陕西凤翔秦公一号大墓车马坑马骨遗骸古DNA研究*
2018-09-05蔡大伟朱司祺胡松梅田亚岐孙洋陈曦周慧
蔡大伟 朱司祺 胡松梅 田亚岐 孙洋 陈曦 周慧
(1.吉林大学边疆考古研究中心;2.陕西省考古研究院;3.山东省石刻艺术博物馆;4.吉林市博物馆)
马的驯化是欧亚草原游牧文明兴起、繁荣的关键,深刻地影响着人类社会的发展进程。在中国,家马和马车是商代晚期突然大量出现的,然而晚商之前有关马的考古材料非常少。早期驯化阶段的缺失和商代晚期家马的“突然”出现,使中国家马的起源引起了广泛的争论。一些学者认为中国家马是从中亚草原地带传入的,甘青地区和北方草原地带可能是其进入中国的主要通道[1,2]。也有学者认为中国家马是由本土的普氏野马驯化而来[3,4],在驯化时间上存在“新石器时代中期”[5]、“龙山时期”[6]、“夏末商初”[7]等看法。近年来的古DNA研究显示中国古代马既有本地驯化的因素,也受到外来家马基因流的影响,欧亚草原很可能是家马及驯化技术由西向东传播进入中国的一个主要路线。但是,由于缺乏西北地区家马的DNA数据,家马在中国的扩散路线尚不清楚。
秦公一号大墓位于凤翔县城南雍水河北岸平地上,于1976~1986年先后发掘十年。大墓规模庞大,远远超过先秦诸侯的墓葬,是中国发掘的最大的先秦墓葬。根据墓葬中出土的石磬上的铭文,考古学家推断秦公一号大墓的墓主人为秦景公(?~前537年)。墓葬展
示了春秋晚期秦国的强大,随葬物品丰富,墓葬规格很高,出土的黄肠题凑是周秦时代规格最高的葬具,并有183人的人殉[8]。2007年,陕西省考古研究院人员在秦公一号大墓西南侧发现车马坑,坑为东西向长方形竖穴,东西长17.5、南北宽3.1米,坑内底部自东向西依次摆设了5组车马,考古人员根据其有车无轮的情况,推断其为正式陪葬坑之外的一个祭祀坑。这批车马坑的出土,一方面对于我们了解先秦车制、车马殉葬制度及祭祀制度诸多问题具有重要意义;另一方面,殉葬的马匹对我们探索中国家马的起源具有重要意义。本研究希望利用古DNA技术重建古马的遗传结构,通过对比分析,揭示陕西地区古代马与其他地区古代马之间的遗传关系,为研究家马在中国的扩散提供有价值的线索。
