氮沉降和磷添加对杉木光合及叶绿素荧光特征的影响
2018-11-07钱树玥王巧刘宁李全雷赵枫宋新章哀建国
钱树玥, 王巧, 刘宁, 李全, 雷赵枫, 宋新章, 哀建国
氮沉降和磷添加对杉木光合及叶绿素荧光特征的影响
钱树玥, 王巧, 刘宁, 李全, 雷赵枫, 宋新章, 哀建国*
亚热带森林培育国家重点实验室, 浙江农林大学, 临安 311300
为探讨杉木()光合及叶绿素荧光参数在大气氮沉降和磷添加情况下的变化, 实验以10龄杉木为研究对象, 共设9个处理水平: 低氮(N30: 30 kg·ha-1·a-1), 高氮(N60: 60 kg·ha-1·a-1), 低磷(P20: 20 mg·kg-1), 高磷(P40: 40 mg·kg-1), 低氮低磷(N30 + P20), 低氮高磷(N30 + P40), 高氮低磷(N60 + P20), 高氮高磷(N60 + P40)和对照处理组(CK)。结果表明: 在夏季, 氮磷添加对杉木的最大净光合速率(Pn max)无显著影响。单独添加氮、磷都抑制了杉木的最大荧光产量(Fm)、初始荧光产量(F0)、PSII潜在活性(Fv/F0)值; 单独添加磷促进了杉木的叶色值(SPAD); 在磷添加情况下, 低氮增加了杉木的Fm, 高氮增加了杉木的SPAD值, 降低了杉木的非光化学淬灭系数(qN)值。在秋季, 单独添加氮促进了杉木的最大净光合速率。单独添加氮、磷抑制了杉木的SPAD值。在磷添加情况下, 氮沉降增加了杉木的SPAD值, 降低了杉木的F0值。夏季杉木叶片N含量与SPAD呈显著正相关(<0.01), 秋季杉木叶片N含量与SPAD和光化学淬灭系数(qP)呈显著负相关(<0.05), 而与Fm和F0呈显著正相关(<0.05)。
氮沉降; 磷胁迫; 杉木; 最大净光合速率; 叶绿素荧光
1 前言
磷是植物生长发育所需的营养元素之一, 在植物生长过程中起着极为重要的作用[1]。虽然南方林区以酸性红壤为主, 全磷含量丰富, 但能被植物吸收利用地有效磷极易被Fe3+、Al3+和Ca2+等离子固定[2], 导致土壤为植物提供的有效磷含量极低[3], 因此, 植物长期处于低磷胁迫的状态。已有研究表明: 在低磷胁迫下植物的生长与繁殖在形态、生理、生化等方面都会发生显著变化[4, 5], 在低磷胁迫下菜豆和向日葵的光合作用明显下降[6]; 低磷胁迫下玉米叶片的最大净光合速率与最大PSII的光能转换效率等荧光参数值受到抑制, 植株生物量下降[7]; 在低磷胁迫下, 水稻叶片的光合速率和叶绿素荧光参数下降[8]; 缺磷导致植物最大净光合速率、最大荧光产量、电子传递速率、最大PSII的光能转换效率等降低, 进而抑制了叶片光合作用[9]。可见磷对植物的生长发育起到极为重要的作用。郭淑青等人发现在15 mg·kg-1的磷浓度水平可提高金露梅叶片的光合能力[10]; 在小麦[11]、玉米[7]等作物上的研究表明, 磷添加能保持植物良好的光合性能。为了应对磷胁迫引起的一系列问题, 许多学者研究了磷添加对植物光合等生理特征的影响, 分析植物光合生理生态特征对磷添加响应的研究提供重要的理论依据。
氮沉降是酸沉降的一种, 主要是指大气中含氮化合物经过物理化学变化形成硝酸重新进入生态系统的过程[12]。近年来, 大气氮沉降的增加对生态系统的影响, 及其所带来的生态效应逐渐成为人们关注和研究的热点[12]。已有研究表明植物叶片中的氮含量、叶绿素含量与荧光动力学参数值均与氮沉降相关[13–15]。氮沉降抑制了植物的生长速度, 影响着植物的生长状况[16]。据1999—2003年观测数据显示, 我国已成为世界第三大高降区[17]。在未来的几十年中, 我国氮沉降现象将日益严重[18]。目前我国亚热带地区的氮沉降量达到65.53 kg·ha-1·a-1, 是我国氮沉降最严重的区域[18, 19]。