内生菌协同宿主植物修复土壤复合污染的研究进展
2018-11-23吴春雅戴传超张凤敏
吴春雅, 戴传超, 张凤敏, 何 伟
(南京师范大学 生命科学学院,江苏 南京 210023)
随着工业化的发展,各种各样的污染物如重金属、有机污染物被排放到土壤中,造成严重的土壤复合污染。相比于单一污染物,复合污染物之间可能具有协同、相加或拮抗作用,提高了修复土壤复合污染的难度。复合污染指同一环境中存在两种或两种以上的污染物,可分为复合重金属污染、复合有机物污染及重金属-有机污染物复合污染[1]。土壤中的重金属溶解度低,不能降解为无害产物,其可通过食物、水、空气及皮肤接触进入人体[2]。有机污染物包括碳氢化合物、有机溶剂、挥发性有机化合物、卤化有机物和多环芳烃等,可抑制微生物的生长及代谢活动,影响植物生长发育,破坏土壤生态环境,且它们中的一些代谢物具有致癌性[3]。此外,有机化合物还可以进入食物链对生物造成毒性[4]。另外,植物本身也会产生一些有机物如酚酸、吲哚对植物和土壤造成损害[5]。目前已有多种方法用于治理土壤污染,主要分为物理修复、化学修复、生物修复。其中植物修复技术被认为是一种新型、有前景的生物修复方法,其使用绿色植物去除土壤中的污染物,目前已发现高浓度的污染物对大多数植物具有毒性,易损伤植物的代谢系统并影响植物生长状况[6],限制了该技术的应用。目前污染物、微生物和植物之间的相互作用引起研究人员的密切关注[7],当微生物与植物联用时,植物可为微生物提供栖息地及营养,而微生物也能通过增强植物的耐受性、促进植物生长及降解污染物改善植物生长环境。十几年来,研究人员运用各种微生物与植物联用修复土壤污染,近几年发现内生菌与植物关系更加密切,并且可能是植物的生长促进剂[8],因此提出使用内生菌与植物联用提高植物修复效率[9]。内生菌是指定殖于健康植物的各种组织和器官内部的细菌或真菌, 并且被感染的宿主植物不表现出外在病症[10]。最近已经发现内生菌主要通过促进宿主植物的生长,改变植物摄取污染物能力和酶降解污染物等方法增强宿主植物去除土壤中污染物的能力[11],因此内生菌与植物联用是一种非常有前景的植物修复方法[9]。本文总结和讨论了具有复合污染抗性内生菌的种类及内生菌促进植物生长和影响植物解毒/积累/移位的机制,展望了内生菌在植物修复中的应用前景和方向。
1 修复环境污染物的植物内生菌
有研究表明内生菌在植物中的定殖数量分布为根>茎>叶[12]。一般来说,大多数内生菌来自于根围、叶环或其他植物的附生细菌群落,可通过种子及受损的叶片组织传播[13]。近几年来,研究人员已从多种植物中分离出能够修复土壤重金属污染的内生菌,如超累积植物芥菜、东南景天、蜈蚣草、商路等,以及非超累积植物大豆、油菜、水稻、芦苇等。还有从小飞蓬、紫花苜蓿、黑麦草、看麦娘、互花米草等植物中分离出来的具有修复土壤有机污染物的内生菌。如表1所示,植物内生菌在修复复合污染的土壤中发挥重要作用。

表1 修复环境污染物的植物内生菌Table 1 Plant endophytic bacteria that repair environmental contaminants
1.1 修复土壤重金属复合污染的内生菌