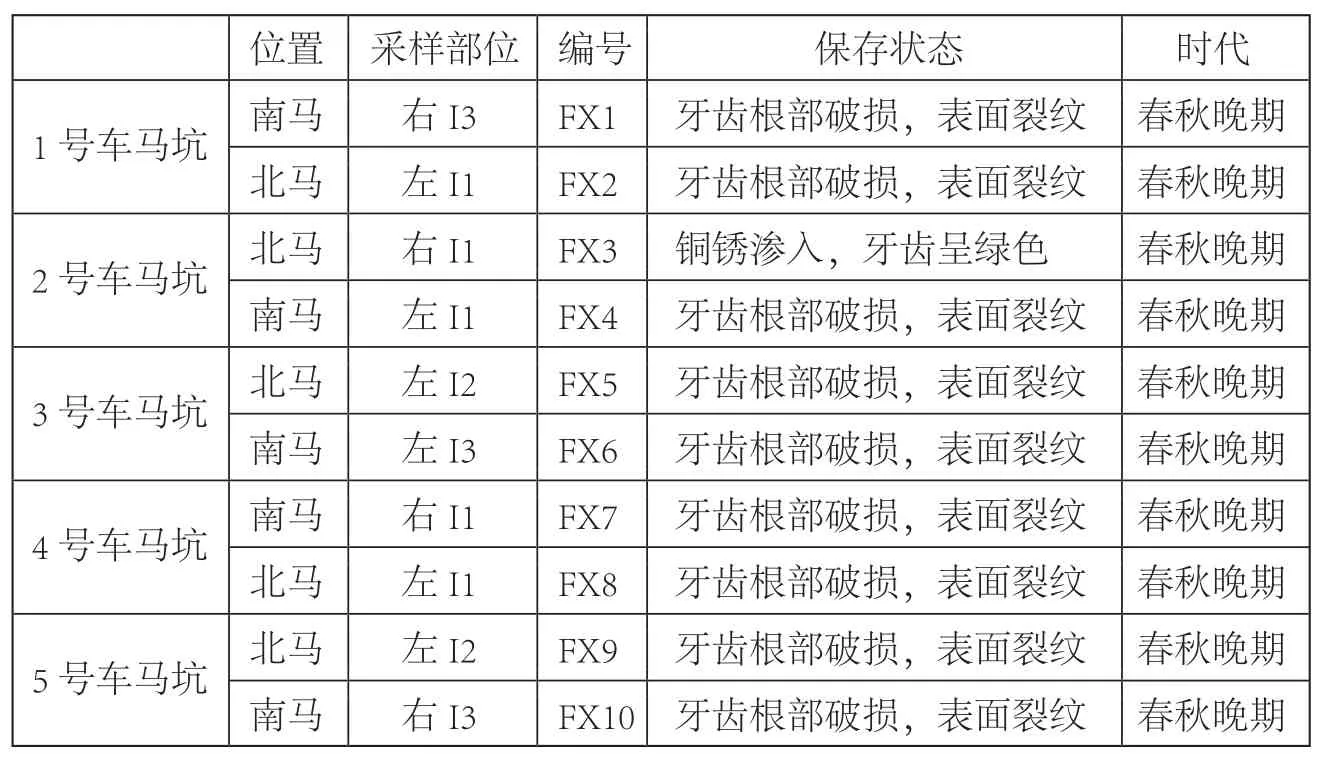
表一 秦公一号大墓取样及样本保存情况
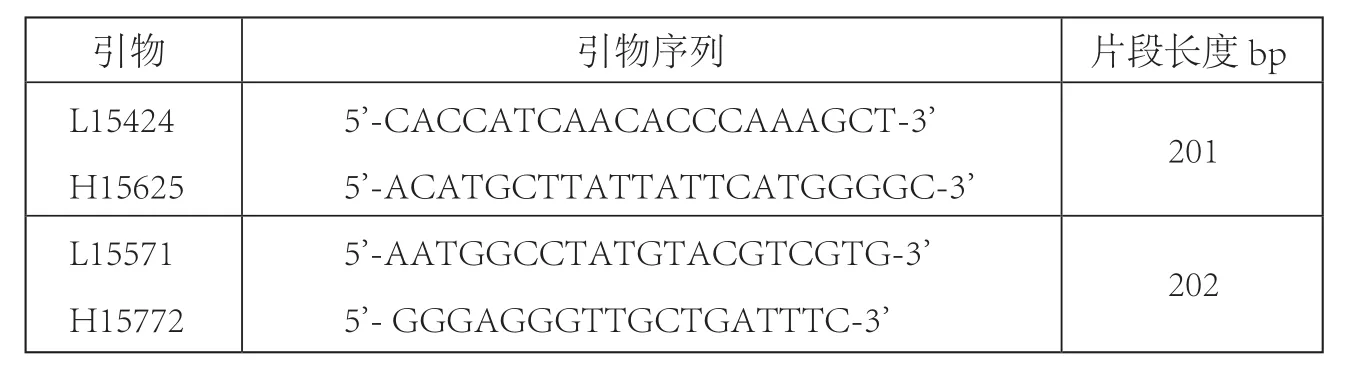
表二 本研究所用扩增引物
一、材料和方法
1. 样本来源及保存状况
本研究样品来自2007年7月至11月发掘的陕西凤翔秦公一号大墓1号祭祀坑。坑底东西向依次摆设5组车(自东向西依次编号1~5号车),每组车前分别有挽马工具。我们对每匹马的门齿进行了采样,进行古DNA分析(表一)。
2. 古代样本处理
首先用毛刷除去样本表面污垢,接着用电动打磨工具去除表层1~2 mm,然后用10%次氯酸溶液浸泡古代材料5~10min,在紫外线照射下晾干,随后将样品放入液氮冷冻粉碎 机 FREEZER/MILL6750(SPEXP CetriPrep,USA)之中,液氮冷却,打磨成粉,-20℃冷冻保存。
3. 古DNA提取、扩增和测序
古DNA提取参照杨东亚等人的文献进行[9]。
PCR扩增:依据马的参考序列X79547[10]设计两对套叠引物,扩增349bp (核苷酸位置15424—15772) 线粒体DNA片段(表二)。扩 增 反 应 均 在 Mastercycler® personal 热 循 环仪 (Eppendorf,Germany) 上 进 行。50 μL反应体系中含 2.5 mM Mg2+,1X Buffer,0.2 mMdNTPs, 0.5 μM 正反引物,1U Taq 酶,2 μL模版。程序如下:95℃ 5 min,92℃ 1 min变性,50-55℃ 退火 1 min,72℃ 延伸 1 min,36个循环后,72℃ 延伸10 min,4℃保持。
