高地钩叶藤和大钩叶藤维管束与导管变异研究
2018-10-12张菲菲汪佑宏季必超张利萍刘亚梅
张菲菲,李 担,汪佑宏,季必超,薛 夏,张利萍,刘亚梅
棕榈藤(Rattan)属棕榈科(Palmae或Arecace- ae)省藤亚科(Calamoideae)省藤族(Calameae),天然分布于东半球的热带地区及邻近地区。其明显的特征是攀援,茎外部常常包被有刺叶鞘。
棕榈藤材是可再生非木材林产品之一,具有很高的经济价值和使用价值,仅次于木材和竹材。其藤茎素有“藤条”之称,外观像竹子,茎为实心。在相同的立地条件下,同种藤的直径变异很小,但节间长度变异很大[1]。藤茎富含纤维,质地柔韧,抗拉、抗弯强度大,是编织藤质家具和工艺品的重要原料,占据很大的市场[2-4]。
高地钩叶藤(Plectocomia himalayana)为棕榈科省藤亚科钩叶藤属,属于攀援、丛生的藤类。在我国主要分布于云南一些海拔在1 450~1 800 m的箐沟、竹林、山地常绿阔叶林中;在国外主要分布在不丹、印度、尼泊尔、老挝、泰国。其藤茎质地较粗糙,一般用于编织较粗糙的藤器或栅栏用[5-6]。目前,有关高地钩叶藤的材性的研究报道很少[7],其物理力学性能、化学成分和纤维形态等材性方面均需要研究。
大钩叶藤(Plectocomia assamica)属于钩叶藤属,藤茎粗大(9~14 cm),我国分布于云南南部和东南部局部地区,国外仅在勐腊马关境内海拔820~1 230 m的次生林中以及印度、缅甸北部有分布[8-9]。其藤茎质脆,目前很少利用。
目前对高地钩叶藤和大钩叶藤的利用之所以很少,主要是由于对其性质和结构的了解不够。棕榈藤的功能主要取决于维管束及其周围的维管束鞘[10]。维管束散生于基本薄壁组织中,其分布密度对藤茎的坚韧程度影响很大,导管孔径的大小和形状还对水分的流动性有着显著的影响,同时导管孔径的大小也影响着藤材的韧性,是选择优良品种的主要依据[11-13]。有学者对高地钩叶藤和大钩叶藤的组织比量、力学性能的研究,尚莉莉[14]将其与玛瑙省藤材进行比较表明,大钩叶藤材的材性较差,直接应用不能满足商业化利用的要求,需对其进行改性研究,本研究针对高地钩叶藤和大钩叶藤维管束与导管,为其材性和改性的研究提供理论基础。
1 材料与方法
1.1 材料
