小檗碱体外对三转基因阿尔茨海默症小鼠海马神经元内质网应激及凋亡的影响
2018-11-02龚琼欧阳雅帆黄艳露黄敏
龚琼,欧阳雅帆,黄艳露,黄敏
(中山大学附属第七医院,广东深圳 518107)
阿尔茨海默症(AD)是一种以认知功能障碍和学习记忆能力减退为临床症状的神经退行性疾病,脑组织淀粉样蛋白沉积(Aβ)和Tau蛋白纤维缠结是其病理基础[1]。内质网应激是AD病因学中的一个重要过程,致病性错误折叠蛋白质的积累和细胞内钙信号的破坏是诱发内质网应激的基础机制,可导致神经元功能障碍和细胞死亡[2,3]。在有害或者应激条件下,细胞通过激活未折叠蛋白质反应(UPR)的适应性反应途径逃避严重损害。葡萄糖相关蛋白78(GRP78)是通过关键途径启动UPR的主传感器[4]。在内质网应激时,错误折叠蛋白抑制GRP78和传感蛋白之间的相互作用,从而启动UPR信号传导,产生细胞凋亡作用[5]。小檗碱是从黄连素中提取的一种天然异喹啉类生物碱,具有多种药理作用[5],如抗炎、调节血糖、抗病毒、调脂等,而且对神经系统中自身免疫性疾病、脑血管疾病、癫痫、焦虑抑郁、AD等疾病亦有一定改善作用[6,7]。最近研究表明,小檗碱能降低AD中Aβ沉积和改善三转基因AD小鼠的学习记忆能力[8]。2016年7月~2018年3月,本研究观察了小檗碱对三转基因AD小鼠海马神经元内质网应激及凋亡的影响,现分析结果并探讨其作用机制。
1 材料与方法
1.1 材料 三转基因AD小鼠购自美国Jax实验室,能稳定表达人源突变基因APPSwe、tauP301L以及鼠源突变基因PSl[9,10]。小檗碱购自美国Sigma公司,纯度约99%。原代细胞培养皿购自美国Invitrogen公司,DMEM/F12(1∶1)细胞培养液购自美国Gibco公司,MTT试剂盒、细胞裂解液、乳酸脱氢酶(LDH)细胞毒性试剂盒、TUNEL细胞凋亡检测试剂盒均购自沈阳万类生物科技有限公司,二甲基亚砜(DMSO)和3-(4,5-二甲基噻唑-2)-2,5-二苯基四氮唑溴盐(MTT)购自美国Sigma公司,GRP78、Caspase-3和Bcl-2、Bax以及GAPDH抗体、兔鼠通用二抗均购自美国CST公司。
1.2 原代海马神经元分离和培养 取出生24 h内的三转基因AD小鼠,75%乙醇全身消毒,在显微镜下无菌条件取出海马组织,剔除血管膜,用眼科剪剪碎,加入0.125%胰蛋白酶,37 ℃消化30 min,用含10%FBS的DMEM/F12完全培养基终止消化。加入含血清的培养基(高糖EMEM+2 mmol/L谷氨酰胺+0.1 U/mL胰岛素+10%FBS),吹打混匀3次,每次吹打后静置3~5 min。取上清液,200目细胞筛过滤,4 ℃、1 000 r/min离心5 min。弃掉上清,加入神经元培养基,巴氏管吹打混匀细胞,将细胞接种至培养皿、96孔板中、玻片上、6孔板中(已用0.1 mg/mL多聚赖氨酸包被),分别加入完全培养皿中,置于37 ℃、5% CO2的培养箱中培养。12 h后更换为无血清的培养基,培养48 h加入阿糖胞苷(终浓度为10 μmol/L),以抑制非神经元细胞的过度生长,每3~4天换液1次,倒置显微镜下观察神经元生长状况。
1.3 海马神经元分组及处理 在上述1.2神经元培养第7天时,随机分为1 μmol/L小檗碱组、10 μmol/L小檗碱组、对照组,调整细胞密度为5×104/孔,每组设6个复孔,前两组分别加入1、10 μmol/L小檗碱两种浓度(小檗碱溶于DMSO中),对照组加入等体积的DMSO,干预24 h,收集细胞进行后续实验。
