大理州温泉捕食线虫真菌资源调查研究*
2018-07-26范喜杰杨晓燕
范喜杰,张 欣,张 发,杨晓燕*
(1.大理大学公共卫生学院,云南 大理 671000;2.大理大学农学与生物科学学院,云南 大理 671003;3.大理大学东喜玛拉雅研究院,云南 大理 671003)
捕食线虫真菌(Nematode-trapping fungi)是一类以营养菌丝特化形成黏性捕食器官(黏性菌网、非收缩环、黏球和黏性分枝)及机械捕食器官(收缩环)捕食线虫的真菌[1―2]。由于该类真菌在形态和生态功能上的特殊性及其在线虫生物防治中的潜在价值,逐渐成为微生物学研究的热点之一[3―5]。捕食线虫真菌广泛分布于世界各地,甚至在南极[6]、热泉[7]和重金属污染区[8]等极端环境中都有分布。到目前为止,绝大部分研究都集中在陆生环境中的捕食线虫真菌[9―11],对于水环境中捕食线虫真菌的研究相对较少[12―14]。目前全世界已知的2万多种线虫,有许多生活在水环境中[15],水体中丰富的线虫种类必定伴随着丰富的水生捕食线虫真菌的存在。从已有文献报道可知水体中水生真菌资源丰富,生物多样性极高[16,17]。
众所周知,温泉是一种特殊的水生环境,广泛分布于全球各地。由于温泉与地球早期环境比较接近,温泉中的微生物生态系统相对简单、稳定和封闭。同时,在数千万年的进化历程中,温泉微生物已经具备独特的生存适应能力,这些微生物所具有的特殊生理机制和蕴涵的独特基因资源,产生的酶或其他代谢产物及其在生命起源和生物进化过程中所处的特殊地位,已经受到国内外学者的广泛关注[18―20]。如:有研究者从美国黄石公园温泉中分离得到嗜热细菌Thermusaquaticus[21],从云南腾冲温泉中分离得到硫化叶菌Sulfolobustengchongensis[22],从冰岛温泉中分离得到氢氧化细菌Hydrogenophilusislandicus[23],这些皆表明温泉中蕴藏着丰富的微生物资源。而目前对温泉中微生物资源的研究主要针对放线菌、嗜热真菌、细菌和古生菌等,国内外仅Hu等[24]报道过一株温泉食线虫真菌(Arthrobotrysmultisecundaria),该菌分离自云南腾冲温泉,表明温泉捕食线虫真菌资源还未引起国内外学者的关注。云南省温泉资源非常丰富,是中国地热资源最丰富的地区之一,所以本研究对大理州人为干扰较少的温泉进行捕食线虫真菌资源调查,了解温泉中捕食线虫真菌资源的分布状况,分离、鉴定和保存温泉捕食线虫真菌资源,为捕食线虫真菌的起源和进化研究提供菌种资源。
1.材料和方法
1.1 样品采集
