人为干扰对中亚热带森林生物量及其空间分布格局的影响
2018-10-18李尚益陈金磊李雷达刘兆丹张仕吉
李尚益,方 晰,2,*,陈金磊,李雷达,辜 翔,刘兆丹,张仕吉
1 中南林业科技大学生命科学与技术学院,长沙 410004 2 湖南会同杉木林生态系统国家野外科学观测研究站,会同 438107
森林生物量是研究和评价森林生态系统结构与功能过程最基本的参数,不仅能揭示森林生态系统能量平衡、养分循环和生产力等功能过程的变化规律[1- 3],而且能反映生态系统功能的强弱,对研究森林生态系统碳循环过程和全球气候变化具有重要意义[4- 7]。20世纪70年代初,在国际生物学计划(International Biological Programme,IBP)和人与生物圈计划(Man and the Biosphere Programme,MAB)[8]推动下,森林生物量研究迅速发展,研究方法和技术多样,生物量估算准确度不断提高,特别是20世纪90年代国际地圈-生物圈计划(International-Geosphere-Biosphere Programme,IGBP)的实施以及国际社会对全球气候变化的关注,森林生物量研究再次成为现代生态学研究热点。目前,森林生物量研究已在个体、种群、群落、生态系统、景观、区域、生物圈等多个尺度上进行了研究[9-10],基于样地调查[11-12]、遥感技术[6]、激光雷达技术[2]和生态过程模型[13]对不同地理种源[11-13]、发育阶段[11,14]和自然地带[15-17]的生态系统生物量的估算,也有围绕气候、环境、资源等人类社会生产实践的生态系统生物量动态变化研究[18-19]。但基于样地调查数据,对不同程度的人为干扰和保护对森林生物量影响的研究仍少见报道。
中国亚热带地区水热条件优越,森林类型多样和物种丰富[20],是中国重要的农林业生产基地,也是中国森林受人类干扰最早和最为严重的地区之一。常绿阔叶林是该地区的地带性植被类型,是亚热带陆地生态系统的重要组成部分[21-22],但由于人为干扰严重,取而代之的是大面积的次生林、次生灌丛和灌草丛等[23],群落结构趋于简单,抗外界干扰能力下降、涵养水源和土壤肥力质量减弱等特征。研究森林生态系统结构和功能过程对不同人为干扰程度的响应,无疑对揭示森林生态系统在人为干扰下的退化机制,探讨退化植被恢复和重建具有理论和实践双重意义。目前国内多数研究将重点放在不同程度的人为干扰和保护对群落结构和物种多样性[24-28],土壤肥力质量和蓄水能力[29-31]等方面的研究,在人为干扰对森林生物量影响方面的研究,也主要集中在人为经营管理措施(如采伐、抚育间伐、造林、施肥等)对人工林生物量影响的研究[32-35],而对处于不同人为干扰程度的次生林、次生灌丛和灌草丛生物量的比较研究很少,对揭示人为干扰对森林生态系统结构与功能过程的影响机制仍缺乏相关数据。为此,本研究以湘中丘陵区4种处于不同程度人为干扰的植物群落:檵木(Loropetalumchinense)-南烛(Vacciniumbracteatum)-满山红(Rhododendronmariesii)灌草丛、檵木-杉木(Cunninghamialanceolata)-白栎(Quercusfabri)灌木林、马尾松(Pinusmassoniana)-石栎(Lithocarpusglaber)-檵木针阔混交林和石栎-红淡比(Cleyerajaponica)-青冈(Cyclobalanopsisglauca)常绿阔叶林为对象,研究不同程度的人为干扰后森林群落生物量的空间分布格局及其影响因素,为探讨人为干扰后的森林生态恢复机制,为准确地反映人类活动与森林生态功能过程的关系,以及为促进中亚热带森林植被恢复与保护、环境与经济可持续发展提供科学依据。
