文冠果花性别分化过程中形态与解剖结构特征和气象因子分析
2018-12-05苏淑钗刘觉非黄曜曜刘金凤张行杰
张 宁,敖 妍,2*,苏淑钗,2,刘觉非,黄曜曜,刘金凤,张行杰
(1 北京林业大学 省部共建森林培育与保护教育部重点实验室,北京 100083;2 国家能源非粮生物质原料研发中心,北京 100083;3 赤峰市林业种苗站,内蒙古赤峰 024000;4 胜利油田胜大生态林场,山东东营 257000)
文冠果(XanthocerassorbifoliumBunge)隶属无患子科(Sapindaceae)文冠果属(XanthocerasBunge),落叶小乔木或灌木,雌雄同株,具有耐寒、耐旱、耐瘠薄等特性,主要分布于中国北方地区[1]。其综合利用价值较高,在食品、化工、医药方面都已经开发利用,并具有较高的生态价值和观赏价值。文冠果结实早,结果期长,种仁油脂含量可达67%[2],种子油是很好的食用油,也可用作制备生物柴油。该树种已被国家林业局列为重要生物质能源树种,并于2014年发布了其原料林可持续培育指南。但生产中文冠果繁花少实,种子产量低,这是目前亟待解决的问题。文冠果为总状花序,顶芽花序上多着生雌能花,侧芽花序上多为雄能花,因而雌能花数量远少于雄能花[3]。研究雌能花和雄能花发育过程,确定性别分化时期是进行性别调控的基础,对促进雌能花分化,提高雌雄比例,进而大幅提高种子产量具有重要理论和实践意义。
植物性别分化是一个复杂的过程,气象条件、地理种源、栽培地点都可能影响该过程[4-9]。研究表明,气象因子尤其是温度能影响植物体内碳水化合物等营养物质的积累及激素分配,例如,高温会造成营养大量消耗,直接影响开花质量和花性别[10-14]。另外,植物生长期或某个发育期一般需要具有特定阈值的活动积温[15-16],温度会影响花芽分化的速率和进程,并影响花性别[17-19]。对于某些植物,例如胡麻、黄瓜等,适度低温利于雌花分化[11,20]。此外,湿度对植物的花芽分化和性别分化进程也有重要影响。适度的水分胁迫促进植物对碳水化合物的积累,有利于花芽分化;相反,较大空气湿度和低温以及光照程度不足等均会对花芽分化及开花产生影响[21-24]。土壤和空气相对湿度对雌雄花的比例有很大影响[25-30]。一定范围内,土壤持水量和空气湿度越大,越有利于雌花分化[11]。目前,国内外有关文冠果花芽发育的研究主要集中于分化过程观察[31-32]、雌雄蕊选择性败育[33-36]等,明确了文冠果花芽分化过程,以及雌雄蕊败育发生时期。但以往研究多是针对某地区某年份的研究,由于时间和空间限制,难以将其结论应用到其他地区或同一地区的其他年份。而且,对文冠果性别分化出现的时期及持续时间长短与气象因素关系的研究报道甚少。文冠果花芽分化和性别分化是生长发育过程中的关键阶段,是复杂的形态建成过程。其分化结果直接影响种实产量和品质的提高。研究分化过程关键期的内外形态特征及与气象条件的关系,对人为调控雌雄比例、提高种实产量有着重要作用。为此,本研究将雌能花和雄能花性别分化过程外部形态特点与内部解剖结构相对应,同时监测温度、湿度、降水等气象因子,明确了文冠果性别分化的关键期,掌握了该过程内外部形态特征的相关性,同时阐述了性别分化各阶段与气象因子的对应关系。研究结论可为不同地区文冠果依据花芽外部形态和气象数据推测内部分化进程、判断性别分化关键期提供参考,为适时采用栽培管理措施调控性别分化提供依据。
1 材料和方法
1.1 试验地概况
试验地位于山东省东营市东营区胜大林场,地处E118°12′42″~118°59′52″,N37°14′13″~37°31′57″,海拔8.8 m。属温带大陆性季风气候,年降水量550~600 mm,多集中在夏季。年平均气温约12.5 ℃, 1月均温0.06 ℃,7月均温28.31 ℃,≥10 ℃积温约4 300 ℃,无霜期长达206 d。
1.2 试验材料