重金属污染不仅会对与植物质量和产量产生不利影响,而且会改变植物相关微生物群落的大小、组成和活性。目前,研究较多的内生菌主要着生于植物根部,如Bacillussp.、Pseudomonassp.、Achromobactersp.等,其均具有去除土壤中复合重金属的能力,Babu等[14]从Alnusfirm中分离出的BacillusthuringiensisGDB-1能将土壤中77%的铅(100 mg/L)、64%的锌(50 mg/L)以及34%的砷(50 mg/L)转移到植物体内,以去除土壤中的重金属。内生菌不仅能在根中富集重金属,还能向茎、叶转移,从而提高植物对重金属的吸收效率。丁建等[20]从Spartinaalterniflora中分离的LysinibacillussphaericusQZ1-1可促进Spartinaalterniflora中铜、铅、铬的富集,并能将重金属向地上转移,为植物修复土壤复合重金属污染提供了良好的实验菌株。不同内生菌对不同金属的耐受性不同。如从Nypafruticans中分离的Pestalotiopsissp.对重金属的耐受性为锌>铅>铜>铬[21]。Sim等[22]从Phragmites中分离三种菌,发现与Phomopsissp.和Saccharicolabicolour相比,Trichodermaasperellum对各种金属具有更好的耐受性和适应性(高达2 000 mg/L),Trichodermaasperellum和Saccharicolabicolour对铜亲和力更高,其次是铬和铅,而Phomopsissp.对金属的亲和力为铜>铅>铬。Visioli等[23]发现主要定殖在根表面和叶表皮组织细胞间隙的ArthrobacterNcr-1和MicrobacteriumNcr-8共接种可促进植物生长,并可去除土壤中的镍、铁、钴和铜。
1.2 修复土壤有机复合污染物的内生菌
目前有研究表明Pseudomonassp.是降解多环芳烃(PAHs)的主要菌种,几乎可以降解所有四环以下的多环芳烃[24]。许多具有降解有机污染物的内生菌可以从受污染环境的植物根、茎部分离出来,与茎相比,根中降解有机污染物的内生菌数量更多。BurkholderiafungorumDBT1主要存在于杂交杨树(Populusdeltoides×Populusnigra)根组织中,并能耐受488 mg/kg的萘、菲、二苯并噻吩和芴,植物本身可以消除82%~87%的PAHs,而接种菌株DBT1后PAHs的降解率可高达99%[17]。污染物的添加可能会改变植物内生菌群落,菲污染增加了盐沼植物Spartinaalterniflora中Proteobacteria的丰度,但减少Firmicutes、Bacteroidetes及Chloroflexi的数量。通过芘处理增强Spartinaalterniflora中Proteobacteria和Firmicutes的丰度,说明PAHs污染可以改变Spartinaalterniflora根部内生细菌群落,盐沼植物中的内生细菌可能在植物修复土壤PAHs污染中起重要作用[25]。不同内生菌对有机污染物的降解率不同,并且添加一定的营养物质可能增强污染物的降解。