1 研究地概况
研究地设置在湖南省长沙县(113°17′—113°27′E,28°23′—113°24′N),两地空间距离约为28km,地处幕连九山脉中支连云山山脉的余脉,地形起伏较大,海拔高度为100—550 m,坡度多在20°—30°之间,属于典型的低山丘陵地貌。该区年平均气温17.0℃,7—8月份极端高温40℃,1月份极端低温-11℃,雨量充沛,相对湿度较大,年降水量1412—1559 mm,降雨集中在4—7月份,为中亚热带东南季风湿润气候。土壤以板岩和页岩发育而成的红壤为主,地理位置和自然条件决定了该区地带性植被为中亚热带常绿阔叶林,属于湘中湘东山丘盆地栲(Castanopsisfargesii)林、马尾松林、毛竹(Phyllostachysheterocycla)林、油茶(Camelliaoleifera)林及农田植被区的幕阜、连云山山地丘陵植被小区。由于该区人为干扰(采伐、火烧)频繁,强度各异,原生天然植被常绿阔叶林破坏比较严重,形成了多种处于不同程度的人为干扰植物群落(灌草丛、灌木林、马尾松针阔混交林、常绿阔叶林)以及以杉木、马尾松、湿地松(Pinuselliottii)为主的人工林群落,为中亚热带森林群落恢复(或演替)及其相关研究提供了场所。
2 研究方法
2.1 样地设置
在地域相邻,海拔、坡度、坡向以及土壤、气候条件基本一致,处于不同人为干扰程度的4种植物群落:檵木-南烛-满山红灌草丛、檵木-杉木-白栎灌木林、马尾松-石栎-檵木针阔混交林、石栎-红淡比-青冈常绿阔叶林(分别简写为:LVR、LCQ、PLL和LAG,下同)内设置长期定位观测样地:LVR群落设置4个20 m×20 m样地,LCQ群落设置3个20 m×20 m样地;PLL、LAG群落均设置3个30 m×30 m样地。4种植物群落的人为干扰的基本概况如下:
(1)LVR群落(严重干扰):1965年冬炼山、人工全垦整地,1966年春营造的马尾松人工纯林,经营期间无施肥历史,1990年皆伐后,多年遭到反复砍伐、火烧、放牧活动。2012年后,停止各种人为干扰。
(2)LCQ群落(较重干扰):1965年冬炼山、人工全垦整地,1966年春营造的杉木人工纯林,经营期间无施肥历史,1988—1989年皆伐后,每隔3—5年择伐一次。2004年后,停止择伐。
(3)PLL群落(中度干扰):20世纪70年代初,天然常绿阔叶林采伐后,自然恢复为39%针叶树马尾松,25%常绿阔叶树石栎,11%檵木和23%其他树种组成的针阔混交林。
(4)LAG群落(无明显人为干扰):20世纪60年代初以来,无明显人为干扰,保存比较完好,现为为69%常绿阔叶树(其中38%石栎,18%红淡比和5%青冈)和31%其他树种组成的常绿阔叶林。
2.2 样地群落调查
2016年10—11月落叶树种落叶前完成,采用样方调查法进行群落调查,所有植物均鉴定到种。(1)在LVR每块样地,沿对角线均匀设置4个2 m×2 m样方调查灌木层、草本层植物,记录植物名称、株数、多度、盖度、平均高度和生活型等。(2)在LCQ每块样地,沿对角线均匀设置4个5 m×5 m样方调查灌木层植物,记录植物名称、株数、多度、盖度、树高和生活型,对树高高于1.5 m的灌木,按照一定顺序编号挂牌,测定胸径(DBH)、树高、冠幅、枝下高、健康状态;对树高低于1.5 m的灌木,测定地径、树高;草本层植物调查与LVR草本层植物的调查相同。(3)在PLL、LAG每块样地,对树高高于1.5 m的木本植物,按照一定顺序编号挂牌,记录植物名称及其所在层次,测定胸径、树高、冠幅、活枝下高、健康状态;灌木层、草本层植物调查与LCQ样地灌木层、草本层植物的调查相同。