样地内文冠果为当地起源的8年实生人工林。平均树高2.8 m,平均胸径13 cm,株行距3.0 m×3.0 m,树冠体积在4.5~5.0 m3之间[37]。选择林分内具有代表性的生长健壮、树势一致、无病虫害的文冠果植株10株作为观察和采样对象。
1.3 试验方法
(1)雌、雄能花分布观测:在每株样树东西南北方向各选择3个枝条,共120个枝条,挂牌标记,花开放后统计顶、侧花序上雌、雄能花数量。计算各自在顶侧花序上所占比例。
(2)性别分化观测:参考前人研究结果[32],于2017年2月21日至4月30日期间取样。其中,2月21日至3月31日,取顶、侧芽(花序),每5 d取样1次;4月1日至4月30日,取顶、侧花序上的单花,每3 d取样1次,直到雄能花散粉、雌能花可授时,共取样18次。分化初期花序未伸长时,每次采集样株树冠南向外围枝条的顶、侧芽,侧芽采集枝条形态学下端第3~5个花序。花序伸长后,为保持一致性,采集顶、侧花序形态学下端第6~7个花蕾。每次每棵树顶、侧花芽(蕾)各取5个,共50个芽。样品分别用于外部形态学和解剖学观察。
(3)形态学观察:对选取的顶、侧芽观察其外部形态特点,记录形状、颜色,用电子游标卡尺(上海首丰精密仪器有限公司)测量横、纵径,并拍照。
(4)解剖学观察:样品用FAA固定液固定,带回实验室,常规石蜡切片制作,番红-固绿染色,加拿大树胶封片,Olympus CX-41 光学显微镜观察并拍照记录[38]。
(5)气象因子观测:2017年2月21日至4月30日期间,进行形态学和解剖学观察的同时进行气象因子观测,主要监测样地的温、湿度数据。在试验地中部放置ARN-17温湿度自动记录仪(河北奥尔诺电子科技有限公司),记录地表以上20 cm处空气温、湿度,数据每隔1 h记录存储1次。降水数据由气象站获得,试验地无灌溉。
1.4 数据处理
文冠果生物学零度根据芽膨大前10 d平均气温公式法计算[39]。
K=∑T/N
(1)
式中:K表示生物学零度;T表示芽膨大前10 d平均温度(先计算每天平均温度值,再计算10 d平均值所得);N表示观测年份。
日平均气温高于生物学零度时起计算各时期开始时和各时期内的有效积温、平均温度和平均湿度[40]。
≥K有效积温∑T=∑(t-K)
(2)
≥5℃有效积温∑T=∑(t-5)
(3)
≥10℃有效积温∑T=∑(t-10)
(4)
式中:t表示日平均气温。(其中t-K、t-5、t-10均大于0,若小于等于0,以0计算)。
采用Excel 2013和SPSS13.0统计分析软件分析数据。
2 结果与分析
2.1 文冠果不同性别花的分布
表1为顶、侧花序上雌、雄能花的比例差异。可见,雌能花和雄能花的比例在顶、侧花序上存在极显著差异,顶花序上89.79%是雌能花,侧花序上91.04%是雄能花。雄能花在顶花序上分布的变异系数较大,但在侧花序上的变异系数仅为3.22%。雌能花在侧花序上的分布比例变异系数较大,为32.7%,但在顶花序上分布比例的变异系数仅为8.15%。总体来看,顶花序上雌能花的分布和侧花序上雄能花的分布是相对稳定的。因此,雌能花和雄能花样品分别采自顶花序和侧花序。
2.2 文冠果花性别分化过程外部形态与内部解剖特点
2.2.1性别分化早期外部形态与内部结构2月21日,文冠果花芽被褐色鳞片,侧芽横径3.98~4.87 mm、纵径3.32~4.12 mm,顶芽横径6.67~8.32 mm、纵径5.46~6.21 mm(图1,①)。内部雄蕊呈棒状体(图1,①a)。3月3日~3月18日,花芽开始萌动,芽呈圆锥形膨大,芽鳞错开,露出黄绿色伴有红色的芽体,侧芽横径4.33~6.63 mm、纵径5.61~8.75 mm,顶芽横径5.21~7.92 mm、纵径6.73~9.91 mm(图1,②③)。内部顶端生长点明显增大突起,雌蕊原基出现(图1,②a)。雄蕊原基细胞分裂较快,雄蕊伸长,顶端形成分生组织,出现幼嫩花药,花丝伸长,花粉囊出现(图1,③a)。