因此, 我国亚热带地区的森林生态系统极易受到氮沉降的影响[19]。一些学者研究发现氮沉降促进了亚热带木荷()、锥栗()、黄果厚壳桂()的最大净光合速率和气孔导度, 而且在100 kg·ha-1·a-1的氮沉降水平下最大净光合速率达到最大, 但超过该水平氮沉降则表现为抑制作用[20]。目前, 氮沉降对植物生理生态学影响的研究主要集中在生长发育、形态变化、养分利用效率等方面[21]。因此, 研究氮沉降对植物光合及叶绿素荧光的生理生态影响具有重要的意义。
杉木()是中国南方重要的造林和用材树种, 种植广泛, 面积1239.1×104hm2, 蓄积量为47357.33×104 m3, 分别占全国人工林面积和蓄积量的26.55%和46.89%, 在中国人工林中占据重要地位[22]。杉木林林地土壤一般呈酸性或强酸性反应, 土壤磷素含量很低, 且有效磷水平极低, 甚至难以检测, 导致土壤供磷不足[2]。目前, 杉木人工林对氮沉降的响应已有一些研究, 如氮沉降对生产力、凋落物分解、土壤养分、养分平衡[23]等的影响, 但氮沉降和磷添加对杉木生理特征的影响目前还没有报道。因此, 本实验以杉木林为研究对象, 研究氮沉降和磷添加对杉木光合及叶绿素荧光等生理特征方面的影响, 以期为认识氮沉降和磷添加对杉木生理特征的影响提供科学依据和参考。
2 材料与方法
2.1 试验地概况
试验地位于浙江省杭州市临安区高坎村(119°67′E, 30°21′N), 属于中亚热带季风气候, 四季分明, 气候温和, 雨量充沛, 年均降水量1420 mm左右, 年均气温15.6 ℃, 年均无霜期230 d 左右。土壤为黄土壤, 地形地貌为低山丘陵。
2.2 试验设计
本实验时间为2017年1月份至2017年10月份。实验选取试验样地内27棵长势相近且平均高度为3m的10龄杉木, 以每棵杉木中心设置一个3 m × 3 m独立样方, 每个样方之间至少相隔6m。模拟氮沉降处理参照国际上氮沉降模拟研究的试验方法, 依据我国亚热带地区的实际氮沉降量及未来增加趋势及磷胁迫处理参考国内外相关研究[17, 19], 设置9个处理, 每个处理3个重复: 低氮(N30: 30 kg·ha-1·a-1), 高氮(N60: 60 kg·ha-1·a-1), 低磷(P20: 20 mg·kg-1), 高磷(P40: 40 mg·kg-1); 低氮低磷(N30 + P20), 低氮高磷(N30 + P40), 高氮低磷(N60 + P20), 高氮高磷(N60 + P40)以及对照处理组(CK)。
土壤基质的速效P水平通过将KH2PO4均匀地洒在样方内, 进行30 cm的翻耕, 使其土壤上层速效磷含量达到P20和P40两个供磷水平, 此后试验处理中不再对杉木进行磷添加。同时, 根据氮处理梯度水平, 每个月进行模拟氮沉降喷施一次, 每年12次。具体方法为: 每月月初将每个样方所需喷施的一定量的NH4NO3溶解在4升自来水中, 用背式喷雾器从杉木的树冠上方人工来回进行均匀喷洒。对照处理喷洒同量的自来水但不添加氮, 以减少处理间因外加自来水不同而造成的影响。
2.3 光合生理和叶绿素荧光的测定
在2017年7月与10月天气晴好的09: 00—12: 00 am, 取每个处理组中3株待测杉木上3片中部向阳的邻近健康叶片, 并采用Li-6400便携式光合测定仪(美国LI-COR公司)测定杉木叶片的最大净光合速率(Pn max)。采用SPAD-502叶绿素计(日本美能达公司)测量与用于光合测定相同的杉木叶片的叶色值(SPAD), 每株杉木的叶片测定5次, 每个处理组重复3次; 采用PAM-2500便携式叶绿素荧光分析仪(德国Walz公司)在同一时间测量相同杉木上相同叶片的各荧光参数[24], 其中包括: 最大荧光产量(Fm)、初始荧光产量(F0)、最大PSII的光能转换效率(Fv/Fm)、PSII潜在活性(Fv/F0)、光化学淬灭系数(qP)、非光化学淬灭系数(qN)、实际量子产量(YII)。