计算4种植物群落木本植物的密度、平均胸径、平均树高,Shannon-Wiener多样性指数等特征指标。4种植物群落的基本特征及其主要树种组成如表1所示。
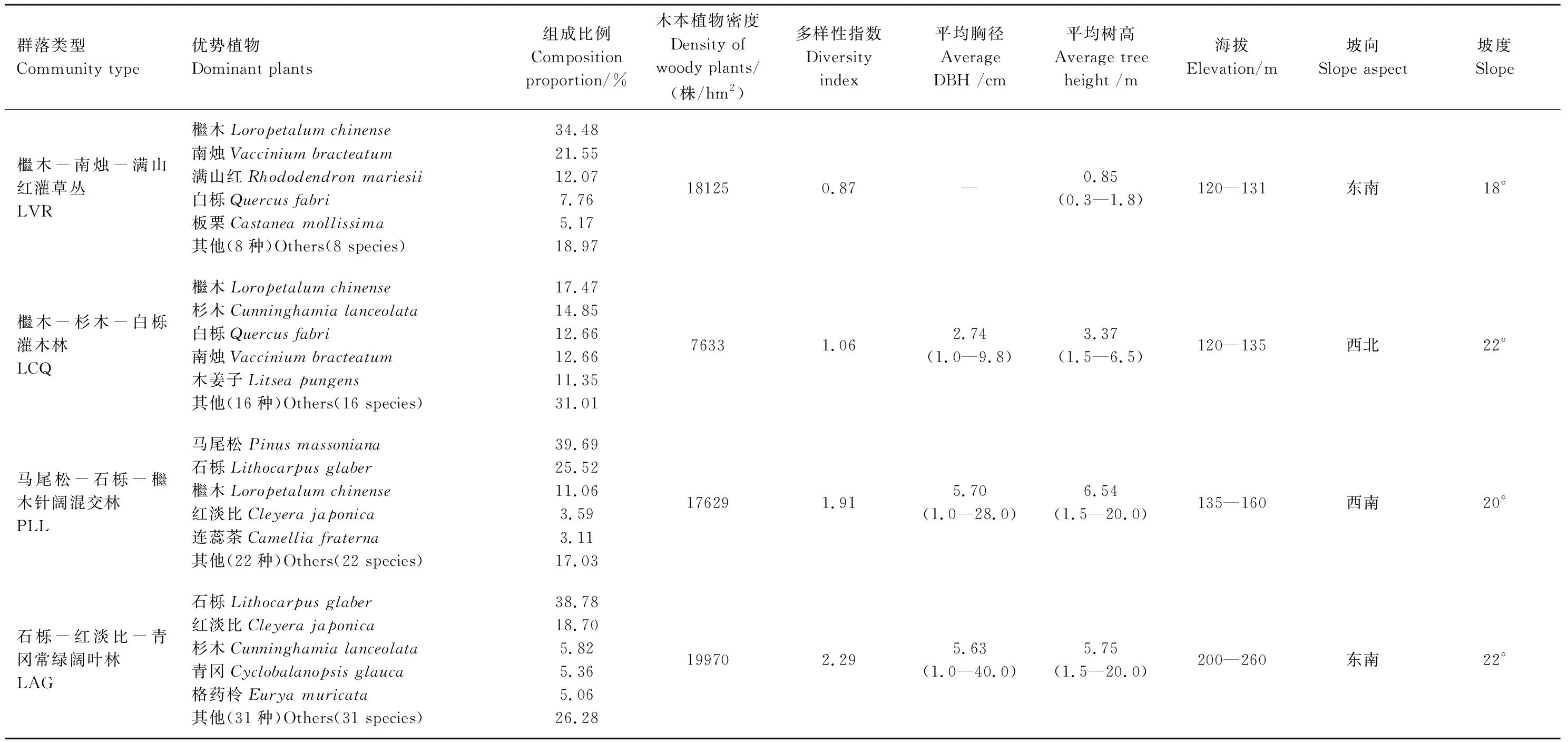
表1 样地的基本特征
LVR:Loropetalumchinense-Vacciniumbracteatum-Rhododendronmariesiiscrub-grass-land,LCQ:Loropetalumchinense-Cunninghamialanceolata-Quercusfabrishrubbery,PLL:Pinusmassoniana-Lithocarpusglaber-Loropetalumchinenseconiferous-broad leaved mixed forest,LAG:Lithocarpusglaber-Cleyerajaponica-Cyclobalanopsisglaucaevergreen broad-leaved forest
2.3 生物量的测定
在样地群落调查基础上,采用收获法测定群落生物量,2016年10—11月落叶树种落叶前完成。4种群落生物量具体的测定方法如下:
2.3.1 檵木-南烛-满山红灌草丛
在每块样地的4条边界外围随机设置1个2 m×2 m样方,记录样方中灌木、藤本植物名称、株(丛)数和草本植物名称、丛数。收割样方内的全部植物,同种灌木分为叶、枝、干、根,同种藤本植物分为叶、干和根,同种草本植物分为地上部分和地下部分,测定鲜重。将同样地的4个样方同种植物相同器官混合均匀,采集分析样品(约取0.5 kg)。
同时,设置1个1 m×1 m样方,根据地表凋落物层的分层标准[36],按未分解层、半分解层、已分解层收集样方内全部的凋落物,测定鲜重。将同样地的4个样方同一分解层的凋落物混合均匀,采集分析样品(约取1.0 kg)。
将分析样品置于80℃下烘至恒重,测定含水率。根据分析样品的含水率计算各样方干物质重量,估算样地单位面积群落的生物量。
2.3.2 檵木-杉木-白栎灌木林
根据LCQ样地群落调查数据,计算每块样地树高高于1.5 m的每种灌木平均胸径、平均树高,确定为平均标准木。在每块样地的外围,每种灌木分别选取3株平均标准木,按叶、枝、干、根测定平均标准木的鲜重,采集同一样地3株平均标准木叶、枝、干、根的分析样品(约取0.5 kg,每树种每一器官3个重复),用于含水率测定和各组分的干物质重量的换算。根据各树种9株平均标准木各组分生物量与胸径、树高的关系,构建各种灌木各组分生物量的相对生长方程(表2),计算各种灌木单株生物量。
对树高低于1.5 m灌木层、藤本层、草本层、地表凋落物层生物量的测定,与LVR群落相同。结合样地群落调查数据,估算样地单位面积群落各组分的生物量。
2.3.3 马尾松-石栎-檵木针阔混交林
根据PLL样地群落调查数据,计算每块样地每一树种的平均胸径、平均树高,在每块样地外围,每一树种分别选取3株平均标准木(即每一树种9个重复),按“分层切割法”在1.3、3.6 m处和以后2 m为1个区分断开,树梢部分不足1 m的作梢头处理,分别测定每株平均标准木叶、枝、干鲜重;地下树根采用挖掘法测定生物量,以树桩为中心在1.5 m为半径范围进行圆柱形挖掘,深度直至挖出完整的主根,按细根(<0.2 cm)、小根(0.2—0.5 cm)、粗根(0.5—2.0 cm)、大根(>2.0 cm)和根头测定树根鲜重,采集各级根系分析样品。同一样地同树种3株平均标准木各组分混合均匀后采集分析样品(约取0.5 kg,各组分3个重复)。同样,由各树种各组分含水率换算成干重,每一树种根据9株平均标准木各组分生物量与胸径、树高的关系,建立各树种各组分生物量的相对生长方程(表2),计算各乔木树种单株生物量;结合样地群落调查数据,估算单位面积每种树种各组分生物量。