测序:PCR扩增产物通过2% 琼脂糖(Biowest,German)凝胶电泳检测,使用QIAEX®Ⅱ GEL Extraction Kit 凝胶回收试剂盒(QIAGen,Germany) 纯化PCR产物。纯化产物用ABI PRISM® 310全 自 动 遗 传 分 析 仪(Applied Biosystems,USA)通过Dyeprimer试剂盒进行正反双向直接测序。
4. 污染的防止
所有实验都是在专门的古DNA实验室中完成。PCR前和PCR后实验操作分别在两个相距很远的、独立的实验楼中进行。每次实验前,实验室都要经过紫外线长时间照射,对超净台进行紫外线照射和通风至少30min,并随时用DNA 去除剂擦拭。进行实验时,实验人员穿着双层防护服,戴一次性帽子、口罩、双层手套,使用高压灭菌的一次性实验耗材。通过设置DNA抽提和PCR扩增空白对照,来判断实验步骤和实验试剂是否有污染。最后,对于同一个实验样本,不同实验人员要进行多次DNA抽提和PCR扩增重复操作,确保实验结果的准确性。
二、结果和分析
1. 秦公一号墓古代马DNA序列和世系归属
在秦公一号大墓出土的10个样本中,FX3、FX8、FX10未能成功提取DNA,其余7个样本均成功提取DNA并获得可靠的古DNA序列。其中FX4、FX5 成功获得349bp序列,FX1、FX2、FX6、FX7、FX9成功获得201bp序列(表三)。与参考序列X79547相比共检测出22个变异位点,所有碱基变异位点全部是转换,无颠换,无插入发生(表四)。
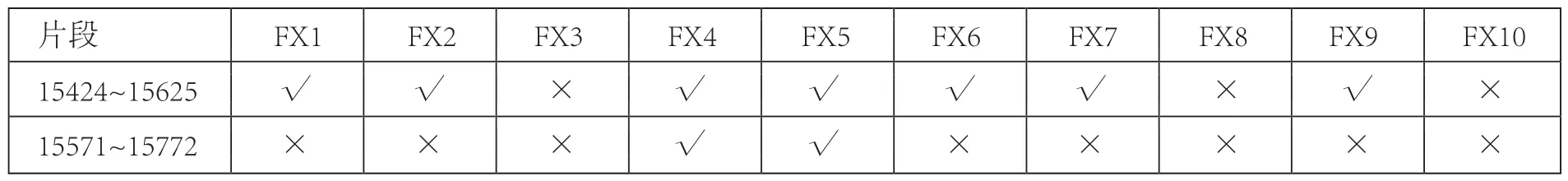
表三 秦公一号墓古代马实验结果
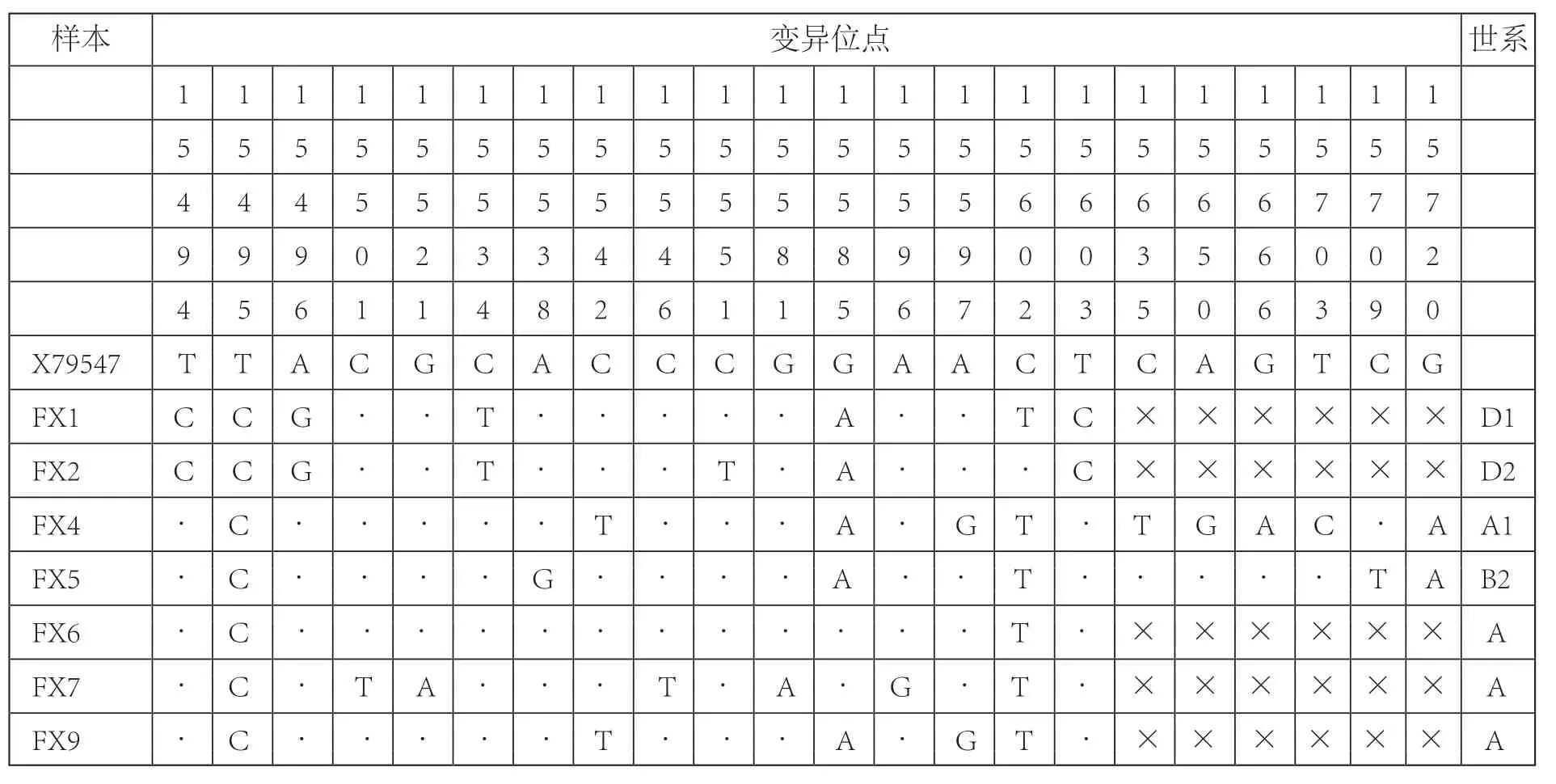
表四 秦公一号墓古代马DNA序列和世系归属