试验所用大钩叶藤采自云南盈江县,高地钩叶藤采自云南省梁河县。齐根砍伐3株生长健康的野生高地钩叶藤、大钩叶藤,并剥去外层带刺的藤鞘。整藤气干后分别在藤茎的基部、2 m处、中部和梢部截取若干个试块备用(试块长1~2 cm)。
试验仪器:101-1EBS型电热鼓风干燥箱,上海科恒实业发展有限公司;DZF-1B型真空干燥箱,上海科恒实业发展有限公司;KD-1508A型切片机,浙江省金华市科迪仪器设备有限公司。
1.2 方法
试样进行密度分析后,进行微波软化,然后把软化的材料进行聚乙二醇(Polyethylene glycol)包埋。将冷却的包埋块表面多余的聚乙二醇固体削除后用旋转式切片机切取厚度为35 μm左右的横切面切片。使用Leica CW4000自动成像系统测量导管的直径以及维管束的径向和弦向直径,并采用计点法(网格法)测量导管与维管束的密度[15-16]。每个指标统计30组后计算相应密度与直径的平均值。
2 结果与分析
2.1 维管束解剖特征径向变异结果分析
高地钩叶藤和大钩叶藤维管束解剖特征径向分布见图1。可以看出,藤皮、藤中到藤芯,高地钩叶藤维管束密度分别为 5.586、1.776 个·mm-2和1.243个·mm-2,大钩叶藤的维管束密度分别为4.819、1.951 个·mm-2和 1.208 个·mm-2,均呈现逐渐降低的变化趋势,这与对黄藤的研究结果一致[17]。高地钩叶藤和大钩叶藤维管束密度均值分别为2.87个·mm-2和 2.66 个·mm-2,前者比后者高 7.9%,经 F检验,在0.05水平上差异不显著(表1)。
藤皮、藤中到藤芯,高地钩叶藤维管束径向直径分别为 466.208、678.742 μm 和 664.815 μm,呈先增大后减小的变化趋势;大钩叶藤维管束径向直径分别为 541.414、665.954 μm 和 715.111 μm,呈现递增的趋势。其中高地钩叶藤和大钩叶藤维管束径向直径均值分别为 603.23 μm 和 640.83 μm,前者比后者低6.2%,经F检验,在0.05水平上差异不显著。