1.4 海马神经元细胞凋亡率检测 采用TUNEL法。将上述三组神经元培养基弃掉,PBS洗涤10 min×3次,4%多聚甲醛固定样本爬片1 h;PBS洗涤玻片5 min×3次,室温下封闭液封闭玻片30 min;PBS洗涤玻片5 min×3次,加入含0.1%~0.2% Triton X-100的PBS,冰浴5 min。将50 μL酶溶液与400 μL的标记液混合配制成500 μL TUNEL反应试剂,充分混匀后在玻片上滴加50 μL TUNEL反应试剂,37 ℃、避光、湿盒中孵育1 h。PBS洗涤玻片5 min×3次,滴加DAPI,5 min后吸出。PBS洗涤玻片5 min×3次,滴加抗淬灭剂50 μL,封片。采用Image J软件统计凋亡细胞数目。细胞凋亡率=凋亡细胞数/细胞总数×100%。
1.5 海马神经元细胞存活率检测 采用MTT法。以5×103/孔的细胞密度接种于96孔细胞培养板中,培养板中事先用多聚赖氨酸处理,培养第7天时加入无血清的培养基洗涤3次,1 μmol/L小檗碱组、10 μmol/L小檗碱组分别加入1、10 μmol/L小檗碱,对照组加入等体积的DMSO,每组设6个复孔,检测小檗碱作用24 h时,各组细胞相对存活率。采用酶标仪(波长570 nm)检测各孔吸光度(A)值。对照组细胞存活率标化为100%,计算细胞相对存活率。细胞相对存活率(%)=(实验孔A570值/对照组A570值)×100%。
1.6 海马神经元培养上清液LDH释放率检测 取上述3组培养基上清液,4 ℃、1 000 r/min离心5 min,收集上清液,其他步骤按照LDH细胞毒性试剂盒说明书操作,酶标仪(波长490 nm)检测小檗碱干预24 h各孔A490nm值。LDH释放率(%)=(处理样品A490值-样品对照孔A490值)/(细胞最大酶活性A490值-样品对照孔A490值)×100%。
1.7 海马神经元细胞GRP78、Caspase-3、Bcl-2、Bax蛋白表达检测 采用Western blotting法。将上述1.3中的3组细胞培养基弃掉,用PBS洗涤3次,加入简易型细胞裂解液(Lysis Buffer+PMSF+蛋白酶抑制剂)200 μL至6孔板,充分吹打,然后超声破碎、离心,提取上清液,BCA法蛋白定量合格。然后进行SDS凝胶电泳,电泳结束将蛋白电泳产物转印至PVDF膜上,5%脱脂奶粉封闭2 h,TBST洗涤10 min×3次,分别加入一抗[GRP78(1∶1 000)、Caspase-3(1∶3 000)、Bcl-2(1∶1 000)、Bax(1∶1 000)、GAPDH(1∶3 000)],4 ℃过夜;次日PBST洗涤10 min×3次,室温下加入辣根过氧化物酶标记的二抗(1∶5 000)。TBST洗涤10 min×3次,采用万类ECL发光液显影,GraphPad Prism5分析作图,数据图片采用Quanlity One软件扫描。以GAPDH为内参,计算目的蛋白相对表达量。目的蛋白相对表达量=目的蛋白电泳条带灰度值/GAPDH蛋白电泳条带灰度值。
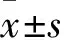
2 结果
2.1 各组海马神经元细胞凋亡率比较 1 μmol/L小檗碱组、10 μmol/L小檗碱组、对照组海马神经元细胞凋亡率分别为(23.43±3.21)%、(16.72±4.02)%、(43.59±3.68)%,组间两两比较P均<0.05。
2.2 各组海马神经元细胞存活率比较 1 μmol/L小檗碱组、10 μmol/L小檗碱组、对照组海马神经元细胞存活率分别为(48.25±12.09)%、(67.16±11.41)%、(39.57±13.54)%,组间两两比较P均<0.05。