线粒体DNA被广泛应用于家马的起源研究中, Jansen[11]等将现代家马划分为7个线粒体DNA世系A—G,其中几个较大的世系可以进一步划分为多个亚组,世系A共有7个亚组A1—A7,世系B和C各有2个亚组B1—B2和C1—C2,世系F和D各有3个亚组F1—F3和D1—D3。根据变异位点分析,FX1和FX2分别属于D1和D2亚组,FX4属于A1亚组,FX5属于B2亚组(表四)。由于缺乏后段的信息,推测FX6、FX7、FX9属于A世系,具体的亚组信息需要进一步的实验来补全序列验证。
2. 秦公一号大墓古代马与现代马的遗传关系
为了追踪秦公一号大墓古代马与现代家马的遗传关系,我们利用BLAST程序在美国国家生物技术信息中心(NCBI) 的DNA序列数据库(GenBank)中进行共享序列搜索,即搜寻与这些古代序列完全匹配的共享序列。FX1、FX4、FX5、FX6、FX9在GenBank中搜索到了完全相同的共享序列,而FX2和FX7没有搜索到完全一致的共享序列。图一显示了古代马的共享序列在世界各地的分布情况,FX1主要分布在欧洲,尤其是伊比利亚半岛,在东亚等地有少量分布。FX4主要分布在西南西伯利亚、中亚和东亚。FX5所共享的单倍型数量较少,主要分布在中东、中亚、北亚和东亚。FX6在欧亚大陆分布最为广泛,在欧洲、阿拉伯半岛、中亚、北亚和东亚均有分布,表明FX6是现代家马的主要基因型。FX9主要分布在中亚、北亚和东亚地区。
3. 秦公一号大墓古代马序列在中国古代遗址中的分布情况
为了探索秦公一号墓古代马与其他地区古代马的关系,我们调查了秦公一号墓古代马在中国13个古代遗址[12~17]中的分布情况(图二、表五)。基于考古发掘资料分析,这些遗址的年代不尽相同。首先,大山前遗址的时代最早,属于中国北方青铜时代早期夏家店下层文化(年代距今2000~1500 BC)。其次是西周时期(年代距今1046~771 BC)的陕西少陵原遗址和枣树沟脑遗址,以及青海卡约文化(年代距今1000~800 BC)丰台遗址。