2.3.4 石栎-红淡比-青冈常绿阔叶林
乔木层生物量估算:根据LAG群落调查数据,采用PLL建立的马尾松、石栎、红淡比各器官生物量相对生长方程分别估算马尾松、石栎、红淡比单株生物量,采用Ouyang等[37]建立的青冈各组分生物量的通用生长方程(表2)估算青冈单株生物量,采用刘雯雯等[38]建立的区域尺度杉木各器官生物量通用相对生长方程(表2)估算杉木单株各器官生物量,其他落叶阔叶树种、常绿阔叶树种单株各器官生物量分别采用Ouyang等[37]建立的落叶阔叶林、常绿阔叶林各组分的通用生长方程(表2)进行估算;结合群落调查数据估算样地单位面积各组分生物量。
PLL、LAG群落灌木层、藤本层、草本层、地表凋落物层生物量的测定和估算,与LVR群落的测定方法相同。
2.3.5 相对生长方程的建立
研究表明,主干比较明显的树种,以树高H(m)、胸径D(cm)或D2H作为自变量可以很好地预测其生物量。本研究样地的乔木和灌木多为主干较为明显的树种,因此以各树种各组分生物量W(g)为因变量,以树高H(m)、胸径D(cm)为自变量[39-40],选用线性函数(1)、指数函数(2)、对数函数(3)和幂函数(4)进行拟合[41],根据判定系数R2的大小选取最优相对生长方程。
y=ax+b
(1)
y=abx
(2)
y=a+blnx
(3)
y= axb
(4)
群落各层次地上部分生物量是由叶、枝、干(或茎)生物量之和构成,地下部分生物量是由根系生物量构成;整个群落地上部分生物量是由各层次地上部分生物量之和构成,地下部分生物量是由各层次根系生物量与地表凋落物层现存量之和构成。
2.4 土壤样品的采集、处理及分析方法
在2016年4月、6月、10月采集土壤样品。采集土壤样品时,将每个固定样地对角线分成3等份,在每一等份中间设置1个采样点(即每一固定样地布置3个采样点),清除采样点地上植物和死地被物,挖掘土壤剖面,按0—10、10—20、20—30 cm分层,从下至上采集土壤样品,并观察和记录土壤的颜色,同时用环刀法测定土壤容重。在室内清除土壤样品中动植物残体、石砾后,将同一固定样地的3个采样点同一土层样品等量混合均匀,约取2 kg(每种植物群落每一土层构成3—4个重复),自然风干后,分别过2、1、0.25 mm土壤筛保存于样品瓶中备用。过2 mm土壤筛的土壤样品用于测定土壤颗粒组成,过1 mm土壤筛的用于测定pH、水解氮、速效磷和速效钾含量,过0.25 mm土壤筛的用于测定有机碳、全氮、全磷、全钾含量。
(三)猪场常见寄生虫的成因调查猪寄生虫的成功控制与良好的管理有关,但事实证明,如果不结合驱虫药的正确使用将不足以控制的寄生虫感染。现已发现很多管理良好的猪场也存在着严重的蠕虫感染,这表明寄生虫有令人难以置信的生存、感染和繁殖的能力,它们几乎能在所有的猪场中生存。猪只需接触少量感染性虫卵或幼虫就可以保持感染状态,随后便感染整个猪场。
土壤颗粒组成用吸管法测定,根据国际土壤质地分类标准划分土壤质地[42],pH用土水比1∶2.5 pH计法测定,有机碳用重铬酸钾-浓硫酸水合加热法测定,全氮用KN580全自动凯氏定氮仪测定,全磷用碱熔-钼锑抗比色法测定,全钾用火焰分光光度计法测定,水解氮用碱解扩散法测定,速效磷用双酸浸提UV- 5100B紫外可见分光光度比色法测定,速效钾用醋酸铵浸提-—火焰光度法测定[42]。不同植物群落土壤理化性质如表3所示。
2.5 数据统计分析