2.3 各组海马神经元培养上清液LDH释放率比较 1 μmol/L小檗碱组、10 μmol/L小檗碱组、对照组海马神经元培养上清液LDH释放率分别为(12.43±3.41)%、(8.23±2.58)%、(18.79±4.23)%,组间两两比较P均<0.05。
2.4 各组海马神经元细胞GRP78、Caspase-3、Bcl-2、Bax蛋白表达比较 见表1。
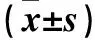
表1 各组海马神经元细胞GRP78、Caspase-3、Bcl-2、Bax蛋白相对表达量比较
注:与对照组相比,*P<0.01;与1 μmol/L小檗碱组相比,#P<0.05。
3 讨论
AD发病的因素包括Aβ沉积、Tau蛋白过度磷酸化、活性氧自由基增多等,这些因素均能导致内质网的错误折叠蛋白增加,从而引发内质网应激[3]。早期内质网应激是细胞对伤害的适应性反应,持久或过强的内质网应激则可导致细胞凋亡。有研究表明,抑制过度的内质网应激能明显减少细胞凋亡。GRP78是诱导内质网应激发生的关键分子蛋白,是内质网应激的标志性蛋白,可调节内质网应激的错误折叠蛋白功能,从而抑制内质网损伤。有研究表明,在氧化应激失衡、缺血缺氧等状态下内质网会激活细胞内的UPR,从而激活转录因子6和启动GRP78基因,上调GRP78蛋白表达,进而改善内质网功能[3,4]。
本研究结果发现,与对照组比较,1 μmol/L小檗碱组、10 μmol/L小檗碱组细胞凋亡率、LDH释放率下降,细胞存活率增加,且10 μmol/L小檗碱组变化更明显。表明小檗碱能降低神经元细胞凋亡率、抑制LDH释放水平,对三转基因AD小鼠海马神经元具有明确的保护作用,且具有量效关系。
目前小檗碱发挥神经保护作用的具体机制还不清楚。有文献报道,小檗碱能通过增加自噬水平LC3-Ⅱ、Beclin-1表达,降低Aβ沉积和改善认知功能,同时减少海马区神经元凋亡[8]。此外,Liang等[11]研究表明,小檗碱能通过降低Aβ25-35诱导体外原代神经元线粒体细胞色素C水平,提高其抗氧化能力,从而降低神经元凋亡。本研究进一步探索小檗碱对AD神经元的保护作用是否与抑制内质网应激有关。
本研究结果发现,1 μmol/L小檗碱组、10 μmol/L小檗碱组GRP78蛋白表达高于对照组,且10 μmol/L小檗碱组GRP78蛋白表达更高,表明小檗碱可呈剂量依赖性上调GRP78蛋白表达,从而改善内质网功能。内质网应激可激活UPR,UPR分别是由GRP78蛋白和RNA激活蛋白样内质网激、ATF-6等组成,能启动内质网应激进而介导凋亡信号通路,激活下游信号通路,如Caspase、CHOP、Bcl-2家族等[12]。Caspase-3在执行凋亡过程中发挥重要作用,最终导致细胞凋亡[13,14]。本研究结果发现,1 μmol/L小檗碱组、10 μmol/L小檗碱组较对照组Caspase-3、Bax蛋白表达降低,Bcl-2蛋白表达增加。由此推测,小檗碱可增加GRP78蛋白及抑制Caspase-3蛋白表达,帮助纠正及重组错误折叠蛋白、细胞未折叠的正确组装,抑制内质网应激;同时上调抗凋亡蛋白Bcl-2表达及下调促凋亡蛋白Bax表达,减少神经细胞凋亡,对AD神经元发挥保护作用。但小檗碱何种浓度对AD神经元有最佳保护作用仍需进一步研究。
综上所述,小檗碱体外可抑制三转基因AD小鼠海马神经元内质网应激,减少神经元凋亡,保护三转基因AD小鼠海马神经元。其机制可能与上调GRP78蛋白、抗凋亡蛋白Bcl-2表达及抑制Caspase-3蛋白、促凋亡蛋白Bax表达有关。干预内质网应激相关蛋白GRP78表达,可能对AD治疗具有潜在意义。