3月18日~3月23日,芽继续膨大,鳞片浅绿色,芽体露出,顶部露出花蕾,侧芽横径5.13~8.34 mm、纵径9.36~12.35 mm,顶芽横径5.88~9.32 mm、纵径10.56~13.22 mm(图1,④⑤)。雌蕊继续膨大。花药呈蝶形,4室,左右基本对称(图1,④a),药室内处于造孢细胞时期,最内一层细胞体积较大,胞质浓厚(图1,④b)。随着周缘细胞的分裂,由外向内逐渐分化出花药壁,造孢细胞相应分裂形成体积较大、细胞核明显、细胞质浓厚、无明显液泡的花粉母细胞,即小孢子母细胞(图1,⑤a)。同时花药壁发育完全,由5层细胞组成,由外向内依次为表皮细胞、纤维层细胞、2层中间层细胞、绒粘层细胞(图1,⑤a)。

表1 文冠果不同性别花在顶侧花序分布比例差异
注:**表示在0.01水平差异显著。变异系数为标准差/均值×100%
Note:** indicating the significant difference at 1% level. The coefficient of variation is the ratio of the standard deviation to the mean multiplied by 100%
3月23日~4月2日,花序迅速伸长,鳞片脱落,露出绿色花蕾,侧花序横径8.38~10.01 mm、纵径13.03~21.17 mm,花蕾横径1.76~2.31 mm、纵径1.72~2.10 mm。顶花序横径8.92~10.38 mm、纵径15.76~20.03 mm,花蕾横径1.22~1.54 mm、纵径1.23~1.45 mm(图1,⑥⑦)。内部花粉囊的中层和绒粘层逐渐解体(图1,⑥a),小孢子母细胞彼此分离,进入减数分裂阶段(图1,⑥b)。随后进入小孢子四分体时期,其胞质分裂方式为同时型分裂,四分体的4个子细胞呈四面体排列(图1,⑦a)。出现子房、胚珠雏形(图1,⑦b),部分花出现实心子房(图1,⑦c),柱头伸长(图1,⑦d)。
2.2.2性别分化后期雄能花的形成4月3日~4月9日,雄能花花蕾增大至露白,横径2.05~4.54 mm、纵径2.99~5.32 mm(图2,①②)。内部子房和胚珠出现败育迹象,子房壁细胞不规则扭曲变形(图2,①a),胚珠内大孢子母细胞四分体出现异常(图2,①b)。花药绒粘层分泌胼胝质酶,将花粉四分体的胼胝质壁溶解,单核花粉粒从四分体释放出来,花粉粒的核位于细胞中央,具有浓厚的细胞质(图2,②a)。4月9日~4月12日,雄能花逐渐开放,横径6.89~8.42 mm、纵径5.24~7.33 mm(图2,③)。内部花粉粒细胞体积迅速增大,细胞质明显液泡化,形成中央大液泡,将细胞核挤到一侧,花粉发育进入单核靠边期(图2,③a)。接着进行一次有丝分裂,形成双核花粉粒(图2,③b)。4月12日~4月15日,花朵开放,横径21.20~25.31 mm、纵径14.57~17.65 mm,8枚雄蕊,雄蕊伸长速度快于雌蕊,看不到子房(图2,④⑤)。花粉粒进一步增大,整个细胞从花粉粒壁部脱离,可见3个萌发孔,花粉粒成熟(图2,④a)。子房完全败育,只剩下细胞壁团,胚珠停止发育并退化,柱头严重萎缩(图2,④b);前期出现实心子房的花子房干瘪,顶端萎缩,无花柱(图2,⑤a)。
2.2.3性别分化后期雌能花的形成4月3日~4月9日,雌能花花蕾横径1.83~2.92 mm、纵径2.01~3.12 mm(图3,①)。内部子房胚珠正常发育,柱头继续伸长(图3,①a),胚珠为双珠被,珠心内部体积大、细胞质浓厚的细胞为孢原细胞(图3,①b)。4月9日~4月12日,花蕾露白,横径3.58~7.29 mm、纵径3.86~7.69 mm(图3,②)。此时大孢子母细胞经两次减数分裂,发育为大孢子四分体,其中3个大孢子逐渐退化消失,只有1个大孢子可以发育为胚囊(即功能大孢子)(图3,②a)。