最后,所有其他的遗址都属于春秋战国时期,年代距今500 BC。FX1、FX2、FX7在其他古代遗址中均无发现,为秦公一号大墓古代马所独有的单倍型。FX4、FX5、FX6和FX9在其他古代遗址中有分布,其中FX6序列分布最广泛,在丰台遗址、忻州窑子墓地、小双古城墓地以及毛园民宅均有分布;其次是FX4和FX9在枣树沟脑、忻州窑子墓地、新店子墓地均有发现;而FX5则仅出现在枣树沟脑遗址和于家庄墓地中。从时间上看,秦公一号大墓部分古代马在西周时期就已经在西北地区,并在春秋战国之际出现在宁夏、内蒙古地区,这一结果一方面反映了古代马的基因连续性,另一方面反映了不同地区的马群存在广泛的交流,可能与地区间的人群文化贸易交流活动有关。
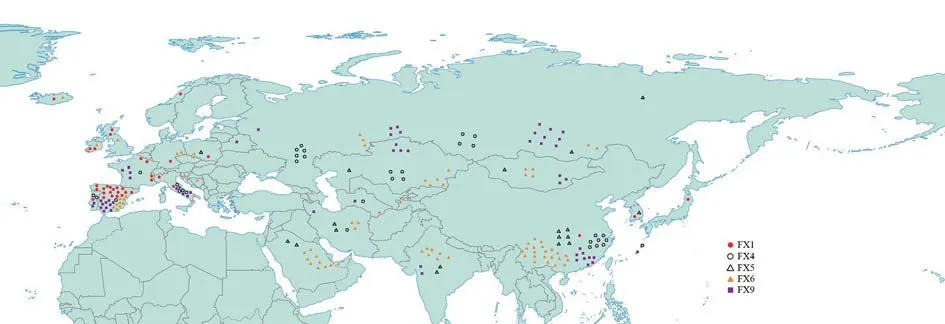
图一 秦公一号大墓样本共享序列在世界分布图
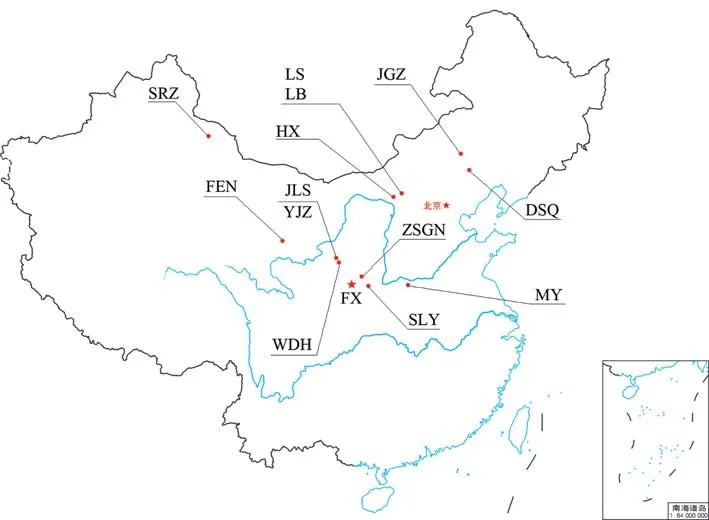
图二 本研究涉及的考古遗址的地理位置分布图
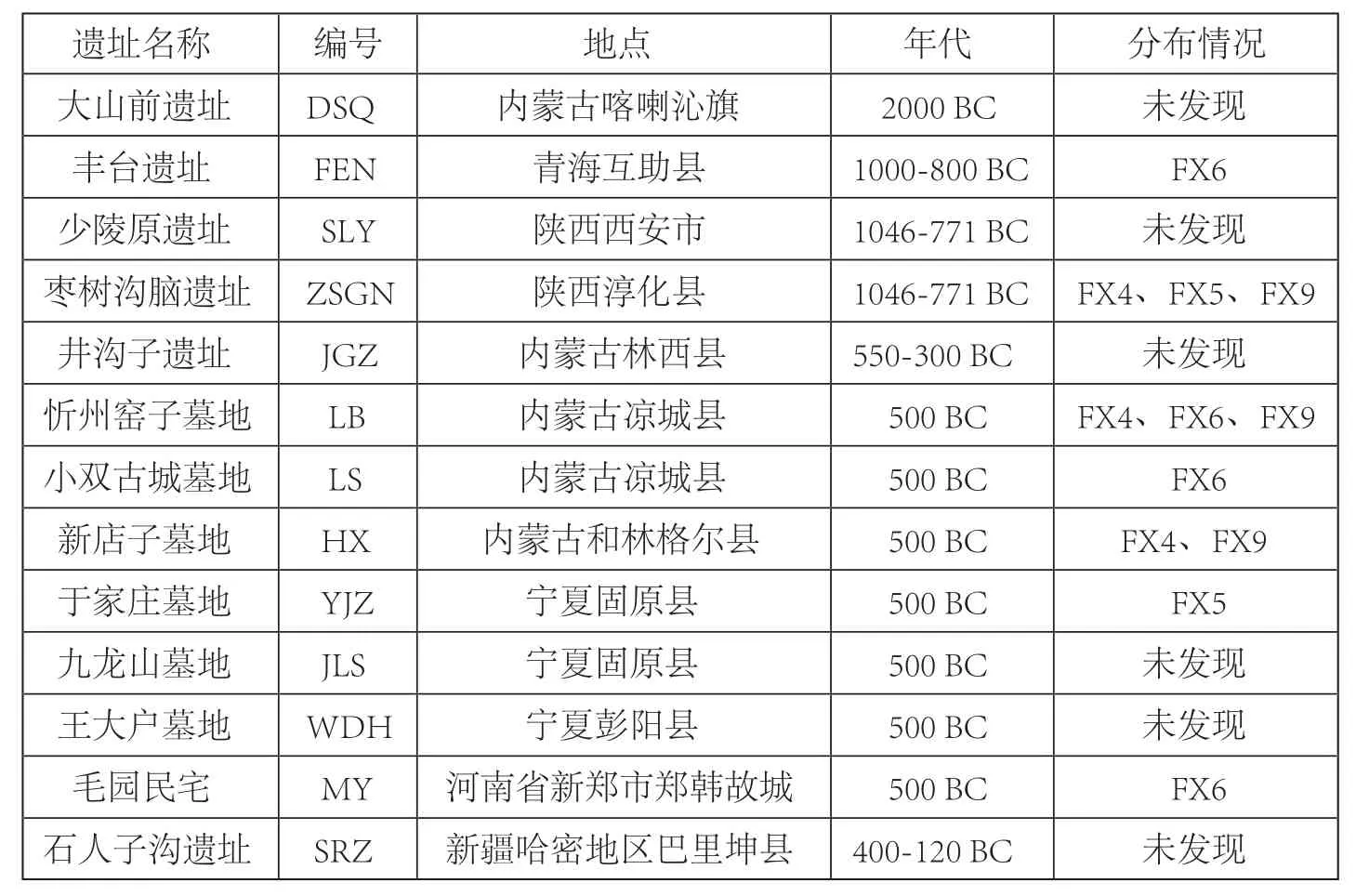
表五 秦公一号墓古代马在13个古代遗址中的分布
三、讨论
1. 秦公一号大墓古代马的母系来源
考古和基因证据表明,家马是在距今5500年前在中亚草原被驯化的[18,19],但是家马和马车从商代晚期才开始在中国大量出现,而且从马车的构造看,中国马车与西方马车在形制上基本相同,很可能与驯化的家马同时从西方传入[20],连接东西方的欧亚草原通道和经由新疆及河西走廊的绿洲之路是家马进入中国两条主要传播路线。在所获得的7个古代马DNA序列中,FX1属于D1亚组,而FX2属于D2亚组。世系D1亚组的建立者最早出现在南西伯利亚阿勒泰地区(900BC)[15],稍晚的时候在内蒙古凉城县忻州窑子墓地(500BC)[14]和哈萨克斯坦Berel遗址(300BC)[21]出现。