光强对欧洲鹅耳枥秋季叶衰期光合性能及内源激素的影响
2018-08-02郭力宇吴驭帆祝遵凌
郭力宇,施 曼,吴驭帆,祝遵凌,4*
(1.南京林业大学 南方现代林业协同创新中心,江苏 南京 210037;2.南京林业大学 风景园林学院,江苏 南京 210037;3.南京林业大学 林学院,江苏 南京 210037;4.南京林业大学 艺术设计学院,江苏 南京 210037)
欧洲鹅耳枥(Carpinusbetulus)为桦木科鹅耳枥属落叶乔木,树种枝叶秀丽,秋叶金黄,耐荫性较强,是橡树林下的天然组分,在欧洲温带地区和小亚细亚森林群落中占有重要地位[1],同时以行道树和植篱形式广泛植于城市生态系统[2],是兼具观赏与城市生境颗粒物生物监测的一个适宜物种[3]。进一步研究表明其叶中可提取脱镁叶绿酸等活性物质应用于生物医药领域[4],近年来,国内也进行了部分园林推广,适应性良好[5-6]。
秋冬季节叶片衰老凋落限制了欧洲鹅耳枥秋叶的有效观赏期,对维持秋冬落叶季节生态系统碳平衡也有一定影响[7]。光照强度是叶片感知与衰老进程中的重要环境因子,暗诱导或强光环境下都会促使叶片衰老的发生,影响老化进程[8]。而在叶片衰老过程中常伴随着叶绿素降解,光合能力下降、养分流失以及内源激素失衡等一系列生理变化[9-10]。本研究通过比较与结合4品种(种)欧洲鹅耳枥秋季叶片衰老时在不同光照强度下的表现,探究影响欧洲鹅耳枥叶片衰老的主要生理变化规律,旨在为各欧洲鹅耳枥园林栽培的光照管理提供一定的理论参考。
1 材料与方法
1.1 试验地概况
试验地位于南京林业大学园林教学试验中心(32°04′N,118°48′E),气候温和,四季分明,光照充足,属亚热带季风湿润气候区。
1.2 试验材料
所用苗木为法国进口4品种(种)欧洲鹅耳枥,国内行盆栽管理,每株1盆,苗龄3 a,苗高80~100 cm,规格统一。分别为实生欧洲鹅耳枥原种(Carpinusbetulus)及其3个嫁接园艺品种(Carpinusbetulus‘Beekman’、Carpinusbetulus‘Frans Fontaine’、Carpinusbetulus‘Lucas’),砧木为原种欧洲鹅耳枥,砧穗径级相一致。下述原种和园艺品种分别以Original、品种名简称。
1.3 试验设计
采用盆栽试验法,以标准透光率为50%的遮荫网营造不同光强环境,设置3种光强梯度处理,双层、单层遮荫网下透光率分别为T25%、T50%,以不遮荫的全日照条件TCK为对照处理,处理时段设于2014年6月30日至9月30日,每处理10盆植株,重复3次,即每个处理30盆苗。盆与盆间距1 m,防止相互遮光。
1.4 测定指标与方法
1.4.1 叶绿素总量测定 采用乙醇丙酮混合液法[11]。洗净5片鲜叶,滤纸吸干后去除中脉并剪碎,称取0.05 g碎叶放入10 mL的离心管中,加入乙醇丙酮混合液(体积比1∶1)10 mL,parafilm封口后避光浸提24 h。待叶片组织完全变白后,取上清液分别在波长663、645 nm下测定吸光值,测定时间于叶衰老中期,各品种(种)每处理重复测定3次。
1.4.2 光合参数测定 叶衰老中期,选择晴朗无风、无云的9:00-11:30,采用英国PP-Systems便携式光合测定仪(CIRAS-2)测定各光合参数,设定光合有效辐射参数(PAR)为1 000 μmol·m-2·s-1,叶室温度25℃。测定净光合速率(Net photosynthetic rate,Pn)、蒸腾速率(Transpiratio rate,Tr)、气孔导度(Stomatal conductance,Gs)、胞间CO2浓度(Inter CO2concentration,Ci)等指标,各品种(种)每个处理随机测定3株,每株重复测定3次,选取向阳不遮荫的叶片,取平均值。