4月12日~4月15日,花朵沿花序轴自下而上依次开放,花横径12.25~18.30 mm、纵径8.30~10.98 mm;子房横径2.02~2.72 mm、纵径2.22~3.20 mm,花柱长2.52~2.74 mm,花丝停止伸长,柱头高于雄蕊(图3,③④)。花药中单核花粉粒有丝分裂异常,液泡化衰败,看不到明显的细胞核(图3,③a)。内部功能大孢子发育,细胞体积逐渐增大,发育为单核胚囊(图3,③b)。之后连续进行3次有丝分裂,依次形成二核、四核、八核胚囊(图3,④a、④b、④c)。至4月18日,柱头发育为圆球形,子房膨大,花药干瘪(图3,⑤)。子房胚珠形态发育完全(图3,⑤a),中空花柱道,柱头开裂,乳突状细胞可见(图3,⑤b)。花药花粉囊没有形成唇细胞,不开裂(图3,⑤c),或花药绒粘层解体不完全,仅留1层萎缩的细胞质残迹,花粉败育,无萌发孔(图3,⑤d)。

①.2月21日花芽;②.3月3日花芽;③.3月8日花芽;④.3月13日花芽;⑤.3月23日花芽;⑥.3月28日花序;⑦.4月2日花序。①a.雄蕊棒状体;②a.雌蕊原基;③a.花药形成、花丝伸长;④a.雌蕊原基膨大、花粉囊形成;④b.造孢细胞时期;⑤a.小孢子母细胞、花药壁形成;⑥a.中层绒粘层解体;⑥b.小孢子母细胞分离;⑦a.四分体时期;⑦b.胚珠原基出现;⑦c.实心子房;⑦d.柱头伸长。Sp.雄蕊原基;Pp.雌蕊原基;A.花药; F.花丝;Sc.造孢细胞;MMc.小孢子母细胞;Ep.表皮;Fl.纤维层;Ml.中层;Ta.绒粘层;T.四面体型;Ova.子房;Ovu.胚珠;S.柱头。图①a、图①b意为该时期对应的外部形态为图①,下同图1 文冠果花性别分化早期外部形态与内部结构特点①. Flower buds on February 21; ②. Flower buds on March 3; ③. Flower buds on March 8; ④. Flower buds on March 13; ⑤. Flower buds on March 23; ⑥. Inflorescence on March 28; ⑦. Inflorescence on April 2. ①a. Stamens clavule; ②a. Pistil primordium; ③a. Anther formation stage and filament elongation stage; ④a. Pistil primordial enlargement and pollen sac formation stage; ④b. Spore cell stage; ⑤a. Microspore mother cells and anther wall formation stage; ⑥a. Middle cashmere layer disintegration; ⑥b. Microspore mother cell separation; ⑦a. Tetrad period; ⑦b. Ovule primordia appear; ⑦c. Solid child room; ⑦d. Stigma extension stage. Sp. Stamen primordium; Pp. Pistil primordium; A. Anther; F. Filaments; Sc. Sporogenous cells; MMc. Microspore mother cell; Ep. Epidermis; Fl. Fiber layer; Ml. Middle layer; Ta. Tapetum; T. Tetrahedral; Ova. Ovary; Ovu. Ovules; S. Stigma. Fig.①a and Fig.①b mean that the corresponding external appearance of this period is Fig.①, the same as belowFig.1 The external morphology and internal structural characteristics at the early stage of flower sex differentiation of X. sorbifolium