Zhu等[18]发现Conyzacanadensis和TrifoliumpretenseL.中的Stenotrophomonassp. P1可降解98.0%萘、83.1%芴、87.8%菲、14.4%芘和1.6%苯并(a)芘,而Pseudomonassp. P3可降解95.3%萘、87.9%芴、90.4%菲、6.9%芘,与菌株P1相比,菌株P3从植物组织中去除PAHs的效率更高,并且两种菌均可以PAHs为碳源生长,另外额外加入有机碳和有机氮营养可显著增强PAHs的生物降解。植物物种的选择也会影响接种后内生菌的定殖率和代谢活动,并最终影响植物修复过程,Fatima等[26]的调查显示Brachiariamutica中内生菌定殖率和代谢活动比Leptochloafusca更高,并且Acinetobactersp. BRSI56和PseudomonasaeruginosaBRRI54共接种对原油的降解率较高,在Brachiariamutica中的降解率最高为78%。目前的研究主要集中于内生细菌协同植物降解复合污染物,而对于内生真菌,有研究表明内生真菌Phomopsissp. B3与水稻联合处理30 d后,土壤中菲的残余量仅为5.932 mg/kg,为初始菲含量的3.95%;单独使用菌株B3修复,土壤中残余的菲为11.186 mg/kg[27],这些研究表明植物也能与内生真菌联合作用提高植物修复土壤污染的效率,为后人研究内生真菌协同宿主植物修复土壤复合有机污染的研究做了前期准备。
1.3 修复土壤重金属-有机复合污染物的内生菌
目前对内生菌协同植物修复土壤重金属-有机复合污染的研究较少,对于复合污染Weyens等主要使用基因工程菌株Burkholderiasp.开展了一系列研究。发现在Lupinusluteus中接种基因工程内生菌BurkholderiacepaciaVM1468可减少污染物对植物毒性,接种后植物根生物量增加了30%。与未接种的对照相比,接种植物中三氯乙烯蒸散量呈下降趋势,根中的镍浓度高出约7倍,茎中增加了5倍[28]。接种内生菌BurkholderiacepaciaVM1468后,在镍和甲苯复合污染土壤中植物对镍吸收增加,但减少了甲苯的蒸散量,同时接种后植物对甲苯降解能力不受镍的影响[29]。接种BurkholderiavietnamiensisBU61的柳树修复镉和甲苯复合污染土壤,发现接种后提高甲苯的降解能力,降低了其毒性和蒸散量,但镉吸收和易位没有受到影响[30]。这些研究证实,基因工程菌株可以用于提高植物重金属耐受性并降解有机污染物,为修复复合污染土壤提供良好的菌株。
2 植物内生菌协同修复土壤重金属复合污染的作用机制
已有充分证据证明内生菌可以帮助宿主植物适应恶劣条件并通过促进植物生长,降低重金属对植物毒性、改变土壤中重金属生物有效性和植物中金属转运能力及表达重金属解毒基因或蛋白提高植物修复效率。总体上,内生菌通过两种手段促进复合重金属污染土壤中植物修复过程,即促进植物生长及改变植物摄取重金属的能力。表2列举了内生菌对复合重金属的吸收机制。
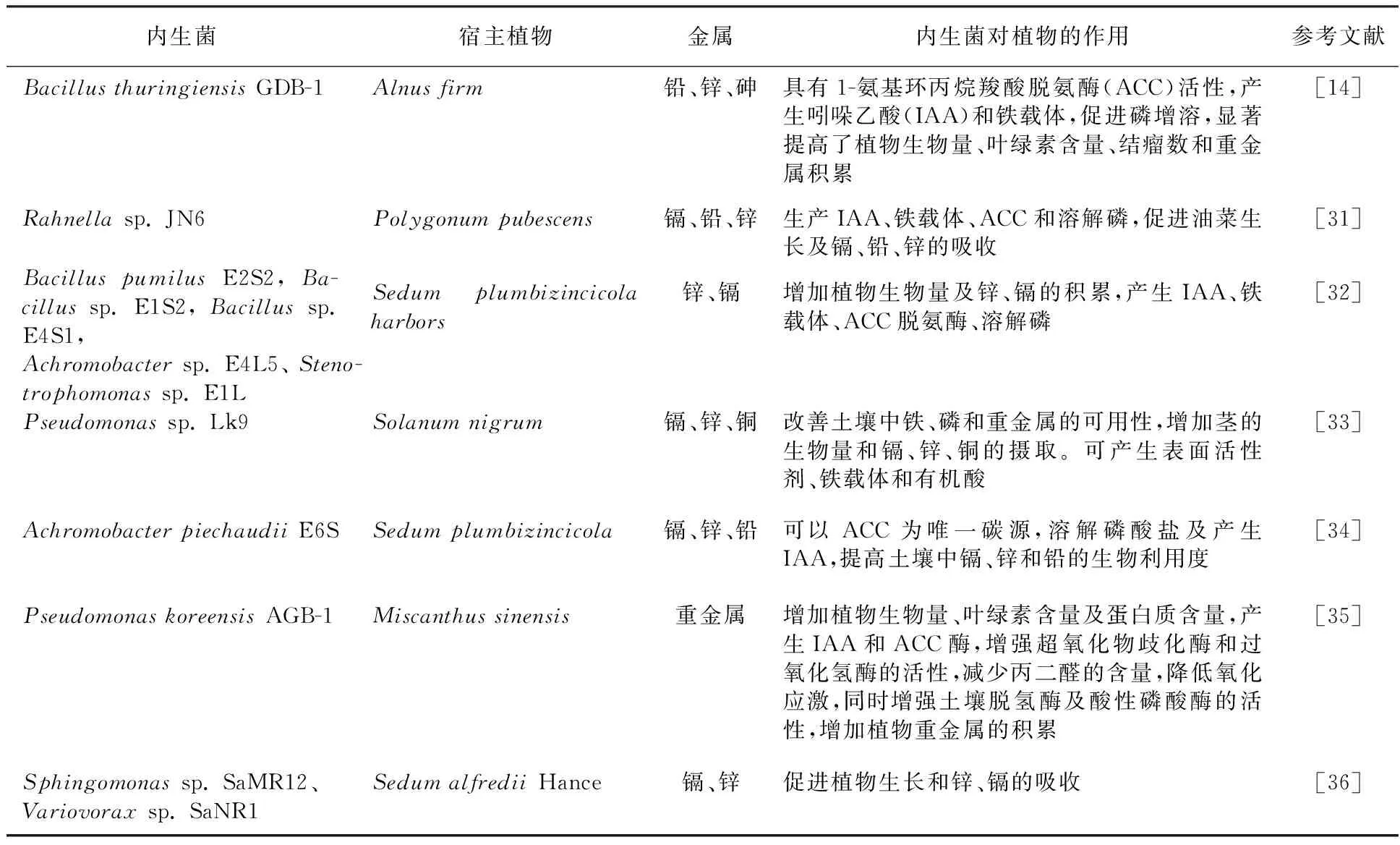