采用Excel 2010和SPSS 21.0进行数据统计处理,用ANOVA单因素方差分析方法分析同一组分(或同一器官)生物量不同群落之间的差异显著性(P<0.05),用Bivariate相关分析方法分析样地生物量与样地树种多样性指数、土壤养分含量的相关性。
3 结果与分析
3.1 不同人为干扰程度森林群落生物量的空间分配格局
3.1.1 乔木层生物量的分配
从表4可以看出,随着人为干扰程度减弱,乔木层生物量增加,LAG比PLL提高了17.86%,其中LAG枝、干、根生物量比PLL分别提高了3.53%、20.57%和55.52%,而叶生物量下降了30.18%,地上部分生物量增加了12.54%,但无论是乔木层生物量还是同一组分生物量,两个群落之间的差异均不显著(P>0.05)。PLL、LAG乔木层各组分生物量的分配格局基本一致,均表现为:干最高,占乔木层生物量的59.13%—60.48%,其次是枝,占17.87%—20.34%,再次是根,占12.38%—16.34%,叶最小,仅占5.31%—8.15%;地上部分生物量占乔木层生物量的83.66%—87.62%,明显高于地下部分(根)。表明人为干扰程度减弱,有利于乔木层生长,特别有利于地下根系的生长,但对各组分生物量的空间分配格局影响不大。
3.1.2 灌木层生物量的分配
如表5所示,4个群落灌木层生物量、各组分生物量以及地上部分、地下部分生物量均呈现出随着人为干扰程度减弱,先增加后下降的变化趋势,LCQ均为最高,且与LVR、PLL、LAG差异显著(P<0.05),但LVR、PLL、LAG之间差异均不显著(P>0.05)。

表2 主要树种各组分生物量的相对生长方程
WL、WB、WS、WR分别表示树叶、树枝、树干、树根的生物量(kg/hm2),H表示树高(m),D表示胸径(cm)
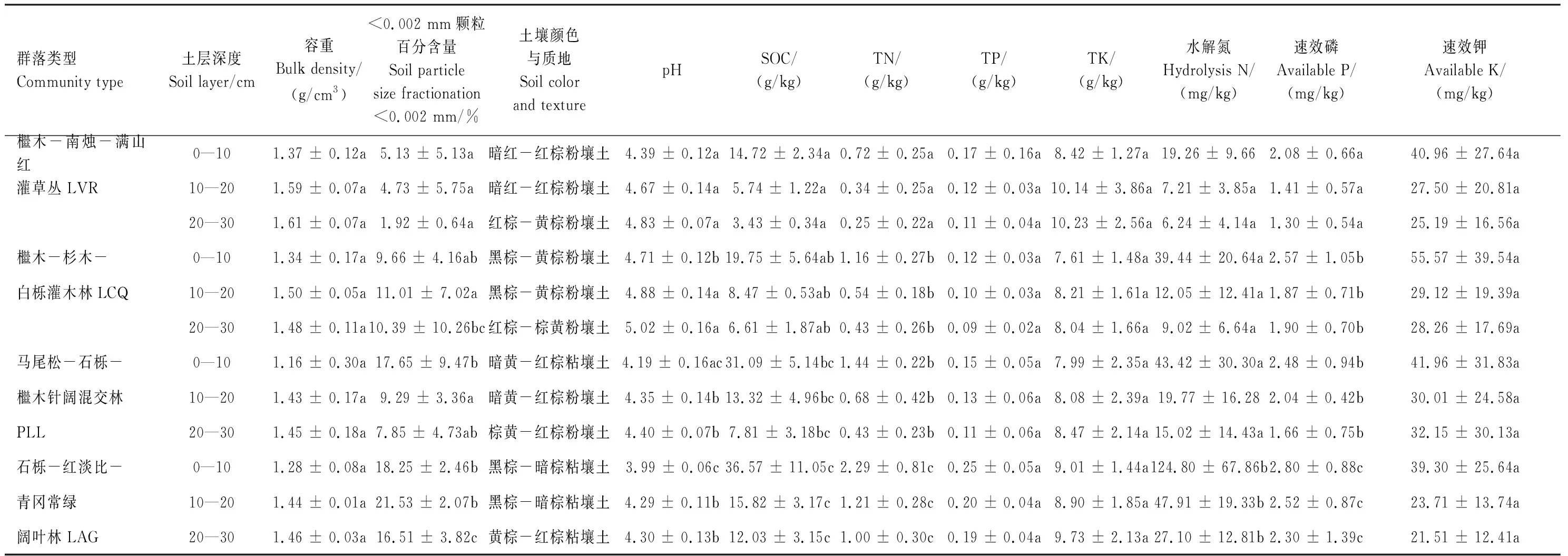
表3 不同森林群落土壤的理化性质
不同字母表示同一土层不同森林群落类型之间差异显著(P<0.05)

表4 乔木层生物量及其组分分配/(kg/hm2)
括号内的数据为百分率(%),不同字母表示同一组分不同森林群落之间差异显著(P<0.05)

表5 灌木层生物量及其组分分配/(kg/hm2)
不同干扰程度群落的灌木层各组分生物量分配格局不同。LVR根生物量最大,占其灌木层生物量的38.93%,其次是枝和干,分别占21.53%和20.43%,叶最小,仅占19.12%;LCQ各组分生物量依次为:干>根>枝>叶,分别占其灌木层生物量的38.63%、33.34%、16.74%、11.28%;PLL依次为:根>干>枝>叶,分别占其灌木层生物量的43.70%、27.47%、15.83%、13.00%;LAG枝(包括干)生物量最高,占其灌木层生物量的42.31%,其次是根,占36.75%,而叶最低。4个群落地上部分生物量显著高于地下部分(P<0.05),占其灌木层生物量56.25%—66.66%,LCQ群落地下部分生物量占其灌木层生物量的百分比最低,仅为33.34%(表5)。
从表6可以看出,LVR草本层生物量及其地上部分、地下部分生物量均为最高,其次是LCQ和LAG,而PLL最低,呈现出随着干扰程度减弱先下降再升高的趋势,但4个群落之间的差异均不显著(P>0.05)。LVR、PLL和LAG地下部分生物量高于地上部分,其中LVR地上部分与地下部分差异显著(P<0.05),而PLL、LAG差异不显著(P>0.05);LCQ地下部分低于地上部分,但差异也不显著(P>0.05)。

表6 草本层生物量及其组分分配/(kg/hm2)
3.1.4 地表凋落物层现存量