①.4月3日雄能花花蕾;②.4月6日雄能花;③.4月9日雄能花;④.4月12日雄能花;⑤.4月15日雄能花。①a.子房胚珠发生败育;①b.大孢子母细胞四分体;②a.单核花粉粒;③a.单核花粉靠边期;③b.双核花粉粒;④a.成熟花粉粒;④b.败育的子房胚珠;⑤a.萎缩的实心子房。S.柱头;Ovu.胚珠;Mc.大孢子母细胞;Mp.单核花粉粒;Up.单核靠边花粉;Vn.营养核;Rn.生殖核;P.花粉;Gp.萌发孔;Pp.雌蕊原基图2 文冠果性别分化后期雄能花形成过程中外部形态和内部解剖特点①. Male flower bud on April 3; ②. Male flower on April 6; ③. Male flower on April 9; ④. Male flower on April 12; ⑤. Male flower on April 15.①a. The ovary and ovules abortion occurred; ①b. The tetrad stage of megaspore mother cell; ②a. Mononuclear pollen grains; ③a. Mononuclear pollen by the edge; ③b. Binuclear pollen grains; ④a. Mature pollen grains; ④b. Aborted ovary and ovules; ⑤a. Shrinking solid ovary. S. Stigma; Ovu. Ovules; Mc. Megaspore mother cell; Mp. Mononuclear pollen; Up. Uninucleate pollen; Vn. Vegetative nucleus; Rn. Reproductive nucleus; P. Pollen; Gp. Germ pore; Pp. Pistil primordiumFig.2 The external morphology and internal anatomical characteristics in the process of male flower formation at the late stage of sex differentiation of X. sorbifolium
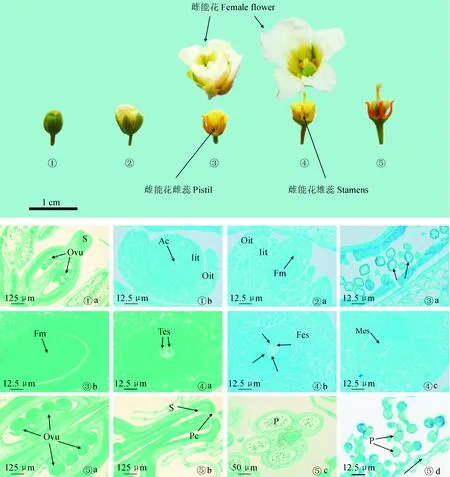
①.4月3日雌能花花蕾;②.4月9日雌能花;③.4月12日雌能花;④.4月15日雌能花;⑤.4月18日雌能花。①a.柱头伸长;①b.胚珠和孢原细胞;②a.小孢子母细胞四分体时期;③a.单核花粉粒有丝分裂时期;③b.单核胚囊;④a.双核胚囊;④b.四核胚囊;④c.八核胚囊;⑤a.胚珠;⑤b.花柱、柱头和乳突状细胞;⑤c.不开裂的花药;⑤d.花药壁残迹、败育花粉。Ovu.胚珠;S.柱头;Ac.孢原细胞;Iit.内珠被;Oit.外珠被;Fm.功能大孢子;Tes.二核胚囊;Fes.四核胚囊;Mes.成熟胚囊;Pc:乳突状细胞;P.