藤皮、藤中到藤芯,高地钩叶藤维管束弦向直径分别为 355.587、501.291 μm 和 497.925 μm,呈先增后减的变化趋势,大钩叶藤维管束弦向直径分别为433.888、510.251 μm 和 542.916 μm,呈现递增趋势。高地钩叶藤和大钩叶藤维管束弦向直径平均值分别为 451.60 μm 和 495.69 μm,前者比后者低 9.8%,经F检验,在0.05水平上差异不显著。
2.2 导管解剖特征径向变异结果分析
高地钩叶藤和大钩叶藤导管解剖特征径向分布见图2。可以看出,藤皮、藤中到藤芯,高地钩叶藤导管密度分别为 7.356、1.868 个·mm-2和 1.636 个·mm-2,大钩叶藤导管密度分别为 5.268、1.945 个·mm-2和 1.237 个·mm-2,均呈现逐渐减小的变化趋势。高地钩叶藤和大钩叶藤导管密度均值分别为3.62 个·mm-2和 2.82 个·mm-2,前者比后者高 28.5%,经F检验在0.05水平上差异显著(表1)。
藤皮、藤中到藤芯,高地钩叶藤导管直径分别为145.942、260.196 μm 和 250.845 μm,大钩叶藤导管直径分别为 184.713、292.596 μm 和310.063 μm,高地钩叶藤呈现先增大后减小的变化趋势,大钩叶藤呈现递增的变化趋势。高地钩叶藤和大钩叶藤导管直径均值分别为 219.00 μm 和 262.46 μm,前者比后者低19.8%,经F检验在0.05水平上差异不显著(表1)。
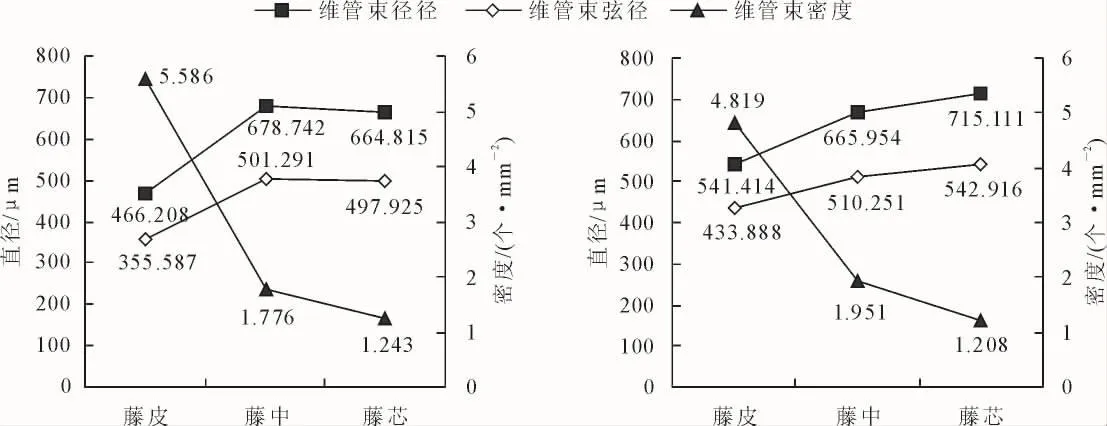
图1 高地钩叶藤(左)与大钩叶藤(右)维管束径向分布Fig.1 The radical variation of vascular bundle of P.himalayana(left)and P.assamica(right)
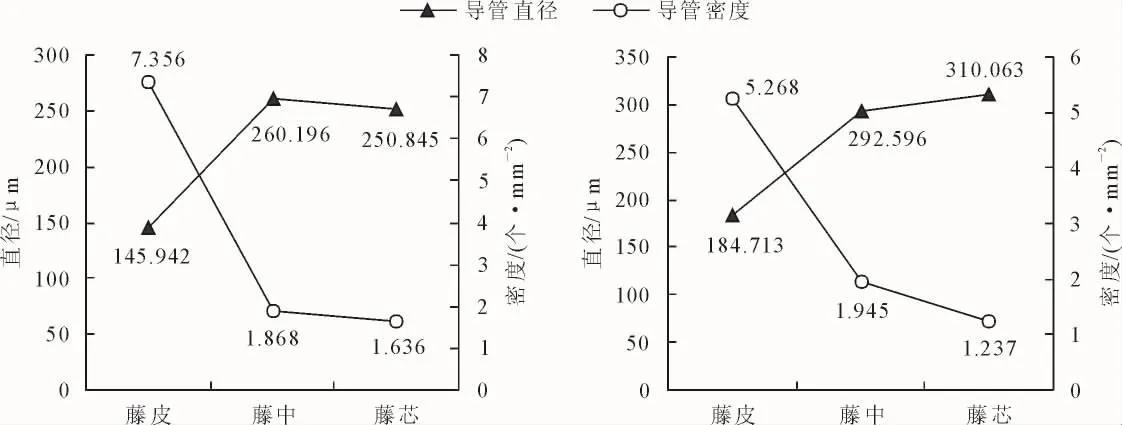
图2 高地钩叶藤(左)与大钩叶藤(右)导管径向分布Fig.2 The radical variation of vessel of P.himalayana(left)and P.assamica(right)
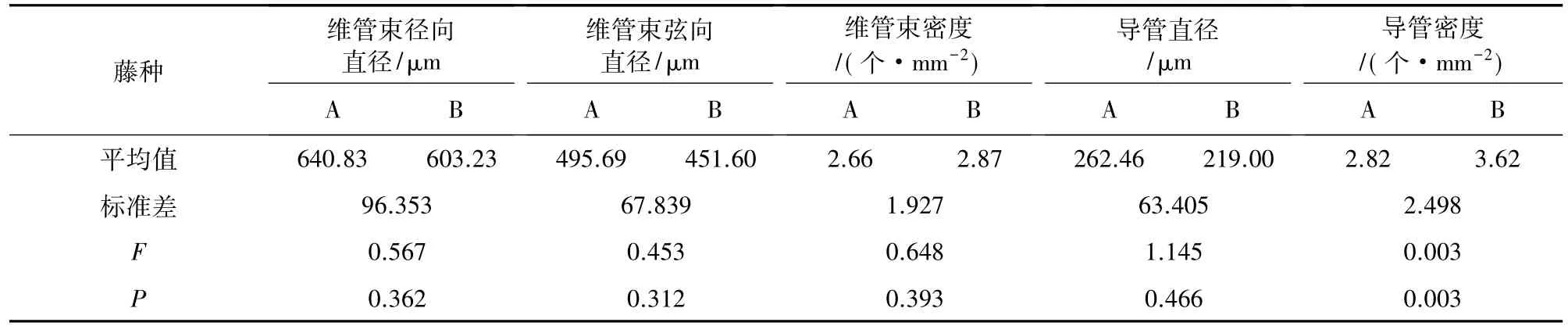
表1 大钩叶藤和高地钩叶藤维管束、导管径向变异及F检验Table 1 The radical variation of vascular bundle and ducts and F-test of P.assamica and P.himalayana