2.4 数据分析
采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)对不同模拟氮沉降强度和磷添加下杉木的最大净光合速率和叶绿素荧光参数数值进行比较分析, 利用皮尔森相关性分析了叶片中的N、P含量和杉木的各生理特征的相关性。数据分析利用SPSS18.0软件, 采用SigmaPlot12.5软件制图。
3 结果与分析
3.1 光合特征
如图1(a)所示, 在夏季, 单独添加氮磷后杉木的最大净光合速率均无显著差异(>0.05)。如图1(b)所示, 在秋季, 单独添加氮显著提高了杉木的最大净光合速率(<0.05); 高磷处理下, 添加氮显著增加了杉木的最大净光合速率(<0.05)。其他处理并无显著差异。如图1(a)与图1(b)所示, 杉木在秋季的最大净光合速率均高于夏季; 两季均在高氮处理下杉木的最大净光合速率达到最高值, 且秋季是夏季的1.21倍。
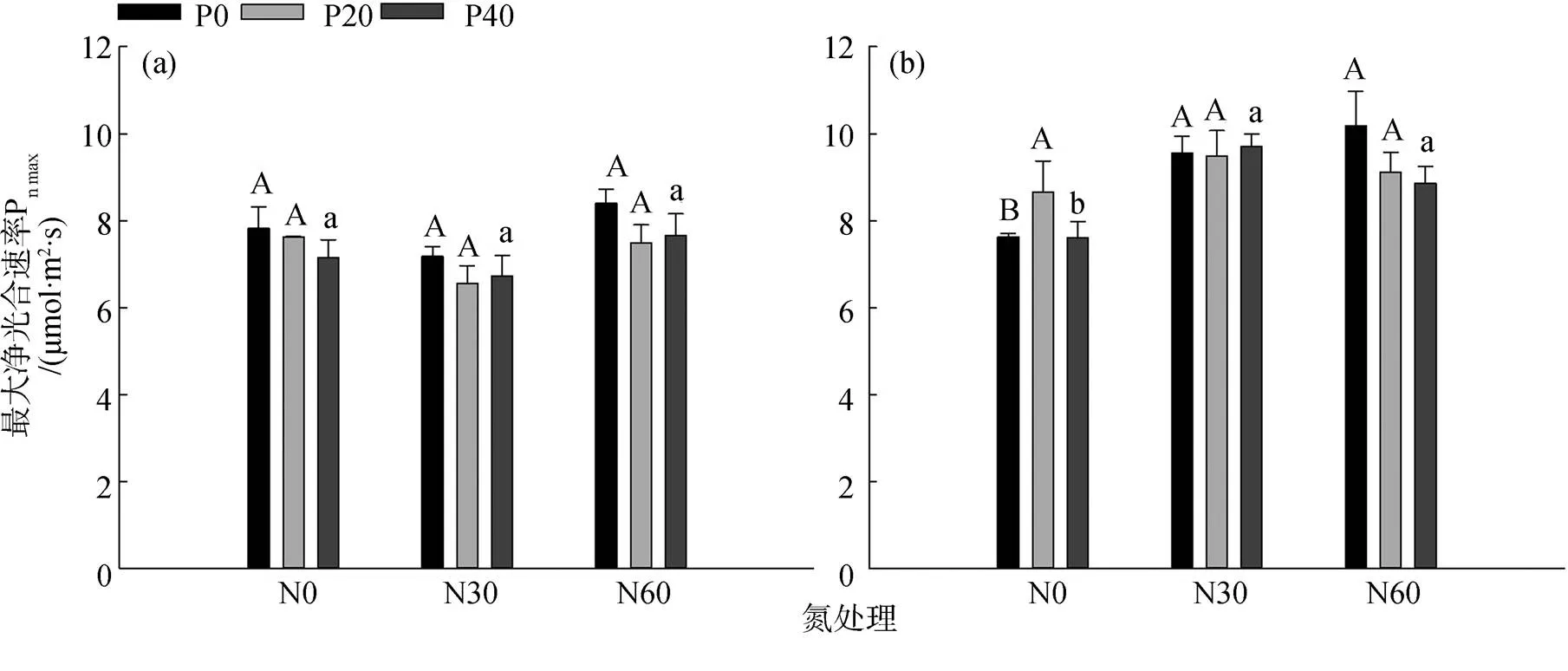
注: 不同大小写与加粗大写字母分别表示在无磷、低磷和高磷处理下, 不同氮处理间杉木最大净光合速率的差异性(p<0.05)。*表示在无氮增加处理下不同磷处理间的差异性, *表示p<0.05; **表示p<0.01; ***表示p<0.001,下同。3
3.2 叶色值
如图2(a)所示, 在夏季, 杉木的SPAD值在高磷情况下显著上升(<0.01); 高氮与磷处理(低磷和高磷)复合下, 杉木的SPAD值显著上升(<0.05)。如图2(b)所示, 在秋季, 单独添加磷后杉木的SPAD值显著下降, 且在低磷情况下抑制性更强(<0.001); 与对照组相比, 高氮显著下降了SPAD值; 磷添加处理下, 氮沉降的添加显著上升了杉木的SPAD值。夏季杉木的SPAD值高于秋季。
3.3 荧光参数
如图3(a、c、e、g、i、k、m)所示, 在夏季, 单独添加氮、磷都显著抑制了杉木的Fm、F0、Fv/F0值(<0.001); 单独添加磷后杉木的qN值在低磷处理下显著增加, 而在高磷处理下显著降低(<0.05); 磷添加处理下, 低氮添加显著增加了杉木的Fm值, 高氮添加显著降低了杉木的qN值(<0.05); 低磷处理下, 低氮添加显著增加了杉木的Fv/Fm、Fv/F0、Y(II)值; 高磷处理下, 低氮添加显著增加了杉木的F0、qP、qN值, 降低了杉木的Fv/Fm值; 高磷处理下, 高氮添加显著增加了杉木的Fm、Fv/F0值。如图3(b、d、f、h、j、l、n)所示, 在秋季, 单独添加磷后显著抑制了杉木的Fv/Fm、Fv/F0、qP、Y(II)值(<0.05); 单独添加磷后杉木的Fm、F0值在低磷处理下显著增加, 而在高磷处理下显著降低; 单独添加氮后显著抑制了杉木的Fm值(<0.05); 磷添加处理下, 低氮添加显著降低了杉木的F0值; 低磷处理下, 高氮添加显著增加了杉木的qN值, 降低了杉木的F0值; 高磷处理下, 低氮添加显著增加了杉木的qN值, 降低了杉木的Fm值; 高磷处理下, 高氮添加显著增加了杉木的Fm、F0值, 降低了杉木的Fv/Fm、Fv/F0、qN、Y(II)值。如图3(a、c、e、g、i、k、m)与图3(b、d、f、h、j、l、n)所示, 单独添加氮后杉木的qN值在低氮情况下显著增加而高氮情况下显著降低(<0.05)。
3.4 杉木叶片N、P含量与光合、叶绿素荧光参数的相关性分析