花粉图3 文冠果性别分化后期雌能花形成过程中外部形态和内部解剖特点①. Female flower bud on April 3; ②. Female flower on April 9; ③. Female flower on April 12; ④. Female flower on April 15; ⑤. Female flower on April 18. ①a. Stigma extension stage; ①b. Ovules and sporogenous cells; ②a. The tetrad stage of microspore mother cell; ③a. The mitotic period of mononuclear pollen grains; ③b. Mononuclear embryo sac; ④a. Two-nucleus embryo sac; ④b. Four-nucleus embryo sac; ④c. Mature embryo sac; ⑤a. Ovules; ⑤b. Styles and mastoid cells on stigmas; ⑤c. Non-cracking anthers; ⑤d. Anther wall remnants and abortion pollen. Ovu. Ovules; S. Stigma; Ac. Archesporial cells; Iit. Inner integument; Oit. Outer integument; Fm. Functional megaspore; Tes. Two-nucleus embryo sac; Fes. Four-nucleus embryo sac; Mes. Matured embryo sac; Pc. Papillary cell; P. PollenFig.3 The external morphology and internal anatomical characteristics in the process of female flower formation at the late stage of sex differentiation of X. sorbifolium
2.3 文冠果花性别分化过程中气象因子的变化
文冠果花芽在3月3日开始萌动,根据1.3.4公式(1)得出文冠果生物学零度K为4.4 ℃。从2月下旬气温开始回升,至3月3日雌蕊原基开始分化,期间平均气温达到4.4 ℃,可视为休眠期结束的温度指标。花芽外发育各时期与该时期内气象因子的对应关系见表2。结合内部解剖结构观察结果,得出文冠果花芽外发育各时期开始时有效积温变化和各时期对应的平均温度变化趋势见图4。各时期对应的降水量、平均湿度变化趋势见图5。


表2 文冠果花性别分化时期与各时期内的气象因子
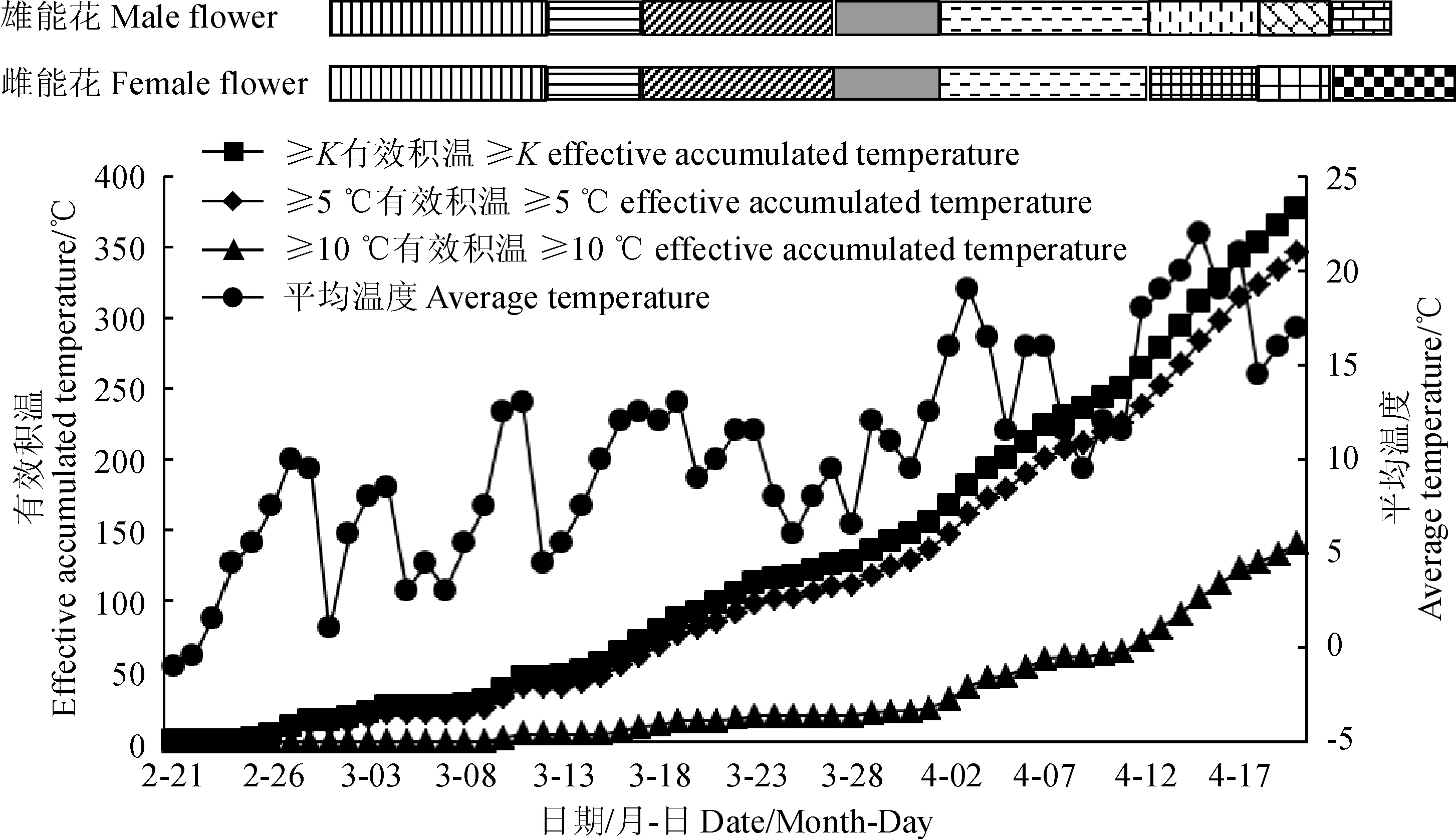
雄蕊原基分化期(2月21日~3月2日); 雌蕊原基分化期(3月3日~3月7日); 花药形成期、造孢细胞时期(3月3日~3月17日); 花药壁分化完全、小孢子母细胞时期(3月18日~3月22日); 小孢子母细胞减数分裂时期;子房胚珠雏形期(3月23日~4月2日); 雄能花小孢子母细胞四分体时期、单核花粉粒时期、子房胚珠败育期(4月3日~4月8日); 雌能花柱头伸长期、大孢子母细胞时期(4月3日~4月8日); 雄能花双核花粉粒形成期(4月9日~4月12日); 雌能花大孢子母细胞四分体时期、花粉败育期(4月9日~4月12日); 雄能花花粉粒成熟期、子房胚珠完全败育(4月13日~4月15日); 雌能花胚囊成熟期(4月13日~4月18日)图4 文冠果花性别分化过程中有效积温和平均温度的变化 Stamen primordium differentiation (February 21st to March 2nd); Pistil primordial differentiation (March 3rd to March 7th); Anther formation stage and spore cell stage (March 3rd to March 7th); Anther wall formation