1.4.3 内源激素含量测定 从欧洲鹅耳枥变色衰老期前后开始取样,每隔10 d取样1次,10月20日开始至11月20日结束,共4次。样品取自冠层外围四面的叶片并置于液氮固定,在中国农大生理实验室采用酶联免疫吸附法(ELISA)测定叶片中内源生长素(IAA)、赤霉素(GA3)、脱落酸(ABA)的含量。
1.5 数据处理
应用Excel 2016软件进行数据的统计与绘图,在SPSS 22.0中进行单因素方差分析(One-Way ANOVA)及Duncan法多重比较(α=0.05)。
2 结果与分析
2.1 光照强度对欧洲鹅耳枥叶绿素含量的影响
在欧洲鹅耳枥叶片衰老期,4品种(种)对光强的响应存在差异(图1)。对于Beekman和Frans Fontaine,遮荫处理与对照差异显著,遮荫处理下的叶绿素含量显著上升, Frans Fontaine在T25%处理下叶绿素含量显著上升,Beekman在T50%即有明显变化,说明各品种(种)对光强响应敏感性有差异。而在Original与Lucas中,3处理下没有显著的趋势变化,Original在T50%处理下叶绿素含量最低,TCK与T25%含量相当;Lucas则在TCK和T50%处理下有较高的叶绿素含量,在T25%下叶绿素含量显著下降。同时发现,在自然全光照环境下Beekman与Lucas的叶绿素含量较高,与Original和Frans Fontaine达显著水平,说明此时Beekman与Lucas持绿性优于其他2个品种(种),衰老程度也可能较低。

注:大写字母表示不同处理同一品种(种)在0.05水平差异显著性,小写字母表示不同品种(种)同一处理在0.05水平差异显著性。下同。
2.2 光照强度对欧洲鹅耳枥光合特性的影响
2.2.1 不同光强下欧洲鹅耳枥净光合速率变化 净光合速率(Pn)是衡量叶片衰老期光合性能最直接的指标,也反映出低光处理下的胁迫响应程度。TCK处理下4欧洲鹅耳枥品种(种)均具有较高的Pn,甚至在T50%处理下Pn也处于2.7~4.4 μmol·m-2·s-1(图2),表明欧洲鹅耳枥衰叶中的叶绿体仍能有效地转化光能以及品种(种)对光较强的利用潜能。其次,随光强度的降低,各品种(种)Pn呈同步下降的趋势,TCK处理下Pn有最大值,说明在全光照条件下各欧洲鹅耳枥秋冬叶衰期进行光合作用累积产物的能力最大化。但这种现象在品种(种)间存在差异,Original在TCK处理下Pn是T50%、T25%处理的1.74、4.52倍;Beekman为1.48、2.67倍;Frans Fontaine为1.34、2.90倍;Lucas为1.06、1.14倍。可以发现嫁接品种相较原种变化幅度低,Lucas净光合速率则在3种光照强度下未表现显著差异,且均具有较高功效的光合速率,一定程度上反映其对光强的敏感性较弱,失绿度小,也与叶绿素含量吻合,其他品种(种)与叶绿素含量无显著趋同性。
2.2.2 不同光强下欧洲鹅耳枥气孔导度变化 气孔导度(Gs)是叶片气孔开张度与CO2利用效率的体现。4种欧洲鹅耳枥在3种光照强度下Gs的强弱一致表现为TCK>T50%>T25%,与Pn图示结果一致(图3),说明两者之间有一定的相关性,低光强对Gs的抑制作用是影响Pn的原因之一,但可能不是主要因素。