如表7所示,凋落物层现存量及各分解层凋落物现存量总体上随着干扰程度减弱而逐渐增加,其中LVR与LCQ、PLL、LAG差异显著(P<0.05),但LCQ、PLL、LAG之间差异不显著(P>0.05)。LCQ、PLL和LAG凋落物层现存量表现为:已分解层>半分解层>未分解层,而LVR则表现为:半分解层>已分解层>未分解层。未分解层凋落物现存量占其凋落物层现存量的百分比随着人为干扰程度减弱没有明显的变化,半分解层呈下降趋势,已分解层呈增大趋势。表明随着人为干扰程度减弱,未分解层凋落物向半分解层和已分解层迁移速率增大,有利于土壤养分的归还。
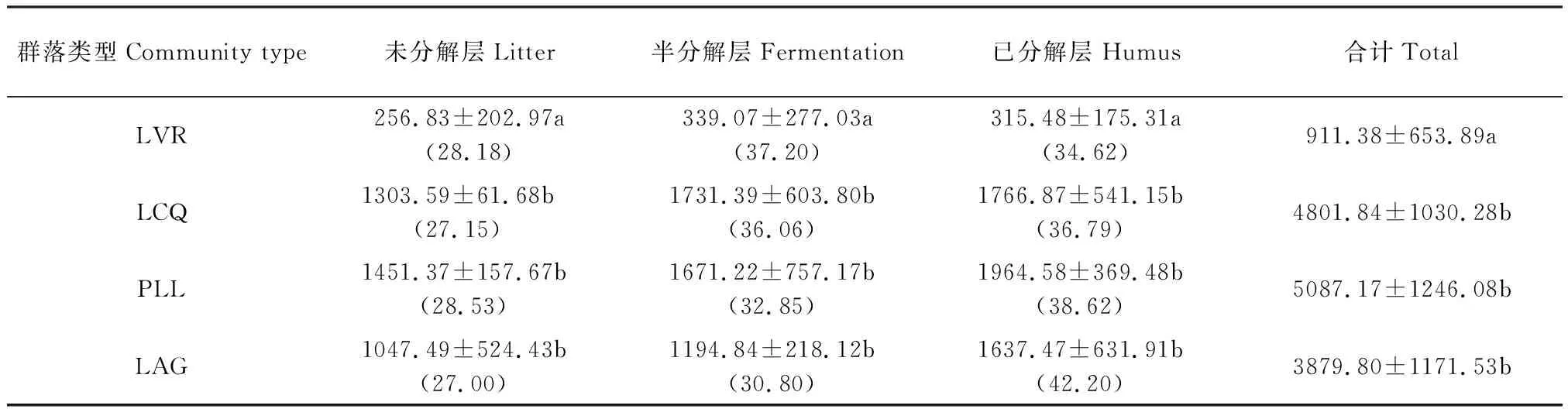
表7 地表凋落物层现存量/(kg/hm2)
3.1.5 群落生物量的空间分配
从表8可以看出,LAG群落生物量最高,分别是PLL、LCQ、LVR的1.15、7.23和28.73倍,且LAG、PLL与LCQ、LVR差异显著(P<0.05),LCQ与LVR差异显著(P<0.05),但LAG与PLL差异不显著(P>0.05),随着干扰程度减弱,群落生物量呈显著的指数函数增长(y=1927e1.191x,r=0.9594,P<0.05)。不同干扰程度群落生物量的空间分布格局不同。LVR灌木层生物量最高,占群落生物量的42.35%,草本层次之,占40.07%,凋落物层现存量最低。LCQ灌木层生物量最高,占群落生物量的69.98%,其次是地表凋落物层,占23.32%,草本层最低。PLL、LAG乔木层生物量最高,占群落生物量的93.74%—95.76%,占有绝对优势,其次是凋落物层,占2.60%—3.94%,再次是灌木层,占0.78%—1.89%,草本层最低,仅占0.43%—0.86%。与LVR相比,LCQ灌木层、凋落物层生物量显著增加(P<0.05),但草本层生物量明显下降;与LCQ相比,PLL、LAG乔木层生物量取代了灌木层的优势地位,而PLL、LAG灌木层生物量显著下降(P<0.05),草本层、凋落物层生物量变化不大(P>0.05);LAG乔木层生物量的优势比PLL更为明显。表明群落上层生物量是各群落总生物量的主要来源,且随着人为干扰程度减弱,群落上层生物量变化显著,优势逐渐增强,群落下层生物量变化较小,优势逐渐减弱。
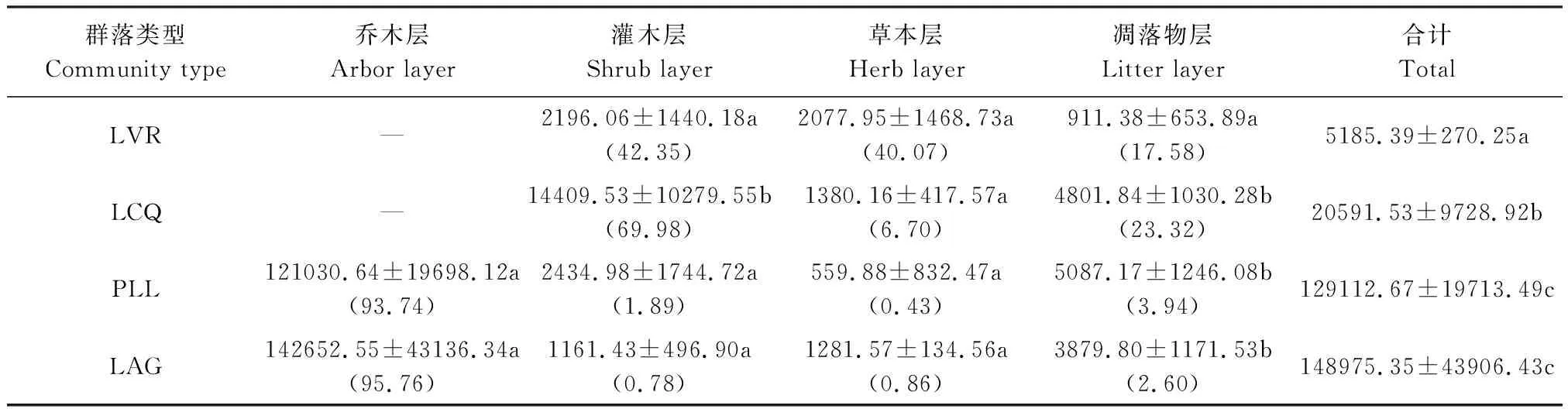
表8 不同干扰程度群落生物量的空间分配/(kg/hm2)
从图1可以看出,4个群落地上部分、地下部分生物量均随着人为干扰程度减弱而增加,从LVR到LAG,地上部分生物量增加了118450.7 kg/hm2,提高了5277.47%,地下部分增加了25339.26 kg/hm2,提高了861.61%。表明随着人为干扰程度减弱,群落地上部分、地下部分生物量表现为异速生长。LVR地下部分生物量比地上部分高31.03%,LCQ地上部分与地下部分相差不大,PLL和LAG地上部分分别比其地下部分高402.76%和326.78%。从LVR、LCQ到PLL、LAG,地上部分生物量占其群落生物量的百分比也呈现增加的趋势,而地下部分呈下降趋势。
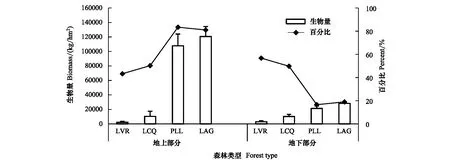
图1 不同干扰程度群落地上部分生物量和地下部分生物量的分配Fig.1 The distribution of aboveground biomass and underground biomass in different degrees of human disturbance communitiesLVR:Loropetalum chinense-Vaccinium bracteatum-Rhododendron mariesii scrub-grass-land,LCQ:Loropetalum chinense-Cunninghamia lanceolata-Quercus fabri shrubbery,PLL:Pinus massoniana-Lithocarpus glaber-Loropetalum chinense coniferous-broad leaved mixed forest,LAG:Lithocarpus glaber-Cleyera japonica-Cyclobalanopsis glauca evergreen broad-leaved forest
