmental Entomology, 2010, 39(5):1579-1585.
[24] 杨志兰.草地螟生长发育、繁殖及飞行的集群优势及其生理原因[D]. 武汉:华中农业大学,2013.
[25] XIE D S, LUO L Z, SAPPINGTON T W, et al. Comparison of reproductive and flight capacity of beet webworm, Loxostege sticticalis (Lepidoptera: Pyralidae), developing from diapause and non-diapause larvae[J]. Environmental Entomology, 2012, 41(5):1199-1207.
[26] 罗礼智, 李光博. 草地螟不同蛾龄成虫飞行能力和行为的研究[C]∥中国生态学会青年研究会,中国昆虫学会昆虫生态专业委员会.青年生态学者论丛(二)昆虫生态学研究.北京:中国科学技术出版社,1992:303-308.
[27] CHENG Yunxia, LUO Lizhi, JIANG Xingfu, et al. Synchronized oviposition triggered by migratory flight intensifies larval outbreaks of beet webworm [J/OL]. PLoS ONE, 2012, 7(2): e31562. doi:10.1371/journal.pone.0031562.
[28] 程云霞,刘科科,康爱国,等.草地螟迁入和再迁飞种群的生理环境特征分析[J].植物保护,2013, 39(3):26-30.
[29] CHENG Yunxia, LUO Lizhi, SAPPINGTON T W, et al. Onset of oviposition triggers abrupt reduction in migratory flight behavior and flight muscle in the female beet webworm, Loxostege sticticalis [J/OL]. PLoS ONE, 2016, 11(11): e0166859. doi:10.1371/journal. pone.0166859.
[30] 羅礼智. 温度对草地螟(Loxostege sticticalis L.)的影响及其迁飞特性的研究[D]. 北京:中国农业科学院, 1985.
[31] 唐继洪,程云霞,罗礼智,等.蛾龄、温度和相对湿度对草地螟自主飞行能力的影响[J].植物保护,2016,42(2):79-83.
[32] 陈阳,姜玉英,刘家骧,等.标记回收法确认我国北方地区草地螟的迁飞[J].昆虫学报,2012,55(2):176-182.
[33] 黄绍哲,罗礼智,姜玉英,等.我国2008年草地螟大发生种群空间分布特征[J].植物保护,2011,37(4):76-81.
[34] 黄绍哲.我国草地螟(Loxostege sticticalis)种群时空动态规律研究[D].北京:中国农业科学院,2012.
[35] JIANG Xingfu, CAO Weiju, ZHANG Lei, et al. Beet webworm (Lepidoptera: Pyralidae) migration in China: evidence from genetic markers [J].Environmental Entomology,2010,39(1): 232- 242.
[36] 吉林省草地螟科研协作组.吉林省草地螟发生规律及防治技术研究报告[J].病虫测报,1987(S1):34-52.
[37] 岳宗岱,袁艺.吉林省草地螟虫源和发生条件的初步分析[J].吉林农业科学,1983(3):78-81.
[38] 康爱国,杨海珍,李强,等.草地螟越冬代虫量与第一代草地螟发生关系的研究[J].昆虫知识,2004,41(1):70-72.
[39] 罗礼智,李光博,曹雅忠.草地螟第3个猖獗为害周期已经来临[J].植物保护,1996,22(5):50-51.
[40] 孙虹雨.草地螟迁飞、降落与东北冷涡的关系[J].气象与环境学报,2014,30(6):85-91.
[41] 呼伦贝尔盟草地螟研究协作组.草地螟发生规律及预测预报和防治研究[J].病虫测报,1987(S1):70-82.
[42] CHENG Yunxia, WANG Kai, SAPPINGTON T W, et al. Response of reproductive traits and longevity of beet webworm to temperature, and implications for migration [J/OL]. Journal of Insect Science,2015,15(1):154.DOI:10.1093/jisesa/iev135.
[43] 罗礼智,李光博.温度对草地螟成虫产卵和寿命的影响[J].昆虫学报,1993,36(4):459-464.
[44] 孟正平,陈玉宝,刘一凌.草地螟生殖力及卵孵化与湿度的关系[J].病虫测报,1987(S1):115-120.
[45] OMELOVA B P. Forecasting population dynamics of grass moth (Loxostege sticticalis)[J]. Zashchita Rastenii,1982 (9):42-46.
[46] 田绍义.草地螟及其防治研究[J].河北农学报,1963,2(3):15-22.
[47] TANG J H, CHENG Y X, SAPPINGTON T W, et al. Egg hatch and survival and development of beet webworm (Lepidoptera: Crambidae) larvae at different combinations of temperature and relative humidity [J]. Journal of Economic Entomology, 2016, 109(4): 1603-1611.
[48] ZALUCKI M P, CLARKE A R, MALCOLM S B. Ecology and behavior of first instar larval Lepidoptera [J]. Annual Review of Entomology, 2002, 47:361-393.
[49] 尹姣,曹雅忠,羅礼智,等.寄主植物对草地螟种群增长的影响[J].植物保护学报,2004,31(2):173-178.
[50] 刘光涛,郭向东,张金,等.草地螟的生物学特性及其发生与条件环境的关系[J].病虫测报,1987(S1):59-64.
[51] 贾宗谊.黑龙江省草地螟发生规律与防治策略的探讨[J].黑龙江农业科学,1983(2):17-21.
[52] 屈西锋,邵振润,王建强.我国北方农牧区草地螟暴发周期特点及原因剖析[J].昆虫知识,1999,36(1):11-14.
[53] 唐红艳,牛宝亮.影响草地螟一代幼虫发生的关键气象因子及指标研究[J].中国农学通报,2010,26(18):270-274.
[54] GATEHOUSE A G, ZHANG X X. Migratory potentials in insects: variation in an uncertain environment[M]∥DRAKE V A, GATEHOUSE A G. Insect migration, tracking resources through space and time. Cambridge: University Press, 1995.
[55] 罗礼智.昆虫迁飞行为的发生与调控[J].科学,1998,50(3):32-35.
[56] 宋文一,程云霞,罗礼智,等.迁飞对草地螟生殖行为及种群暴发的影响[J].植物保护,2016,42(2):26-30.
[57] 孔海龙,罗礼智,江幸福,等.幼虫密度对草地螟食物利用率及消化酶活性的影响[J].昆虫学报,2012,55(3):361-366.
[58] IPCC. A contribution of working groups I, II, and III to the fourth assessment report of the Intergovernmental Panel on Climate Change [M]∥PACHAURI R K, REISINGER A. Climate Change 2007: Synthesis Report IPCC,Geneva,2007:1-22.
[59] 唐继洪,程云霞,罗礼智,等.基于Maxent模型的不同气候变化情景下我国草地螟越冬区预测[J].生态学报,2017,37(14):4852-4863.
[60] CHEN Xiao, JIANG Yuying, KANG Aiguo, et al. Drying soil in North China drove the outbreak range expansion of meadow moth by facilitating long-distance migration [J/OL]. Scientific Reports, 2016, 6:30370.
(责任编辑: 田 喆)