图2 不同光照强度下欧洲鹅耳枥Pn变化

图3 不同光照强度下欧洲鹅耳枥Gs变化
同时发现,Lucas在3种光照强度下,Gs涨幅显著(P<0.05),TCK处理为T50%、T25%处理的1.13、1.47倍,其余欧洲鹅耳枥品种(种)变化幅度较为稳定,可能反映了Lucas对低光强较强的气孔调节机制,对比Beekman,其在3处理下的Gs变化幅度与值均与Lucas相近,但在各处理Pn表现上,却显著低于Lucas,进一步佐证了叶衰期欧洲鹅耳枥Pn限制的主要因素可能不是气孔原因。
2.2.3 不同光强下欧洲鹅耳枥蒸腾速率变化 蒸腾速率(Tr)是叶片光合水分代谢的重要指标,叶片衰老干枯含水量下降与秋季环境湿度降低可能都会造成影响。与Pn、Gs结果相似,各欧洲鹅耳枥随着光强度的降低,Tr也表现为下降趋势(图4)。在TCK处理下,Original、Lucas的Tr均显著高于Beekman、Frans Fontaine,在T50%处理下,Frans与其他3品种(种)Tr差异达显著水平。说明在中度遮荫的环境下,Original与Lucas、Beekman仍存在高效的蒸腾作用。在T25%处理下4品种(种)的Tr均落至最低值,特别是Original与Lucas,相较T50%处理,4品种(种)降幅分别为47.1%、14.3%、4.0%、38.2%,即Original>Lucas>Beekman>Frans Fontaine,说明重度遮荫环境对Original的Tr影响较大。

图4 不同光照强度下欧洲鹅耳枥Tr变化
2.2.4 不同光强下欧洲鹅耳枥胞间CO2浓度变化 Lucas随着光强度的降低,细胞间CO2浓度(Ci)呈小幅上升趋势,稳定在328~344 μmol·mol-1(图5),进一步与其Pn表现衔接,其他3个品种(种)则呈现相反趋势,Ci逐渐升高,可能与胞间CO2作为光合作用原料有关,当植物Pn增大时,Ci应当下降,两者表现为负相关。同时,在TCK处理下Frans Fontaine的Ci值最小为271.67 μmol·mol-1,与其Pn在TCK下的表现有一定联系,而Frans Fontaine在Gs或Tr中3处理下均表现为小幅变化趋势,且值低于其他3个品种(种),结合Gs表现,再一次表明可能是品种(种)间的差异等其他因素导致了Pn的差异性,气体交换特性等对其影响微弱。
2.3 光照强度对欧洲鹅耳枥内源激素的影响
2.3.1 不同光强下欧洲鹅耳枥IAA含量变化 4品种(种)欧洲鹅耳枥叶片衰老期生长素(IAA)含量整体呈上升的趋势,但各品种(种)间以及处理间均存在差异(图6)。Original的T25%、TCK处理,Beekman、Frans Fontaine的TCK处理以及Lucas的T25%、T50%处理均呈现升-降-升的趋势,在10月30日以及11月20日有IAA含量的累积峰,且11月20日的IAA含量显著高于10月30日。在11月10日,各欧洲鹅耳枥3处理的IAA含量回落至低点。表明IAA可能具有双重性,在衰老前期促进欧洲鹅耳枥叶片的生长发育,而后期却加速了叶片的衰老。11月10日3处理均有较低的值,说明此时可能处于欧洲鹅耳枥缓慢衰老期,对环境变化有了一定适应性。对于处理而言,Original、Beekman后期在T50%条件下有较高值,Frans Fontaine、Lucas则在TCK条件下叶片IAA含量高,这与叶绿素、光合参数的结果有一定联系,欧洲鹅耳枥的品种差异可能导致了对不同光照强度响应的差异。同时,Lucas遮荫处理与对照条件之间IAA含量差异不显著,这也进一步说明其对光照条件的敏感程度较低。