2.3 维管束与导管直径轴向变异结果分析
高地钩叶藤和大钩叶藤维管束直径与导管直径轴向分布见图3。可以看出,高地钩叶藤维管束径向直径为梢部>基部>2 m处>中部,随着轴向高度的增加,在 591.741~629.405 μm 的变化范围内呈现先减小后增大的变化趋势;大钩叶藤的维管束径向直径为中部>梢部>2 m处>基部,呈先增大后减小的变化趋势,变化范围在 633.520~710.699 μm。F检验在0.05水平上差异不显著(表2)。
高地钩叶藤维管束弦向直径为梢部>2 m处>基部>中部,呈先减小后增大的变化趋势,变化范围425.662~457.337 μm;大钩叶藤维管束弦向直径为2 m处>基部>梢部>中部,呈先增大后减小的变化趋势,变化范围在 485.723~536.415 μm。经 F 检验在0.05水平上差异显著(表2)。
高地钩叶藤导管直径为梢部>2 m处>中部>基部,呈逐步递增的变化趋势,变化范围在198.170~241.950 μm;大钩叶藤导管直径为中部>基部>梢部>2 m处,呈先增大后减小的变化趋势,变化范围在262.457~304.975 μm。经 F 检验在 0.05 水平上差异不显著(表2)。
2.4 维管束与导管密度轴向变异结果分析
高地钩叶藤和大钩叶藤维管束与导管密度轴向分布见图4。可以看出,高地钩叶藤维管束密度为基部>中部>2 m 处>梢部,呈“降-增-降”起伏式变化,变化范围在 2.592~3.235 个·mm-2;大钩叶藤维管束密度为基部>2 m处>梢部>中部,呈先减小后增大的变化趋势,2.093~2.725 个·mm-2。经 F 检验在0.01水平上差异极显著(表2)。
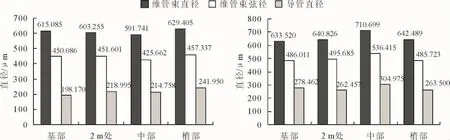
图3 高地钩叶藤(左)与大钩叶藤(右)维管束和导管尺寸轴向分布Fig.3 The axial variation of vascular bundle radial and duct diameter of P.himalayana(left)and P.assamica(right)