and microspore mother cells stage (March 18th to March 22nd); Meiosis of microspore mother cells; The early stage of ovary and ovules (March 23 to April 2); The tetrad stage of microspore mother cell, mononuclear pollen stage and the abortion stage of ovary and ovules in male flowers (April 3 to April 8); Stigma extension stage and megaspore mother cell stage in female flowers (April 3 to April 8); Dual nuclear pollen formation period in male flowers (April 9th to April 12th); The tetrad stage of megaspore mother cell and pollen abortion period in female flowers (April 9th to April 12th); Mature period of pollen grains and complete abortion of ovary and ovule in male flowers (April 13th to April 15th); Embryonic sac maturity of female flowers (April 13th to April 18th).Fig.4 Changes of effective accumulated temperature and average temperature in the process of flower sex differentiation in X. sorbifolium Bunge
由图4、图5和表2可见,花芽膨大前10 d,出现一次7.3 mm的降水。3月3日,≥K有效积温达到20.2 ℃时,雌蕊原基开始分化。3月8日,≥K有效积温达到25.5 ℃时,花药开始分化形成花粉囊,进入造孢细胞时期,雄蕊细胞分裂旺盛,直至3月22日形成小孢子母细胞,此时期≥K有效积温达到80 ℃,空气平均温度10.08 ℃。初春气温回升较慢,因而造孢细胞至小孢子母细胞形成时期持续时间较长,达15 d。期间降水为0,空气相对湿度处于较低水平39.53%。3月23日,≥K有效积温达到111.5 ℃是小孢子母细胞进入减数分裂时期的温度指标。小孢子母细胞减数分裂时期,空气平均温度10.05 ℃,此时期持续时间较长,达11 d。3月23日到4月2日,出现连续降水,累积达22.4 mm,空气相对湿度达到58.22%,适度的降水促进了内部子房胚珠的出现。4月2日,当≥K有效积温达到166.5 ℃时出现子房胚珠雏形,随即小孢子母细胞进入四分体时期,形成单核花粉粒。

图5 文冠果花性别分化过程中降水量和平均湿度的变化Fig.5 Changes of precipitation and average humidity in the process of flower sex differentiation in X. sorbifolium Bunge