图5 不同光照强度下欧洲鹅耳枥Ci变化
2.3.2 不同光强下欧洲鹅耳枥GA3含量变化 赤霉素(GA)属于生长促进剂,有促进叶片的生长发育,抑制叶片衰老脱落的作用。遮荫处理下各欧洲鹅耳枥叶片GA3含量随衰老进程呈持续上升的趋势,在11月10日衰老后期,遮荫处理下GA3含量剧烈上升达峰值(图7)。说明在弱光胁迫下,欧洲鹅耳枥通过上调叶片中GA3含量的抵御机制来维持其在叶片衰老期的养分供应。反观对照处理下,4欧洲鹅耳枥品种(种)呈缓慢下降的波动趋势,逐渐降低叶片中GA3含量,减缓叶片发育和养分消耗,促使叶片及时脱落。在Beekman、Frans Fontaine、Lucas叶片中GA3含量均表现T25%>T50%>TCK,在Original则表现为T50%>T25%>TCK,说明随着遮荫程度的加深,欧洲鹅耳枥叶片GA3含量会进一步上升,其中Original和Lucas在衰老后期GA3变化幅度最高,Original在T25%处理下涨幅为40.2%、T50%处理为51.0%,Lucas在T25%、T50%处理下分别上涨90.3%、51.8%,可能表明这2个品种(种)对弱光胁迫较强的GA3调节机制。
2.3.3 不同光强下欧洲鹅耳枥ABA含量变化 脱落酸(ABA)是植物受到胁迫以及衰老期的敏感激素指标。随叶片的衰老,叶片中ABA含量稳定上升,各欧洲鹅耳枥一致表现为T25%>T50%>TCK(图8),说明欧洲鹅耳枥叶片衰老与ABA具有一定相关性,随着叶片变黄干枯失水,ABA会在叶片中逐渐生成与累积,进而调控叶片的衰老与脱落,而在弱光胁迫下,随胁迫时间加长,处理与对照差异越明显,这种累积效应会随胁迫程度与时间的加深进一步增强,加速叶片的衰老凋落。4品种(种)处理间的差异大小为Original>Frans Fontaine>Beekman>Lucas,说明Lucas对于光强的敏感性低于其他品种(种),荫照条件下衰老程度与全光条件接近,更能适应荫庇环境。
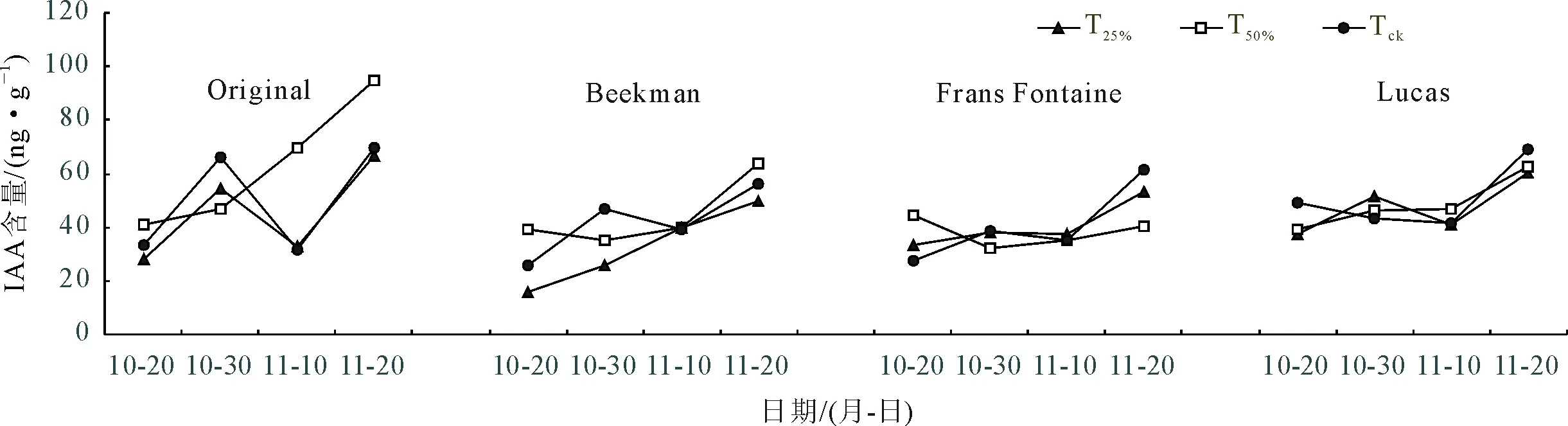
图6 不同光照强度下欧洲鹅耳枥IAA含量变化

图7 不同光照强度下欧洲鹅耳枥GA3含量变化

图8 不同光照强度下欧洲鹅耳枥ABA含量变化
3 结论与讨论
3.1 讨论
植物叶片秋季着色主要原因是叶片内色素比例的倒置,也表示植物叶器官的衰老。随着衰老进程,叶片中叶绿素一般呈持续下降的趋势[12],但在弱光逆境下,植物叶片叶绿素含量因品种、光照强度与时间长短变化较大[13]。本研究中,Beekman和Frans Fontaine在遮荫处理下,叶绿素含量与对照相比显著上升,且两者分别在中度遮荫处理和重度遮荫处理下表现显著差异,而Original与Lucas并未出现显著的波动,表明欧洲鹅耳枥各品种(种)对光强的响应存在差异。低光条件下叶绿素含量的上升,可能是一种补偿效应[14],叶片通过产生较多的叶绿素来吸收光量子以补充光照条件的不足,或是维持了叶绿体结构,进而满足树体生长的需要。持续光照或强光对色素具有漂白作用[15],全光照条件下欧洲鹅耳枥的叶绿素含量低于荫照处理,也可能是光产生色素光氧化的副效应导致,这些在枫香[16]、紫叶李[17]等秋色叶植物转色的研究中也有一定体现。