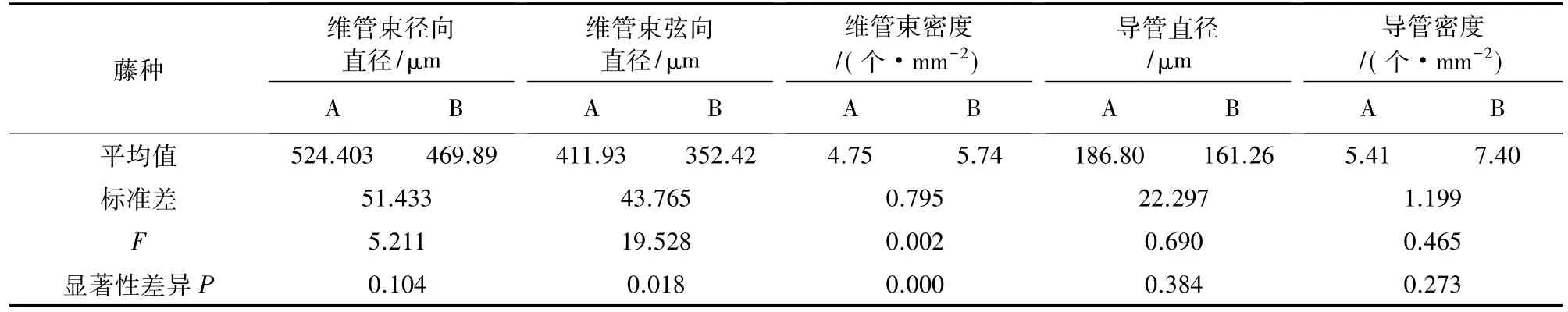
表2 大钩叶藤和高地钩叶藤维管束、导管轴向变异及F检验Table 2 The axial variation of vascular bundle and ducts and F-test of P.assamica and P.himalayana
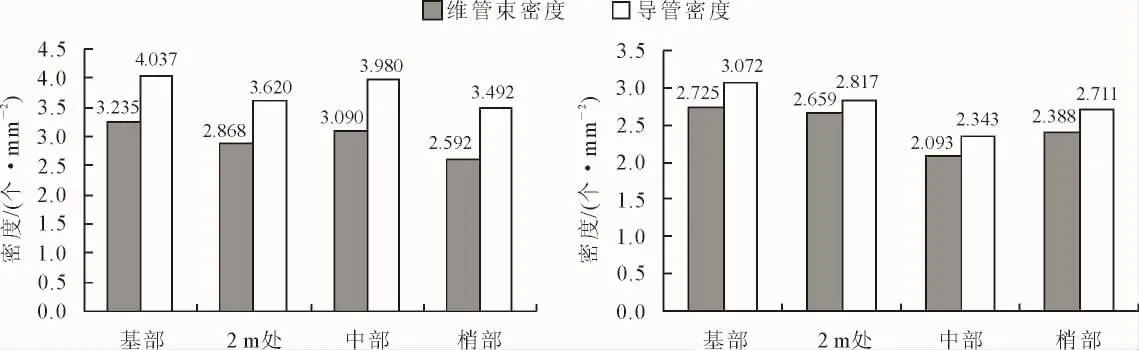
图4 高地钩叶藤(左)与大钩叶藤(右)维管束和导管密度轴向分布Fig.4 The axial variation of vascular bundle radial and duct density of P.himalayana(left)and P.assamica(right)
高地钩叶藤导管密度为基部>中部>2 m处>梢部,与维管束密度的变化规律相同,也呈“降-增-降”起伏式的变化,3.492~4.037 个·mm-2;大钩叶藤导管密度为基部>2 m处>梢部>中部,也与维管束密度的变化规律相同,呈现先减小后增大的变化趋势,2.343~3.072 个·mm-2。经 F 检验在 0.05 水平上差异不显著(表2)。
3 结论与讨论
从藤皮至藤芯2种藤材维管束与导管特征的径向变化中,高地钩叶藤和大钩叶藤的维管束径向直径、维管束弦向直径、导管直径、维管束密度和导管密度呈现不同的变化趋势;经F检验,2种藤材导管的密度在0.01水平上差异极显著。高地钩叶藤和大钩叶藤间维管束与导管特征的这种差异,除与藤种相关外,还与其在径向的位置等存在必然的联系;因此在选择藤种加以利用时,可根据其构造特征差异(如维管束密度大,其力学强度往往也较大)来进行;此外,生产上也常常把藤茎分成藤皮、藤芯进行加工利用。
随着藤材轴向高度的升高(即随着藤龄的减小),高地钩叶藤、大钩叶藤维管束径向直径、弦向直径、导管直径、维管束密度和导管密度呈现出不同的变化趋势。其最大值、最小值先后出现在梢部、中部、2 m处及基部,经F检验,2种藤材维管束弦向直径在0.05水平上差异显著,维管束密度在0.01水平上差异极显著。造成这种结果的原因,主要是取材的部位不同,藤材对应的藤龄也不同造成的;一般藤龄大,其材性也较好,因此,生产上一般都去除藤茎的梢部、不进行加工利用的。