世系D2亚组的建立者最早出现在青铜时代中欧摩尔多瓦Miciurin遗址(1500~1000 BC),在铁器时代出现在图瓦(619~609 BC)、 蒙古(400~300 BC)[15]。FX5属于世系B2亚组,该组的建立者最早也是出现在Miciurin遗址,稍晚的时候在宁夏于家庄墓地以及哈萨克斯坦Berel遗址也曾出现。从D1、D2和B2的出现时间和地点看,有一个明显的由西向东传播的趋势,考虑到中亚草原是家马的起源中心,我们的研究表明秦公一号大墓部分古代马的母系来自欧亚大陆西部,西北地区是家马进入中国的一条重要通道。
2. 秦与北方游牧民族的交流
春秋时期,秦与戎狄之间存在广泛的贸易交流活动,例如在关中地区可以见到狄人遗存中非常有代表性的器物——虎饰牌、虎形铜器、花格剑与铜釜[22]。田亚岐指出东周时期关中秦墓也受到了戎狄文化因素的影响,体现了秦与戎狄文化的相互影响和交融[23]。出于战争的需要,马成为最重要的家畜。秦国的地理位置使其可以很容易的从邻近的游牧民族那里引进优良的马匹。秦人与北方游牧人群的关系可以从其饲养的马匹可窥一斑。FX4、FX6和FX9在春秋战国时期出现在内蒙古忻州窑子、小双古城、新店墓地。这三个墓地都位于北方农牧交错带上,普遍流行用牛、马、羊殉牲,呈现典型的游牧人群特征。古代马FX5出现在宁夏于家庄墓地,该墓地也是游牧人群墓地。无论是与凤翔临近的宁夏地区,还是遥远的内蒙古地区都饲养相同的马,表明秦人与北方游牧人群存在广泛的贸易交流活动,同时也反映出古人在遗传育种上强烈的选择性。
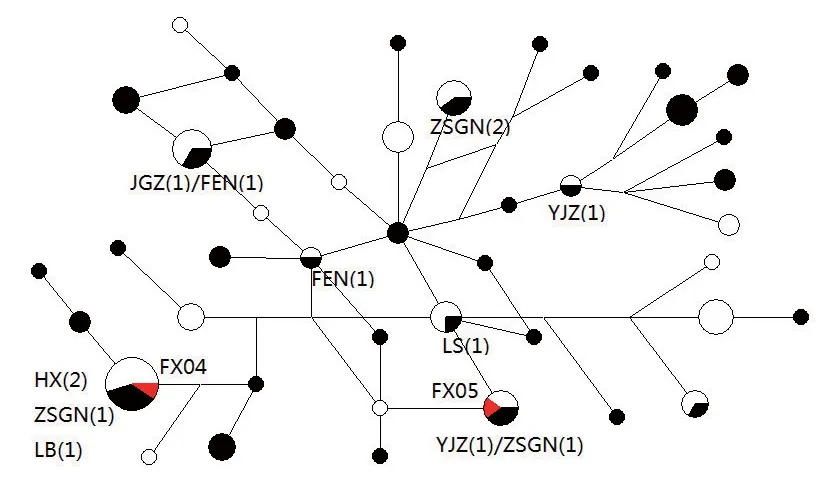
图三 中国古代马序列在土库曼阿哈尔捷金马中的分布图
3. 汗血宝马引入时间的探讨