从表1得出: 在夏季, 杉木叶片N含量与叶色值有显著正相关; 杉木叶片P含量与最大净光合速率有显著正相关。在秋季, 杉木叶片N含量与叶色值和光化学淬灭系数有显著负相关, 与最大荧光产量和初始荧光产量有显著正相关; 杉木叶片P含量与最大净光合速率有显著正相关。
4 讨论
4.1 磷添加对杉木生理特征的影响
磷是作物生长发育所必需的大量元素之一, 缺磷可以使作物的形态指标、产量发生变化, 同时可以引起作物发生一系列的生理生化反应[25]。本研究中, 在夏、秋季里磷添加对杉木的最大净光合速率无显著影响, 说明实验添加的单因素磷不足以直接影响杉木的最大净光合速率。原因可能是当地缺磷情况十分严重, 当磷供给量处于较低水平并成为限制因素时, 对植物叶片的扩展产生显著影响, 即叶片总叶面积、叶片生长速率和叶片数量显著减少, 并且植物体内因缺磷引起光合磷酸化过程中有机磷循环受阻, 从而严重影响植物的光合作用[26]。大量研究表明, 在磷胁迫下, 植物的光合作用、生长以及生物量分配受到影响[27]。
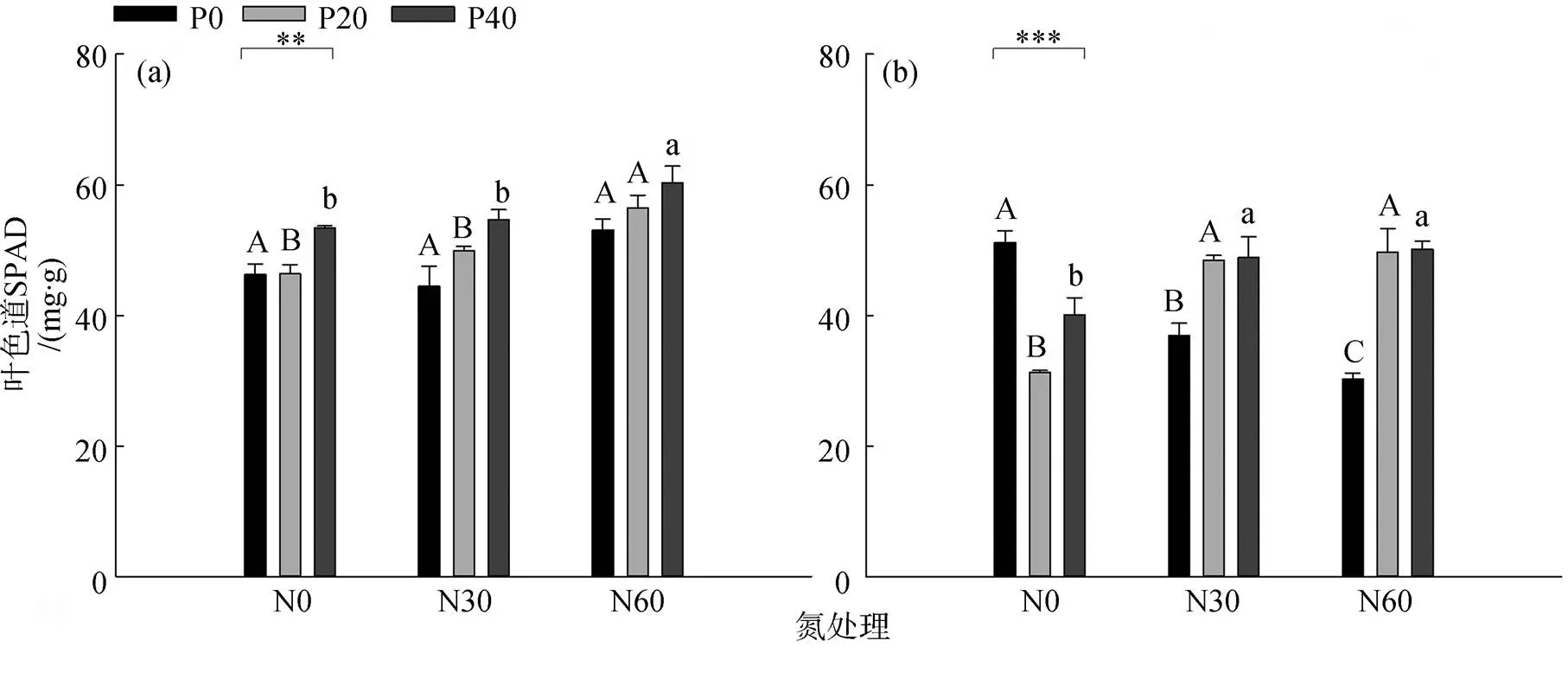
图2 不同氮沉降与磷添加处理下夏(a)、秋(b)季杉木的叶色值


表1 夏季和秋季杉木叶片N、P含量与光合、叶绿素荧光参数的相关性分析
注: *表示<0.05; **表示<0.01; ***表示<0.001; Pn max: 最大净光合速率; SPAD: 叶色值; Fm: 最大荧光产量; F0: 初始荧光产量; Fv/Fm: 最大PSII 的光能转换效率; Fv/F0: PSII 潜在活性; qP: 光化学淬灭系数; qN: 非光化学淬灭系数; Y(II): 实际量子产量。
植物叶片的叶绿素含量与SPAD存在显著正相关[28], 且叶绿素是植物光合作用的主要色素, 也是影响植物生长的重要因素之一, 其含量反映了植物的光合速率水平[29]。本研究中, 夏季杉木的SPAD值随磷的增加而上升, 说明磷添加增加了杉木叶片的叶绿素含量。有学者研究表明, 施用磷肥可以增加小麦[30]、大豆[31]等农作物的SPAD值, 从而促进其生长。但在秋季磷添加降低了杉木的SPAD值, 这与刘俊良[32]等研究结果一致。这是秋季杉木叶片中P含量相较于夏季大幅减少造成的, 叶片P含量降低导致杉木自身叶绿素含量下降, 另外即使夏季向土壤中施加磷肥, 但经过时间的流逝绝大部分磷亦为土壤所固定[33], 因此杉木的生长发育状况仍受到磷供给的限制[34]。
叶绿素荧光常用来判断逆境胁迫等因素对植物光合作用造成的伤害[35]。大量研究表明, 低磷胁迫使小麦、玉米、水稻等植物的叶绿素荧光均发生明显的变化[11, 36]。本研究中, 在夏季, 杉木的Fm、F0、Fv/F0值随磷的增加呈现下降趋势, 表明杉木的PSII光化学潜在活力减弱, 这可能与Fm和F0极易受到天气的影响有关[37]。在秋季, 杉木的Fv/Fm、Fv/F0、qP、Y(II)值显著降低, 表明PSII潜在活性减弱, 且PSII反应中心的开放程度明显减小, 减弱了该反应中心光合电子的传递能力, 导致光合效率下降; 杉木荧光参数Fv/Fm值下降与郭延平等[38]测得缺磷条件下蜜柑叶片Fv/Fm值明显降低的结果一致, 说明该地在秋季缺磷严重, 即使添加磷后, 杉木仍处于低磷胁迫的环境。
4.2 氮沉降处理对杉木生理特征的影响
氮元素影响了陆地植物的光合作用, 研究表明提高土壤中的氮素含量, 能够显著促进植物的生长和生产力[39]。本研究中, 在夏季, 不同氮沉降强度处理下杉木的最大净光合速率均无显著差异, 与Sheriff和Reich等人的研究结果一致[40]。原因是初期添加的氮元素还不足以影响杉木的最大净光合速率。在秋季, 氮沉降处理下杉木的最大净光合速率显著高于对照组, 表明氮沉降促进了杉木的最大净光合速率, 增强光合作用。这是由于在累积效应下, 氮沉降引起杉木叶片氮含量增加[41], 促进了植物光合色素的合成从而提高植物的光合能力[42]。Nakaji[43]等结果发现日本柳杉幼苗的净光合速率随氮输入量的增加而增加, 这与本文的研究结果一致。