表2 内生菌对复合重金属的吸收机制Table 2 Mechanisms of endophytic bacteria absorb multiple heavy metals
2.1 促生机制
促生长菌主要通过溶磷,产生铁载体和植物激素缓解重金属毒性并促进植物生长。在重金属胁迫下,发现一些耐重金属胁迫的内生菌可通过酸化、螯合和有机酸的释放溶解沉淀在土壤中磷酸盐或通过分泌细胞外酸性磷酸酶使土壤中的有机磷矿化,从而提高土壤中磷的利用率,研究发现磷酸盐的增溶与土壤中重金属摄取有关,可增加植物重金属的积累。接种Rahnellasp. JN6可观察到Brassicanapus中可溶性磷酸盐显著增加,并且与未接种的相比,植物地上和根组织中镉、铅和锌的浓度和植物干重也显著增加[31]。植物一般通过产生铁载体溶解不溶性铁,以及一些植物所需的微量元素,如铁、锰、锌、铜、钼、钠、镍等,以促进植物生长。内生菌也能够向植物供应必需的生长调节剂以增强营养物积累和代谢,生长调节剂相互作用可促进植物生长发育。IAA(indole-3-acetic acid,吲哚-3-乙酸),作为主要生长素,参与植物的各种生理过程。内生菌可调节植物生长调节剂增加自身生物量,分散植物某些部位积累的重金属,以降低对植物的毒性。大量研究表明内生菌可产生IAA促进植物生长,BacilluspumilusE2S2产生的IAA量最高,并显著增加了Sedumplumbizincicolaharbors的根长(146%)、茎长(17%)、鲜重(37%)和干重(32%),表明IAA可通过调节细胞分裂和分化促进植物生长和发育[32]。植物在重金属胁迫下,可产生大量乙烯,抑制植物生长,导致植物死亡,而某些内生细菌可产生ACC脱氨酶,同时能以ACC为碳源,降低植物中乙烯的含量。Babu等[14]在Alnusfirm中接种BacillusthuringiensisGDB-1,其具有ACC脱氨酶的活性,可增加植物生物量,进而增加植物中重金属的积累。
2.2 改变植物摄取重金属的能力
重金属从土壤转移到植物中主要取决于土壤中重金属生物有效性,而重金属的生物有效性也受土壤颗粒大小、养分、pH、氧化还原电位、有机物和其他离子等因素的影响。研究表明,具有重金属抗性的内生菌可以减轻污染物的毒性,其主要通过释放金属螯合剂(例如铁载体、有机酸、生物表面活性剂)、酸化土壤、改变氧化还原活性来增加重金属的生物有效性。
铁载体主要存在于根际,可提高Fe3+及其他各种二价金属离子的移动性及可溶性重金属的浓度,所以Saha等[37]认为铁载体在内生菌协同植物修复土壤重金属复合污染中发挥关键作用。Yamaji等[38]从Eleutherococcussciadophylloides中分离出463株菌株,107株能产生铁载体,并累积大量的铯和锰,也能产生螯合铁和铝的铁载体,导致铯污染土壤中铯和锰的解吸。其中4株菌铁载体产量较高,分别为Pseudomonascitronellolisnllol、Stenotrophomonasmaltophiliat142、Pseudomonasdenitrificansn142、Pseudomoansdenitrificansn164,均可增强土壤中铁、铝、铯和锰的解吸能力,降低土壤pH。