3.2 不同程度的人为干扰群落生物量与物种多样性的关系
从表9可以看出,群落总生物量与物种多样性指数呈显著正相关(相关系数为0.554,P<0.05)。地上部分、地下部分生物量分别与物种多样性指数呈弱显著的正相关(相关系数分别为0.551和0.550,P分别为0.051和0.052)。
3.3 不同程度的人为干扰群落生物量与土壤养分含量的相关性
如表9所示,群落总生物量、地上部分、地下部分生物量分别与0—30 cm土层有机碳、全氮、水解氮含量呈极显著正相关(P<0.01),其中,与土壤有机碳含量呈现出最强的正相关性(相关系数为0.875—0.912),与速效磷含量呈显著正相关(P<0.05),与全磷含量呈正相关,但不显著(P>0.05),与全钾、速效钾含量不存在显著相关性(P>0.05)。
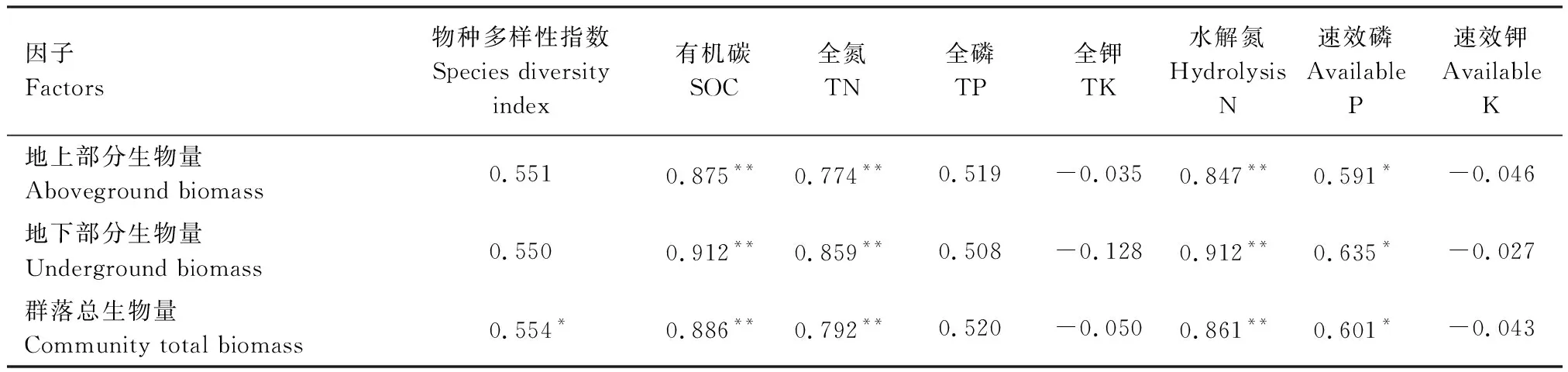
表9 群落生物量与物种多样性指数、土壤养分含量的相关系数
*P<0.05,**P<0.01,n=13
4 结论与讨论
4.1 不同程度的人为干扰对群落生物量的影响
群落生物量与植物多样性的关系一直是陆地生态系统的研究热点之一[43],但至今还没有一致的结论,两者之间的相互作用机制还不完全清楚[44]。研究表明,在人天混种群中,乔木树种混交比例不同,在忽略人为干扰历史(如抚育间伐)的情况下,随着林龄增加,乔木层树种增多,物种多样性指数增大,生物量显著增加,多样性与生物量基本呈单调的线性增长关系[44]。土壤养分与植物生长密切相关,土壤养分含量直接影响群落的生产力,土壤养分越充足,群落生产力越高[45-47]。本研究中,随着人为干扰程度减弱,群落生物量呈显著的指数函数增长(P<0.05)。群落生物量与群落样地树种多样性指数呈显著的正相关(P<0.05),与0—30 cm土壤层有机碳、全氮、水解氮、有效磷含量呈显著的正相关(P<0.05)。究其原因是由于,在LVR群落中采樵、火烧、放牧等人为干扰时有发生,不仅将林木带出林外,也使得灌木平均树高增加缓慢甚至下降。另一方面,从林地带走了大量的养分,不仅显著减少土壤轻腐殖质输入量,而且土壤表层大面积裸露,土壤养分流失严重,土壤肥力明显衰退,因而LVR群落以矮小的灌木、草本植物为主,个体数量虽多但生势差,群落生物量主要来源于灌木、草本植物,因此LVR群落生物量低。人为干扰程度相对减弱后,LCQ群落灌木层植物平均高明显增加,而且物种数明显增多,地表凋落物层现存量也明显增加,土壤碳、氮、磷含量明显提高,灌木层植物得到较好的发育和生长,因此LCQ群落生物量较显著增加。停止了采樵等人为干扰活动后,PLL、LAG群落优势树种由低矮的灌丛或灌木植物恢复为高大的乔木树种,更具有生长优势,生长速度加快,平均胸径和平均树高显著增加,林分树种多样性指数明显提高(表1),地表凋落物层现存量明显积累(表7),土壤养分(C、N、P)含量及其有效性明显提高(表3),更有利于林木的生长,因此PLL、LAG群落生物量显著增加。表明不同程度的人为干扰造成群落物种多样性、土壤养分(C、N、P)含量变化,是导致群落生物量变化的主要因素。群落生物量与0—30 cm土壤层有机碳含量之间存在着最高的相关性,是由于土壤有机碳是生态系统生产力和更新中最重要的一部分,是植被与土壤恢复的关键因素[48]。因此减弱或停止人为干扰,促进灌草丛群落变为正向演替,提高群落物种多样性,提高土壤养分含量,改善土壤养分循环,以形成林木生长与土壤改善相互促进的状态。