植物叶片的光合能力随衰老时段同步下降,因此通过测定有关的光合参数可以较好地反映叶片光合功能是否衰退[8]。净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、细胞间CO2浓度(Ci)是常见的衡量叶片光合功能的指标。各欧洲鹅耳枥的净光合速率、气孔导度、蒸腾速率随着遮荫度的加深逐渐下降,而胞间CO2浓度则呈现相反的趋势,随遮荫度的加深逐渐升高,原因是胞间CO2浓度作为光合作用的原料被消耗[18-19],全光照条件下欧洲鹅耳枥叶片能更有效地执行光合作用,CO2消耗量更多。此外,研究还发现,各欧洲鹅耳枥即使处于衰老期与荫照条件下,叶片具有较高的净光合速率,说明其在衰老期仍能进行高效的光合作用,这因树种而异,与Zhang[7]、罗紫东[9]等在珍珠花、枫香叶片衰老过程光合能力的研究结果一致,有利于增加秋冬季节碳吸收量、延长碳吸收时间。但要指出,各品种(种)净光合速率在3处理下的表现与叶绿素并不同步,Beekman在荫照条件下叶片中叶绿素的含量较高,但并未表现在其光合的功能性上,结合两点,可以推测欧洲鹅耳枥叶衰老期叶片中的叶绿素含量已足够在叶衰期进行正常的光合作用,并不对光合功能产生限制[19],从侧面反映出欧洲鹅耳枥较强的光利用潜能。
内源激素是植物叶片老化过程中主要的生理调控因子之一[21]。叶片衰老常伴随着IAA、ABA含量倍增,GA含量下降的一系列变化[10],以促进叶的脱落。随叶逐渐衰老,各欧洲鹅耳枥叶IAA含量在10月30日和11月20日有较高的积累,且后时段值更高,表明IAA在叶衰期可能具有双重性[22],在衰老前期促进欧洲鹅耳枥叶片的生长发育,而后期却加速了叶片的衰老,由于处于衰老期,这种前期的促进作用与生长期促进效率不同,因此低于IAA启动加速衰老功能的含量。在4品种(种)中IAA含量也存在差异,Lucas荫照处理与全光条件下,衰老叶中IAA含量相近,表明Lucas品种可能属于光照的弱敏感型及强适应型,通过基因型对光能的强捕捉力,使暗呼吸速率减少从而减缓有机物在衰老期的分解以适应胁迫环境[23-24]。GA3在4品种(种)中荫照与对照处理的表现相反,在对照处理中GA3值缓慢下降或略有浮动,而在荫照条件下,则随着处理时间呈持续上升的趋势,表明弱光一定程度上加速了欧洲鹅耳枥叶片衰老的进程,而导致树体养分供应不足,叶中GA3受调节含量上升,以抵御弱光胁迫带来的影响。但也有研究表明叶中GA3有抑制叶绿素分解的作用,两者具有协同性[25],弱光条件下品种(种)叶绿素含量上升可能原因是受到GA3含量上调的影响。ABA被称为“胁迫激素”,在逆境中会大量增加,加强植物抗逆性,各品种(种)叶中ABA含量一致表现为重度遮荫>中度遮荫>全光照条件,并随着叶的老化含量缓步上升,表明欧洲鹅耳枥叶片衰老与ABA含量相关性较高,这与多数植物叶落衰老ABA含量上升相似[26-27],ABA可能作为欧洲鹅耳枥叶衰期主要调控激素启动叶片的衰老,且弱光条件进一步缩短各欧洲鹅耳枥叶片的衰老进程,但Lucas在不同处理间较其他品种(种)差异程度低,说明Lucas品种更适宜弱光环境的应用。
3.2 结论
全光照条件下,4品种(种)欧洲鹅耳枥秋季叶衰期光合性能最佳。Carpinusbetulus‘Lucas’叶片中叶绿素含量、净光合速率、内源激素含量在不同光照强度下差异不显著(P>0.05)或较其他品种(种)差异小,对光强的敏感性弱,在弱光条件下有较强的适应性,具有更广的光适应范围;净光合速率和内源激素ABA是欧洲鹅耳枥叶落期叶衰老较佳的指示指标;各欧洲鹅耳枥叶衰期对光能仍有高效利用性,这对维持秋冬季节生态系统碳平衡有利,延长了碳同化期,可以同时结合各品种(种)的观赏特性、生长习性进行园林栽培与推广。