一般来说,根部渗出的低分子量有机酸在植物根际发挥重要作用,例如获取营养,调节植物生长和缓解某些元素的毒性,还可通过络合反应结合土壤溶液中的金属离子,提高重金属的溶解度,降低土壤pH。Chen等[33]发现Pseudomonassp. Lk9和Solanumnigrum相互作用可促进植物根分泌有机酸,提高土壤中可溶性重金属的浓度,增加植物镉、锌、铜的摄取能力。Ma等[34]发现Sedumplumbizincicola接种AchromobacterpiechaudiiE6S后可产生有机酸和溶解磷酸盐,引起土壤pH降低,并增加土壤中重金属的溶解度,提高了根际土壤中重金属的生物利用度,同时增加植物生物量,提高植物摄取和积累镉、锌、铅的能力,而接种菌株E6S也大大减少了根中镉和锌的转移,细菌细胞以一定顺序结合金属离子(锌>镉>铅),表明细菌接种可以协助宿主植物根吸收和储存重金属,减少重金属向茎叶转移,以降低对植物的毒性。内生菌接种可改变土壤微生物群落结构和根部分泌物,同时促进植物吸收重金属。陈宝[39]发现接种植物内生菌Sasm03能够显著增加根际土壤的细菌群落结构多样性和宿主植物的根系分泌物,促进植物对锌、镉的吸收。
此外,由内生菌产生的生物表面活性剂可用来增加难溶性金属的生物利用度,并加速植物修复速率[40],主要通过降低土壤-水界面张力和流体力来促进重金属从土壤基质到土壤溶液的解吸[41]。因此,产生生物表面活性剂的内生菌可有效地增强重金属的流动性,从而增加植物中重金属的积累。
内生菌可以改变重金属的生物利用度并在植物中易位,其可分泌多种代谢物,包括铁载体、有机酸(如柠檬酸、草酸、乙酸等)改变重金属的生物有效性并增加植物对重金属的摄取能力。如BacillusthuringiensisGDB-1可通过产生铁载体螯合重金属,有机酸降低土壤pH,提高重金属的生物利用度,从而增加Alnusfirm根和茎中镍、镉、锌、铅、铜、砷的浓度[14]。
2.3 重金属解毒基因及相关蛋白的表达
重金属的毒性是影响植物修复的关键因素,为了克服金属的毒性,一些内生菌通过编码重金属解毒基因减轻重金属胁迫,如亚砷酸盐转运蛋白(arsB)基因可催化砷(V)还原成砷(III),亚砷酸盐氧化酶(aoxB)可将砷(III)氧化成砷(V)[42]。金属转运蛋白可介导多种金属离子与亚细胞螯合,有助于金属离子从细胞质运送到细胞外或细胞器内。有研究从重金属耐受的深色有隔内生真菌(DSE,Exophialapisciphila)中克隆出保守的金属转运蛋白基因(EpNramp),该基因表达的蛋白EpNramp可转运铁和镉,EpNramp基因的下调有助于减少细胞质中镉的吸收,可能是通过一系列配体(谷胱甘肽(GSH)、金属硫蛋白)将金属螯合在细胞质内或从胞质溶胶流入隔离室[43]。将谷胱甘肽合酶基因gcsgs导入Enterobactersp. CBSB1,接种基因工程菌株CBSB1-GCSGS的Brassicajuncea对镉和铅摄取量最高[44]。在4种重金属胁迫下,表达谷胱甘肽过氧化物酶基因EpGSTT1、EpGSTT2和EpGSTT3活性显著上调,有利于降低ROS损伤,并有助于Exophialapisciphila的重金属耐受[45]。