本研究中, 在秋季, 杉木的SPAD值随着氮沉降的增加呈现下降趋势, 同时由表1可知: 在秋季, 杉木叶片的N含量与SPAD呈负显著相关, 且杉木叶片N含量增加, 故导致SPAD值下降, 表明在秋季氮沉降抑制了杉木叶绿素的合成。这与蔡瑞国[44]等人的研究结果一致。原因可能是因为边际效应使得杉木的SPAD值降低。也可能与杉木自身原因相关, 有研究认为[45]植物叶绿素含量之所以会降低, 可能是由于作物吸收外界金属元素, 在类囊体上沉积, 并且与膜上蛋白体结合进而阻碍叶绿素的合成和破坏叶绿体酶系统。
本研究中, 在夏季, Fm、F0、Fv/F0值随氮沉降的增加呈现下降趋势, 表明杉木受模拟氮沉降影响后PSII潜在光合作用活性减弱和光合效率降低, 这可能与Fm和F0极易受到天气的影响有关[37]。在夏秋季里杉木的qN值在低氮情况下都显著上升, 表明低氮处理下光保护机制启动, 为避免PSII受到伤害。张雷明等[46]研究发现施氮能够提高小麦qN值, 增强了过剩光能的非化学耗散, 有利于保护光合机构, 缓解环境对光合作用的影响, 这与本次研究结果一致。环境的复杂多变, 使得杉木必须在生理上也发生与之相应的改变来适应变化的环境。
4.3 磷处理下添加氮对杉木生理特征的影响
本研究中, 在秋季, 高磷处理下添加氮后杉木的最大净光合速率呈上升趋势。说明氮磷的添加促进杉木的最大净光合速率, 这与王奇峰等人研究发现氮磷交互对107杨的最大净光合速率有一定的促进作用的结果一致, 原因是氮、磷供给通过增加植物叶片叶绿素含量, 来影响光系统PSII对光能的吸收和传递效率, 从而影响植物叶片的净光合速率[47]。
氮、磷添加可以促进叶绿素的合成[48]。本研究中, 在夏秋季, 磷处理下添加氮, 杉木的SPAD值总体都呈上升趋势。说明氮磷添加在一定程度上增加了杉木的叶色值, 且缺失任何一种元素均不利于叶绿素的合成。氮磷在叶片生化反应进程中都起着重要的作用, 其缺乏或过量都会导致叶绿素含量、同化力合成、酶含量和酶活性的下降[49], 这与我们研究发现的结果类似。
氮、磷的供给不仅能影响叶绿素的分配格局, 而且影响光系统PSII的合成, 进而影响植物对光能的吸收、利用[47]。本研究中, 氮磷复合提高了杉木叶片部分荧光参数, 表明氮磷添加促进了杉木光合荧光特性, 原因可能是施加氮磷有利于改善植物叶片的光合能力, 提高了PSII的活性、PSII反应中心的开放部分比例和PSII光化学最大效率, 使PSII光化学量子产量和表观光合作用电子传递速率得到提高, 减小了非辐射能量的耗散, 从而使植物叶片所吸收的光能可以较充分的用于光合作用[50], 进而促进杉木生长。
5 结论
单独磷添加未能影响杉木的最大净光合速率, 而对SPAD值产生显著影响, 这种影响随着季节而产生变化。在秋季, 氮沉降提高了杉木的最大净光合速率, 但不利于杉木叶绿素的合成。在秋季, 高磷情况下, 氮增加促进了杉木的光合作用和SPAD值, 并且改善了PSII的活性、PSII反应中心的开放部分比例和PSII光化学最大效率, 使PSII光化学量子产量得到提高, 减小了非辐射能量的耗散, 从而使杉木叶片所吸收的光能可以较充分的用于光合作用。夏季杉木叶片N含量与叶色值呈显著正相关, 秋季杉木叶片N含量与Fm和F0呈显著正相关。夏秋季杉木叶片P含量与最大净光合速率呈显著正相关。实验结果表明在磷添加的背景下, 大气氮沉降有利于提高杉木林的生产力。