张骞出使西域时,在大宛国(今费尔干纳盆地)发现了汗血马,归来时说:“西域多善马,马汗血,其先天马子也”。目前的科学研究表明,传说中的“汗血宝马”,实际上就是土库曼斯坦特产的“阿哈尔捷金马(Turkoman Akhal-teke)”。 许多世界著名的马种如纯血马、阿拉伯马、特雷克纳马都有阿哈尔捷金马的血统。汗血马作为一种珍贵、优良的马种,必定是古人重要的贸易对象,而秦国优越的地理位置使其很容易与游牧人群进行马匹的贸易交流,作为强大的秦国君主的陪葬车马坑,一定会选择最好的良马殉葬,因此能否在秦公一号大墓中发现汗血马的踪迹是我们关注的焦点问题。在共享序列搜索中,我们发现FX5的共享序列较少,主要是中亚马、阿拉伯马、北亚蒙古马和中国马。值得注意的是,FX5与两个现代土库曼阿哈尔捷金马序列(DQ327958、EU093049)完全相同。FX4的共享序列主要集中在西南西伯利亚和中亚,其中也两个阿哈尔捷金马序列(DQ327950、EU093051)这种情况引起了我们的注意,为了揭示中国古代马与阿哈尔捷金马的母系遗传关系,我们选择了现代土库曼阿哈尔捷金马的序列DQ327950–DQ327967,GQ119632-GQ119636,EU093045-EU093063作为对比,进行了中介网络分析(图三)。中介网络图显示西周时期陕西枣树沟脑和青海丰台遗址,以及春秋战国时期宁夏于家庄、内蒙古新店子、忻州窑子、小双古城、井沟子的古代马都与土库曼阿哈尔捷金马有关系,这表明汗血马的基因型很早就已经出现在西北地区,这远远早于汉代张骞出使西域的时间,这进一步验证了早在汉代之前,我国与西方就已经有了经济文化交流。
值得注意的是,中国古代马的基因型也出现在其他品种的家马之中,尽管这可能与古代先民培育马种的过程中选择良马有关,但中国古代马与汗血马的关系还需要进一步的研究,今后的古DNA全基因组研究有望为我们更清晰地揭示出中国古代马与汗血马的关系。
[1]袁靖. 中国古代家马的研究[C]//陕西省文物局,陕西省考古研究所,西安半坡博物馆.中国史前考古学研究.西安:三秦出版社, 2003, 436-443.