研究表明,群落凋落物层现存量主要取决于群落凋落物量及其分解速率两个因素[49]。本研究中,凋落物层现存量总体上随着人为干扰程度减弱而逐渐增加,是由于随着人为干扰程度减弱,物种种类增加,大型树木比例增加(表1),群落中物种的新陈代谢加快,凋落物量增加[50];也是由于PLL林下凋落物以针叶为主,较难腐烂分解,而LAG林下凋落物以阔叶树叶为主,较易于分解,因此PLL凋落物层现存量高于LAG。
4.2 不同程度人为干扰对群落生物量空间分布格局的影响
群落生物量在不同层次上的分配受到群落特性和物种组成等因素的综合影响[43]。影响群落生物量的因素较多,包括光合、呼吸、脱落、病虫害消耗等,其中光合作用是一个重要因素,在不同群落中,相同层次的生物量与光照强度密切相关,人为干扰会造成群落光照强度的差异[51]。本研究中,不同干扰程度群落生物量的空间分布格局不同。可能是由于LVR群落遭受采樵、火烧、放牧等严重人为干扰后,以矮小的灌木、草本植物为主,没有形成明显的灌木层,凋落物不能有效积累,因而灌木层、草本层生物量相当,凋落物层现存量最低;随着人为干扰程度减弱,没有形成乔木层的LCQ群落,灌木层植物在全光照条件下能充分发育和生长,灌木平均高明显增加,形成明显的灌木层,生物量显著增加,林下光照强度减弱,不利于草本层植物生长,生物量明显下降,另一方面,在调查过程中发现,LCQ群落落叶灌木树种较多,大量枯枝落叶的产生使得林下地表凋落物层现存量较高,因而群落生物量呈现出灌木层>凋落物层>草本层的分布格局,与湖南浏阳大围山自然保护区杜鹃灌丛生物量不同层次间的分布格局一致[1];停止了采樵等人为干扰活动后,PLL、LAG形成乔木层,更具有生长优势,代谢更为强烈,凋落物量明显增加,上层郁闭度增加,林下光照强度减弱,不利于灌木层、草本层植物生长,生物量下降,从而呈现出乔木层生物量占有绝对的优势,其次是凋落物层,灌木层、草本层最低。表明不同程度的人为干扰下,群落植物组成和群落结构不同,群落生物量的空间分配格局不同。
在不同环境条件下,群落生物量的分配格局反映了植物对环境的响应规律和资源分配策略[3]。最优分配理论认为,植物通过调节各器官生物量的分配来适应外界环境,从而最大限度地获取有限资源,以维持其最大生长速率[52]。当光照受到限制时,植物会将资源更多地分配到茎和叶等地上部分,而当水分和养分受到限制时,植物会将更多的资源分配到根系中[53]。本研究中,LVR群落由于人为干扰严重,造成地上部分生物量不能有效积累,而树蔸及根部残留在林地里,生物量没有明显变化,因此无论群落生物量还是灌木层、草本层地下部分生物量高于地上部分。在LCQ群落,没有形成乔木层,灌木层植物在全光照条件下,有利于地上部分的生长,生物量明显提高,占其灌木层生物量的比例增高,地上部分生物量与地下部分相差不大。人为干扰减弱或停止后,PLL、LAG群落形成乔木层,物种组成和群落结构逐渐趋于稳定,凋落物量显著增加,土壤水分和养分得到明显改善,有利于植物的生长和发育,地上部分、地下部分均有明显生长,生物量明显增加,但由于大型乔木树种增多(表1),对光照的竞争更为激烈,乔木层植物将更多的生物量分配到地上部分,增强对光照的竞争能力,以获得更大的生长空间和更多的资源,因而地上部分生物量增长速度明显高于地下部分,从而表现为异速生长,也体现了PLL、LAG群落对亚热带地区温暖湿润环境条件的适应[1,54-55]。