[1] MARSCHNER H. Mineral nutrition of higher plants (2nd Edn)[J]. London: Academic Press, 1995.
[2] 陈竑竣, 李贻铨, 陈道东, 等. 杉木人工林土壤磷素形态及其有效性研究[J]. 林业科学研究, 1996, 9(2): 121–l26.
[3] HINSINGER P. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review[J]. Plant Soil, 2001, 237: 173–195.
[4] 国家林业局. 全国森林资源统计 (1999–2003) [M].北京: 中国林业出版社, 2005.
[5] 樊后保, 廖迎春, 刘文飞, 等. 模拟氮沉降对杉木幼苗养分平衡的影响[J]. 生态学报, 2011, 31(12): 3277–3284.
[6] LIMA J D, DA MSTTA F M, MOSQUIM P R. Growth attributes, xylem sap composition, and photosynthesis in common bean as affected by nitrogen and phosphorus deficiency[J]. Journal of Plant Nutrition, 2000, 23(7): 937–947.
[7] 王帅, 杨劲峰, 韩晓日, 等. 不同施肥处理对旱作春玉米光合特性的影响[J]. 中国土壤与肥料, 2008, (6): 23–27.
[8] 潘晓华, 刘水英, 李锋, 等. 低磷胁迫对不同水稻品种幼苗光合作用的影响[J]. 作物学报, 2003, 29(5): 770–774.
[9] 张海伟, 徐芳森. 不同磷水平下甘蓝型油菜光合特性的基因型差异研究[J]. 植物营养与肥料学, 2010, 16(5): 1196–1202.
[10] 郭淑青, 李文金, 张仁懿, 等. 氮磷添加对金露梅叶片化学计量及光合特征的影响[J]. 广西植物, 2014, 34(5): 629–634.
[11] 张建恒, 李宾兴, 王斌, 等. 不同磷效率小麦品种光合碳同化和物质生产特性研究[J]. 中国农业科学, 2006, 39(11): 2200–2207.
[12] 周晓兵, 张元明, 等. 干旱半干旱区氮沉降生态效应研究进展[J]. 生态学报, 2009, 29(7): 3835–3845.
[13] PALANI S, TKALICH P, BALASUBRAMANIAN R, PALANICHAMY J. Ann application for prediction of atmospheric nitrogen deposition to aquatic ecosystems[J]. Marine Pollution Bulletin, 2011, 62(6): 1198–1206.
[14] CORNELL S E. Atmospheric nitrogen deposition: Revisiting the question of the importance of the organic component[J]. Environmental Pollution, 2011, 159(10): 2214–2222.
[15] STEVENS C J, DUPRE C, DORLAND E, et al. The impact of nitrogen deposition on acid grasslands in the Atlantic region of Europe[J]. Environmental Pollution, 2011, 159(10): 2243–2250.