[2]傅罗文,袁靖,李水城.论中国甘青地区新石器时代家养动物的来源及特征[J].考古,2009(5):80-86.
[3]张春生.野马、家马及东亚养马中心[J].农业考古,2004(1):252-254.
[4]邓涛.中国矮马与普氏野马的亲缘关系[J].畜牧兽医学报,2000(1):29-34.
[5]奥尔森著,斯坦利 J.殷志强译.中国北方的早期驯养马[Z].1986:89-91.
[6]王宜涛.也谈中国马类动物历史及相关问题[N].中国文物报,1998-8-21.
[7]王志俊,宋澎.中国北方家马起源问题的探讨[J].考古与文物,2001(2):26-30.
[8]丁云,王言. 秦公一号大墓的发掘与秦史研究的新认识[J]. 渤海学刊,1988(3):80-84.
[9]Yang D Y,Eng B, Waye J S,et al. Technical note:improved DNA extraction from ancient bones using silica-based spin columns[J].Am J Phys Anthropol.1998, 105(4):539-543.
[10]Xu X,Arnason U.The complete mitochondrial DNA sequence of the horse,Equus caballus:extensive heteroplasmy of the control region[J].Gene.1994,148(2):357-362.
[11]Jansen T,Forster P,Levine M A,et al. Mitochondrial DNA and the origins of the domestic horse[J].Proc Natl Acad Sci U S A.2002,99(16):10905-10910.
[12]蔡大伟,韩璐,谢承志,等.内蒙古赤峰地区青铜时代古马线粒体DNA分析[J].自然科学进展,2007(3):385-390.
[13]蔡大伟,曹建恩,陈全家,等.内蒙古凉城县春秋时期古代马线粒体DNA分析[C]//边疆考古研究.北京:科学出版社,2008:328-333.
[14]Cai D,Tang Z,Han L, et al. Ancient DNA provides new insights into the origin of the Chinese domestic horse[J].Journal of Archaeological Science.2009,36(3):835-842.
[15]Cieslak M,Pruvost M,Benecke N,et al.Origin and history of mitochondrial DNA lineages in domestic horses[J].PLoS One.2010,5(12):e15311.
[16]赵欣,Antonia T.Rodrigues,尤悦,等.新疆石人子沟遗址出土家马的DNA研究[J].第四纪研究.2014(1):187-195.
[17]赵欣,李悦,陈洪海,等.陕西省淳化县枣树沟脑遗址马坑出土马骨的DNA初步研究[J].南方文物,2015(3):70-76.
[18]Warmuth V,Eriksson A,Bower M A,et al.Reconstructing the origin and spread of horse domestication in the Eurasian steppe[J].Proc Natl Acad Sci U S A.2012,109(21):8202-8206.
[19]Outram A K,Stear N A,Bendrey R,et al.The earliest horse harnessing and milking[J].Science.2009,323(5919):1332-1335.
[20]龚缨晏.车子的演进与传播——兼论中国古代马车的起源问题[J].浙江大学学报(人文社会科学版),2003(3):22-32.
[21]Keyser-Tracqui C,Blandin-Frappin P,Francfort H P,et al.Mitochondrial DNA analysis of horses recovered from a frozen tomb (Berel site,Kazakhstan,3rd Century BC)[J].Anim Genet.2005,36(3):203-209.
[22]杨建华.中国北方东周时期两种文化遗存辨析——兼论戎狄与胡的关系[J]. 考古学报,2009(2):155-184.
[23]田亚岐.东周时期关中秦墓所见“戎狄”文化因素探讨[J].文博,2003(3):17-20.