接下来进入雄能花雌蕊和雌能花雄蕊败育时期。4月8日,当≥K有效积温达到230.6 ℃、≥5 ℃有效积温达到207.5 ℃、≥10 ℃有效积温为60 ℃时,雄能花胚珠内大孢子四分体出现异常,子房壁和胚珠细胞出现萎缩败育迹象;雌能花进入大孢子母细胞时期。此时空气平均温度11.5 ℃,平均相对湿度67.83%。4月12日,当≥K有效积温达到264 ℃、≥5 ℃有效积温达到238.5 ℃、≥10 ℃有效积温为71.5 ℃时,雄能花进入双核花粉粒形成期;雌能花大孢子母细胞进入四分体时期,花药中单核花粉粒有丝分裂发生异常,液泡化衰败。此时空气平均温度18 ℃,平均相对湿度38.75%,处于较低水平。4月3日到4月12日,是文冠果生殖器官发育的关键时期,此期间降水量仅为4 mm。4月13日到4月15日,出现较高降水量34 mm,适当的水分有利于雌雄配子体的成熟。至4月15日,≥K有效积温达到311.8 ℃,≥5 ℃有效积温达到284.5 ℃,≥10℃有效积温为102.5 ℃时,雄能花花粉粒成熟,但子房胚珠只剩细胞残迹。雄配子体成熟时期,空气平均温度达到20.33 ℃,平均相对湿度45.5%,持续时间仅3 d。4月18日,雌能花胚囊成熟,但花粉败育或花药不开裂。此时≥K有效积温达到353.1 ℃,≥5℃有效积温达到324 ℃,≥10 ℃有效积温为127 ℃。雌配子体成熟时期,空气平均温度19.25 ℃,平均相对湿度48.8%,持续时间6 d。
3 讨 论
明确花芽发育过程及与外部形态的对应关系,可为采取合理的栽培管理措施提供重要的参考依据。薄壳山核桃、板栗、山鸡椒等都有关于雌雄花芽发育进程外部形态的变化与内部解剖特点之间关系的报道[41-43]。本研究将文冠果性别分化各时期的外部形态特点、内部结构变化与气象因子对应起来,可以方便快捷地判断雌雄花芽分化时期。
典型的植物雌性不育主要表现为花柱与子房的异常发育[44]。文冠果雄能花雌性器官败育有两种情况,一种是雌蕊无花柱和柱头,仅由瘦瘪的子房组成,出现在4月2日左右,花蕾横径1.76~2.31 mm、纵径1.72~2.1 mm;另一种是雌蕊外表正常,但子房内部发育异常,出现在4月8日,雄能花花蕾横径2.05~4.54 mm、纵径2.99~5.32 mm,子房和胚珠开始出现败育迹象,此时≥K有效积温230.6 ℃。至子房完全败育,胚珠停止发育,柱头严重萎缩,此时≥K有效积温达到311.8 ℃,雌蕊败育期持续时间为8 d左右。
雄性不育是由于植株不能产生正常的花药和花粉[44]。本研究发现,文冠果雌能花雄蕊发育异常比雄能花雌蕊发育异常发生时间晚4 d。当花蕾横径12.25~18.30 mm、纵径8.30~10.98 mm,≥K有效积温达到264 ℃,空气平均温度18 ℃,平均湿度38.75%时,雌能花单核花粉粒有丝分裂异常,液泡化衰败。然后,花药停止生长,花粉囊不开裂,或花药绒粘层解体不完全,花粉败育。此时≥K有效积温353.1 ℃。雌能花花粉败育期持续时间为7 d左右。
目前,关于文冠果花的命名常有雌花、雄花、两性花、可孕花、不孕花[45-47]等多种说法。本研究发现,文冠果2种花都同时具有雌雄蕊,是后期雌雄蕊发育程度不同导致性别分化。基于此,认为从功能角度分别定义为“雌能花”和“雄能花”更准确。
前人研究认为文冠果性器官的败育可能与大小孢子营养不足有关[34-36]。雌能花败育的花药可能缺少某些细胞器,有生理生化代谢上的障碍,因此出现淀粉粒贮藏不足,子房和雄蕊对营养物质的竞争较大,小孢子因得不到足够的营养物质而败育[48]。胡青对2种类型花雌蕊发育情况比较研究发现,α-淀粉酶和过氧化氢酶的一些特殊酶带和差异蛋白与子房的萎缩和胚珠的不正常发育有关[36]。本研究推测温度有可能会影响文冠果雌雄性器官发育相关酶和蛋白的活性变化,但尚有待深入研究。此外,由于文冠果顶芽多为雌能花,侧芽多为雄能花,推测内源激素水平、营养状况对性别分化也有重要影响,这些因素的作用方式和机理都尚需要深入研究。
另外,本研究仅观测了雌能花雄蕊和雄能花雌蕊败育的发生过程和对应的气象条件,但要准确建立气象因素(温、湿度等)与性别分化过程的关系,还需要多年连续观测的数据来分析其相关性,建立回归方程,才能得出规律性结论。而且,可以在多个地点和林分进行连续观测,得出更完整准确的结论。