[16] AMBALAVANAN J. Absorption of nitrogen, oxygen and Argon on Na-CeX Zeolites Kluwer academic publishers, manufactured in the Netherlands absorption J[J]. Physiologia Plantarum, 2002, 8: 271–278.
[17] LU Chaoqun, TIAN Hanqin. Spatial and temporal patterns of nitrogen deposition in China: synthesis of observational data[J]. Journal of Geophysical Research-Atmospheres, 2007, 112: 141–167.
[18] 顾鸿昊, 翁俊, 孔佳杰, 等. 粗放和集约经营毛竹林叶片的生态化学计量特征[J]. 浙江农林大学学报, 2015, 32(5): 661–667.
[19] BOXMAN A W, BLANCK K, BRANDRUD T E. Vegetation and soil biota response to experimentally- changed nitrogen inputs in coniferous forest ecosystems of the NIREX project[J]. Forest Ecology and Management, 1998, 101(1): 65–79.
[20] 李德军, 莫江明, 方运霆, 等. 模拟氮沉降对三种南亚热带树苗生长和光合作用的影响[J]., 2005, 29(4): 543–549.
[21] Evans J R. Photosynthesis and nitrogen relationships in leaves of C3plants[M]. Oecologia, 2002, 78: 9–19.
[22] MITSUKAWA N, OKUMURA S, SHIRANO Y, et alOverexpression of an Arabidopsis thaliana high-affinity phosphate transporter gene in tobacco cultured cells enhances cell growth under phosphate-limited conditions[M]. Proceeding of the National Academy of Sciences of United States of America, 1997, 94(13): 7098–7102.
[23] WHITEHEAD S J, CAPORN S J M, PRESS M C. Effects of elevated CO2, nitrogen and phosphorus on the growth and photosynthesis of two upland perennials: Calluna vulgaris and[J]. New Phytologist, 1997, 135(2): 201–211.
[24] 殷秀敏, 余树全, 江洪, 等. 模拟酸雨对两种针叶植物气体交换和叶绿素荧光特征的影响[J]. 浙江林业科技, 2009, 29(5): 13–18.
[25] 董秋平, 赵恢, 张小芳, 等. 低磷胁迫下不同野生大豆的形态和生理响应差异[J]. 江苏农业科学, 2017, 45(9): 79–83.
[26] MOLLIER A, PELLERIN S. Maize root system growth and development as influenced by phosphorous deficiency[J]. Journal of Experimental Botany, 1999, 50: 487–497.
[27] 吴楚, 范志强, 王政权. 磷胁迫对水曲柳幼苗叶绿素合成、光合作用和生物量分配格局的影响[J]. 应用生态学报, 2004, 15(6): 935–940.
[28] 李绍长, 胡昌浩, 龚江, 等. 低磷胁迫对磷不同利用效率玉米叶绿素荧光参数的影响[J]. 作物学报, 2004, 30(4): 365–370.
[29] 林静雯, 李莹, 罗洁文, 等. 草甘膦对杉木种子萌发及幼苗生长的毒性效应[J]. 江西农业大学学报, 2015, 37(5): 843–848.
[30] 阳显斌, 张锡洲, 李延轩. 施磷量对不同磷效率小麦氮、磷、钾积累与分配的影响[J].核农学报, 2012, 26(1): 141–149.
[31] 董薇, 练云, 余永亮. 大豆耐低磷胁迫研究进展[J]. 大豆科学, 2012, 31(1): 135–140.
[32] 刘俊良, 华元刚, 罗微, 等. 施磷对开割橡胶树根系生长发育的影响[J]. 热带作物学报, 2011, 32(8): 1393–1398.
[33] HOLFORD I C R. Soil phosphorus: its measurement, and its uptake by plants[J]. Australian Journal of Soil Research, 1997, 35: 227–239.
[34] RAGOTHAMA K C. Phosphate acquisition,[J]. Molecular Biology, 1999, 50: 665–693.
[35] 张毅龙, 张卫强, 甘先华, 等. 低温胁迫对6种珍贵树种苗木光合荧光特性的影响[J]. 生态环境学报, 2014, 23(5): 777–784.
[36] LI Dejun, MO Jiangming, FANG Yunting, et al. Ecophysiological responses of woody plants to elevated nitrogen deposition[J]. Journal of Tropical and Subtropical Botany, 2004, 12(5): 482–488.
[37] 刘建福. 澳洲坚果叶片光合速率和叶绿素荧光参数日变化[J]. 西南农业大学学报: 自然科学版, 2006, 28(2): 271–274.
[38] 郭延平, 陈屏昭, 张良诚, 等. 缺磷胁迫加重柑橘叶片光合作用的光抑制及叶黄素循环的作用[J]. 植物营养与肥力学报, 2003, 9(3): 359–363.
[39] XIA Jianyang, WAN Shiqiang. Global response patterns of terrestrial plant species to nitrogen addition[J]. New Phytologist, 2008, 179: 428–439.
[40] SHERIFF D W, NAMBIAR E K S, FIFE N D. Relationships between nutrient status, carbon assimilation and water use efficiency in Pinus radiate needles[J]. Tree Physiology, 1986, 2: 73–88.