在镉胁迫下,参与金属摄取(ZIP)、易位(MTP)和螯合(PCS)的3个玉米基因受DSE定殖和镉胁迫的影响,接种DSE显著下调除了低镉处理的其他所有处理的根和叶的ZIP基因,表明接种DSE后玉米根中镉吸收减少,MTP转运蛋白能够通过增加液泡内金属的螯合来介导突变株对锌和镉的超敏反应。因此上调的MTP是通过特异性细胞区室的隔离(例如液泡)帮助镉的解毒。此外,植物螯合肽合酶基因(PCS)的上调,具有改善谷胱甘肽和相关硫醇肽的催化转肽作用,这可能在镉运输和解毒中起重要作用[46]。
3 内生菌协同宿主植物修复土壤有机复合污染物的作用机制
在植物修复有机复合污染物过程中,内生菌通过促进植物生长,降低有机污染物在植物组织中的比例,改善植物的适应能力,从而增强植物修复效率。此外,内生菌产生不同的酶矿化有机污染物,并降低植物中挥发性有机污染物的毒性[47]。还有些内生菌同时具有促生长和酶降解功能,在修复土壤有机复合污染过程中发挥重要作用。表3列举了内生菌对复合有机污染物的吸收机制。

表3 内生菌对复合有机污染物的吸收机制Table 3 Mechanisms of endophytic bacteria absorb multiple organic pollutants
3.1 促生长机制
植物内生菌可以多环芳烃为碳源,减少土壤及植物中PAHs的含量及毒性,促进植物生长。Peng等[48]从AlopecurusaequalisSobol和OxaliscorniculataL.中分离出68株内生细菌,分为3个门:Firmicutes、Proteobacteria和Bacteroidetes。其中Bacillussp.和Pseudomonassp.为主要种群。它们中的一些能够以PAHs作为惟一碳源和能量迅速生长,表明这些菌株可能具有降解植物中PAHs的潜力。Ho等[52]研究表明AchromobacterxylosoxidansF3B接种至Vetivergrass中可以芳香族化合物(苯、甲苯、乙苯、二甲苯、邻苯二酚、苯酚)为碳源且能够提高宿主植物的生物量。内生菌能产生生长素、ACC脱氨酶刺激植物生长,还可以刺激植物抗性机制,通过竞争营养物和空间,减少病原体的生长和活性,间接改善污染土壤中植物的生长和有机污染物的降解。Bisht等[19]将Bacillussp. SBER3接种至Populusdeltoides,4 d后产生22 mg/mL的IAA,并在120 h后溶解0.96 mg/mL的有效磷。此外,Bacillussp. SBER3还产生铁载体和ACC脱氨酶。除了这些性状,菌株SBER3还能在体外条件下抑制植物病原真菌Rhizoctoniasolani、Macolaminaphaseolina、Fusariumoxysporum和Fusariumsolani的生长。菌株SBER3在液体培养基中,培养6 d后,蒽和萘的浓度分别降低了83.4%和75.1%,在土壤模型系统中共降解了45.6%的PAHs。内生菌接种对植物根系分泌物及微生物群落产生影响。接种Pseudomonassp.改变Scirpustriquete根系土壤的微生物群落,其微生物数量增加但种类降低。接种Pseudomonassp. J4AJ 可以促进Scirpustriquete的生长,从而影响其根际作用,而Scirpustriquete的根际活动又可以反过来为Pseudomonassp.提供各种营养物质,土壤中的耐油菌可以更好地存活和增殖,两者协同作用促进土壤中柴油的降解[53]。