[41] SONG Xinzhang, GU Honghao, MENG Wang, et al. Management practices regulate the response of Moso bamboo foliar stoichiometry to nitrogen deposition[J]. Scientific Reports, 2016, 6: 24107.
[42] 贾瑞丰, 尹光天, 杨锦昌, 等. 不同氮素水平对红厚壳幼苗生长及光合特性的影响[J]. 林业科学研究, 2012, 25(1): 23–29.
[43] NAKAJI T, TAKENAGA S, KUROHA M, et al. Photosynthetic response ofseedlings to high nitrogen load[J]. Environmental Sciences, 2002, 9(4): 269–282.
[44] 蔡瑞国, 张敏, 戴忠民, 等. 施氮水平对优质小麦旗叶光合特性和子粒生长发育的影响[J]. 植物营养与肥料学报, 2006, 12(1): 49–55.
[45] STOBART A K, GRIFFITHS W T, I AMEEN B, et al. The effect of Cd2+on the biosynthesis of chlorophyll in leaves of barley[J]. Physiologia Plantarum, 1985, 63: 293–298.
[46] 张雷明, 上官周平, 毛明策, 等. 长期施氮对旱地小麦灌浆期叶绿素荧光参数的影响[J]. 应用生态学报, 2003, 14(5): 695–698.
[47] 王奇峰, 徐程扬. 氮、磷对107杨叶片光合作用的影响[J]. 西北林学院学报, 2007, 22(4): 9–12.
[48] 武维华主编. 植物生理学[M]. 北京: 科学出版社, 2004.
[49] 徐培洲, 李云, 袁澎, 等. 叶绿素缺乏水稻突变体中光系统蛋白和叶绿素合成特性的研究[J].中国农业科学, 2006, 39(7): 1299–1305.
[50] 张旺锋, 勾玲, 王振林, 等. 氮肥对新疆高产棉花叶片叶绿素荧光动力学参数的影响[J]. 中国农业科学, 2003, 36(8): 893–898.
Effects of nitrogen deposition and phosphorus addition on photosynthesis and chlorophyll fluorescence characteristics of Chinese fir
QIAN Shuyue, WANG Qiao, LIU Ning, LI Quan, LEI Zhaofeng, SONG Xinzhang, AI Jianguo*
The State Key Laboratory of Subtropical Silviculture, Zhejiang A&F University, Lin’an 311300, China
In order to investigate the changes of photosynthesis and chlorophyll fluorescence parameters of Chinese fir () under atmospheric nitrogen (N) deposition and phosphorus (P) addition, our experiment was conducted on 10-year-old Chinese fir by seting up 9 treatment levels: low N (N30: 30 kg·ha-1·a-1), high N (N60: 60 kg·ha-1·a-1), low P (P20: 20 mg·kg-1), high P (P40: 40 mg·kg-1), low N and low P (N30 + P20), low N and high P (N30 + P40), high N and low P (N60 + P20), high N and high P (N60 + P40) and control group (CK). In summer, N deposition and P addition had no significant effect on the maximum net photosynthetic rate (Pnmax) of Chinese fir. Moreover, either N deposition or P addition independently inhibited the maximum fluorescence yield (Fm), initial fluorescence yield (F0) and potential activity of PSII (Fv/F0) value of Chinese fir. P addition alone increased the leaf color value (SPAD) of Chinese fir. Under the condition of P addition, low N addition increased the Fm; high N addition increased the SPAD value but decreased the non-photochemical quenching coefficient (qN) of Chinese fir. In autumn, N deposition alone promoted the Pn max. Either N deposition or P addition independently inhibited the SPAD value of Chinese fir. Under P addition, N deposition increased the SPAD but decreased the F0value both in summer and autumn. N content of leaves was significantly and positively correlated with SPADvalue in summer (<0.01). N content of leaves was remarkably and negatively correlated with SPAD value and photochemical quenching coefficient (qP) in autumn (<0.05), but was significantly and positively correlated with Fmand F0(<0.05).
nitrogen deposition; phosphorus stress; Chinese fir; photosynthesis; chlorophyll fluorescence
10.14108/j.cnki.1008-8873.2018.05.015
Q945.79
A
1008-8873(2018)05-113-09
2018-03-27;
2018-05-27
国家十三五重点研发计划课题“人工林生产力形成的关键生理生态与环境控制机制”(2016YFD0600201)
钱树玥(1993—), 女, 浙江湖州人, 硕士研究生, 主要从事生态学研究, E-mail: 624822697@qq.com
通信作者:哀建国, 男, 博士, 副教授, 主要从事生态学研究, E-mail: 1572783600@qq.com
钱树玥, 王巧, 刘宁, 等. 氮沉降和磷添加对杉木光合及叶绿素荧光特征的影响[J]. 生态科学, 2018, 37(5): 113-121.
QIAN Shuyue, WANG Qiao, LIU Ning, et al. Effects of nitrogen deposition and phosphorus addition on photosynthesis and chlorophyll fluorescence characteristics of Chinese fir[J]. Ecological Science, 2018, 37(5): 113-121.