3.2 共代谢机制
可通过添加额外的营养物或微生物能直接利用的碳氮来刺激内生菌[54],同时增强内生菌降解低分子量有机污染物的能力,高分子量的有机物溶解度较低,难以降解,可通过共代谢提高内生菌对污染物的降解能力。共代谢定义为微生物不能以某些难降解的有机物作为碳源和能源,需要有额外的化合物提供碳源或能源才能降解该有机物[55]。Zhu等[56]发现从PlantagoasiaticaL.中分离出的Paenibacillussp. PHE-3在添加额外的谷氨酸培养84 h后,菲的最大降解效率可达99.9%。此外,菌株PHE-3对菲的降解与儿茶酚2,3-双加氧酶活性呈正相关,表明菌株PHE-3具有降解高分子量PAHs(HMW-PAHs)的能力。低分子量有机化合物可以通过共代谢增强HMW-PAHs的生物降解[57],如菌株PHE-3可以使用菲作为底物共同降解苯并(a)芘,在其他2-3环PAHs的存在下,菌株PHE-3可通过共代谢有效地降解HMW-PAHs。
3.3 酶降解机制
内生菌可通过氧化或还原酶系统将有毒化合物分解为毒性较小的化合物保护其宿主免受污染物的损伤,如漆酶、脂肪酶、细胞外酶[54]和其他氧化反应酶(含细胞色素P450的单加氧酶、过氧化物酶、过氧化氢酶等),儿茶酚2,3-双加氧酶是双加氧酶的超家族的成员,其可催化儿茶酚并取代儿茶酚环,这也是细菌降解多环芳烃的重要机制[56]。有研究发现漆酶在降解酚酸类有机污染物如芥子酸[58]、阿魏酸[59]等方面发挥重要作用,其催化各种芳族化合物的氧化,当内生真菌Phomopsisliquidambari产生的漆酶施用于土壤时,花生的生物量增加了12%,同时土壤中香草酸,香豆酸和4-羟基苯甲酸的含量分别下降了21%、27%和40%[49],说明内生菌产生的漆酶可降解土壤中毒性酚酸化合物,改善土壤环境,可在农业应用上发挥重要作用。内生真菌Phomopsisliquidambari可通过脱羧、氧化、羟基化和开环反应降解有机物肉桂酸,这种降解过程与酚酸脱羧酶、漆酶、羟化酶和原儿茶酸3,4-双加氧酶的作用密切相关,酶的活性和基因转录动力学也与相应代谢物浓度的变化一致[60]。同时Phomopsisliquidambari产生的双加氧酶可降解植物雌激素木犀草素[61],酶代谢途径中产生的某些物质也可能为内生菌提供碳源或能源,以加速有机污染物的降解。Kukla等[62]指出从LoliumperenneL.中分离出的大多数内生菌都具有碳氢化合物降解酶,表明这些细菌可以降低植物吸收烃后产生的毒性。Hong等[63]发现菲和芘处理导致非根际沉积物中的脱氢酶活性增加,促进沉积物中PAHs的生物降解。有研究报道,锰过氧化物酶(MnP)在PAHs降解中起关键作用[64],Dai等[3]的研究表明CeratobasidumstevensiiB6产生的MnP受菲诱导,但MnP的活性与菲降解没有明显的相关性,可能是由于真菌代谢菲的关键酶位于真菌菌丝体中,新鲜培养基促进真菌菌丝体中酶的生成,一种可能是MnP参与了菲代谢物的降解,另一种可能性是MnP通过使用像过氧化氢这样的共底物氧化菲。有研究表明,alkB基因是一种编码单氧化酶或烷烃羟化酶的基因,其丰度与基因表达、碳氢化合物去除成正比,这表明分解代谢基因的活性与烃降解直接相关[26]。BurkholderiaphytofirmansPsJN携带编码各种降解酶的基因,如烷烃单加氧酶(alkB)和细胞色素P450烷烃羟化酶的基因,其能降解土壤中柴油污染[50]。其他烃类降解基因有ndoB和phnAc,分别负责萘双加氧酶和菲双加氧酶的表达,可降解萘和菲。EnterobacterludwigiiBRI10-9对柴油降解率最高为68%,并且菌株BRI10-9与Italianryegrass联合作用效果较好,内生菌BRI10-9具有表达细胞色素P450羟化酶的基因CYP153,该基因具有降解碳氢化合物的能力,其表达和丰度在根际最高,这可能是由于根系渗出大量营养物质可用于烷烃降解的细菌生长和共代谢[9]。
4 内生菌协同宿主植物修复土壤重金属-有机复合污染物的作用机制
由于重金属的存在可能会抑制内生菌对各种有机污染物的生物降解,所以目前对于内生菌协同宿主植物修复土壤重金属-有机复合污染物的研究较少,其主要通过促进植物生长,降低氧化应激水平减轻复合污染物对植物的毒性。
Gutiérrez-Ginés等[65]从LupinusluteusL.根中分离出一些内生菌,其中1株具有较高的促生长能力,还有9株菌株可以有机污染物为唯一碳源生长,有趣的是试验发现与人工施加苯并芘土壤中生长并接种内生菌的植物相比,在天然垃圾填埋土壤生长的植物生长能力下降,表明植物内生菌可能不适应植物体内重金属的浓度。Weyens等[66]接种基因工程菌株PseudomonasputidaW619-TCE后,杨树根重显著增加。此外,接种后内生菌可减少根部的氧化应激水平减轻植物毒性。考虑到植物修复效率,接种菌株W619-TCE导致根部氮吸收增加45%。
5 展 望
土壤环境中存在多种污染物,虽然植物内生菌协同宿主修复土壤污染已经被广泛用于重金属复合污染物及有机污染物复合污染的研究,并在实验室条件下取得一定成效,然而关于重金属-有机物复合污染的研究相对较少。为加速处理土壤中的复合污染物,需要阐明内生菌在植物修复中的作用机制。此外,需要进一步探索以下几个发展方向:①需要进一步加强对土壤中复合污染的研究,增加复合污染物的数目和类别对修复土壤复合污染具有非常重要的实践意义;②可研究复合内生菌修复土壤复合污染的效果及机制。未来需要将具有不同优势内生菌结合起来,发挥各自的长处,共同作用,研发菌剂并大规模地应用于土壤复合污染修复;③需要探索内生菌降解复合污染物的有利性状,以获得在该领域中植物和内生菌的联合使用的最大效果;④可采用基因工程的方法改良内生菌与植物的相互作用,寻找修复土壤污染的新策略;⑤分析与污染物降解相关的基因、蛋白质及生物体内的一些代谢物,探索植物与内生菌相互作用的分子机制;⑥研究内生菌如何影响土壤微生物群落结构及微生物群落修复土壤复合污染的作用机制,将内生菌辅助的植物修复更加有效、大规模地应用于处理土壤复合污染。