2012年3―5月,采集大理市、祥云县、弥渡县、剑川县、漾濞县的温泉底泥共105份,样品于4℃保存,7d内处理完所有采回的泥土样品。样品采集的同时测定温泉pH值和温度。
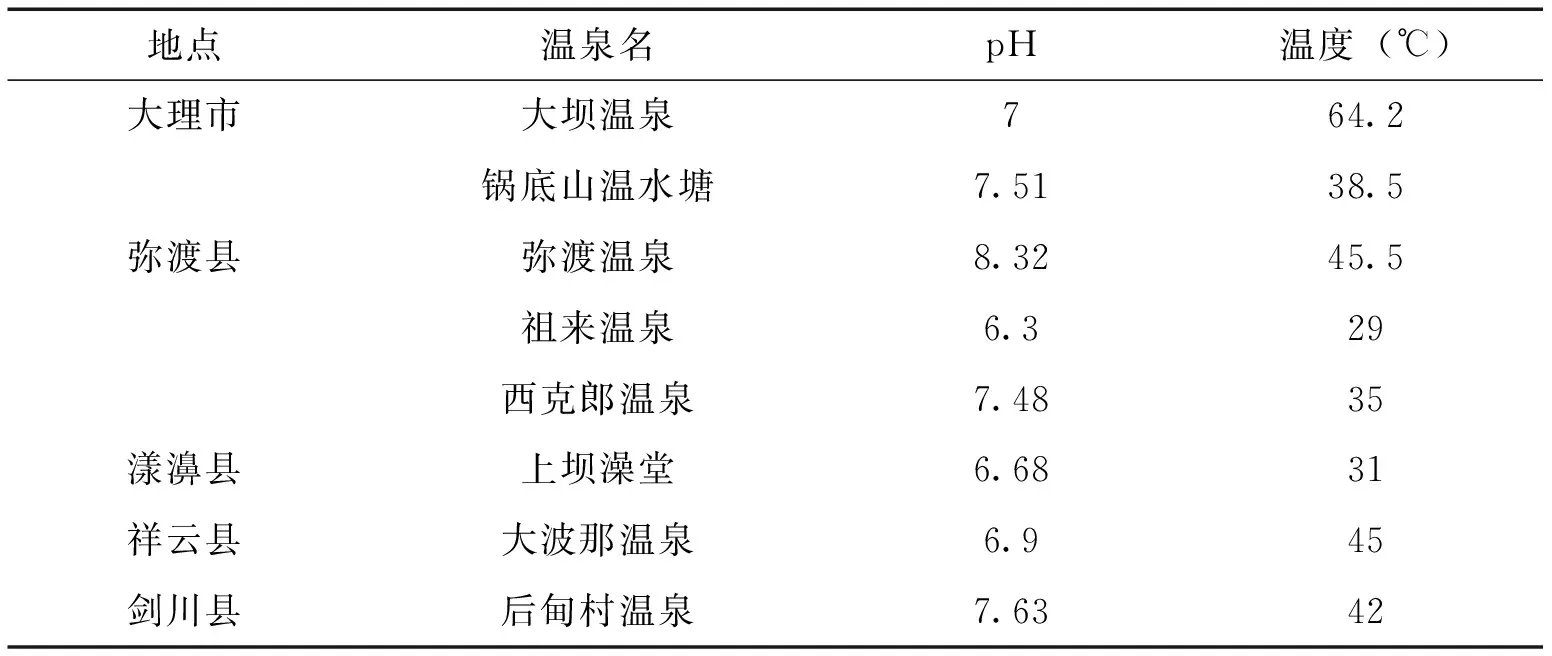
表1 各温泉基本信息
1.2 CMA培养基的制备
玉米粉20g,琼脂18g,水1000mL,自然pH。玉米粉加水适量煮沸30min,四层纱布过滤,滤液加水至1000mL,混匀,分装,121℃高温灭菌30min[25]。
1.3 诱饵线虫的制备
采用贝尔曼漏斗法[26]制备:即取少许全齿复活线虫Panagrellusredivivus放于四层擦镜纸上置于漏斗(漏斗通过软管与收集管相接),加适量无菌水,挤压软管排出气泡,静置一定时间,直至滤出物达到需要量,加无菌水清洗线虫,使线虫浓度约为5×103条/mL。
1.4 捕食线虫真菌的培养、分离与鉴定
采用常规撒土法培养[27],每个样品设置3个重复,撒样后在平板中加入上述线虫悬液1ml(约5000条),室温下诱导培养4周后在体视镜下进行镜检,将镜检到的菌株转接到CMA平板上,并在培养基中间挖一个2×2 cm2的观察室,在30℃的恒温箱内培养5―7d,待观察室长满菌丝后加入诱饵线虫,培养12―24h后进行镜检,观察捕器的产生情况[28]。根据孢子着生方式、孢子梗形态、孢子形态及捕器类型等特征采用Li等[29―30]分类系统对捕食线虫真菌进行形态学鉴定。
1.5 数据处理
检出频率(OF):OF = 某个种出现的土样数/总的土样数×l00% 。
Ni为第i种的数量;N为所有种的数量;Pi为第i个种所占百分数;S为采样点的物种。
2.结果
从大理州温泉中共分离到2属14种捕食线虫真菌,其中Arthobotrys12个种,Dactylellina2个种;在祥云县的大波那温泉中分离出一个新种(A.xiangyunensis,sp.nov)[32];A.musiformis为温泉捕食线虫真菌的优势种(检出率为26.92%)。弥渡县的弥渡温泉中捕食线虫真菌多样性较高,为0.99,其次为祥云县的大波那温泉(0.69);产黏性菌网的捕食线虫真菌占总检出的86.54%(表2)。

表2 温泉底泥中捕食线虫真菌资源调查结果
3.讨论
本研究从大理温泉中共分离、鉴定出2属14种捕食线虫真菌,发现一个新种(A.xiangyunensissp.nov)[32],表明大理温泉中存在较为丰富的捕食线虫真菌资源。在调查的温泉中,各温泉(包括县内的温泉之间)检出种皆不同,其主要原因可能是:温泉作为与地球早期生境最为接近的一种特殊生境,其中保留了最为原始的菌种资源[21―23,33,34],在地球生物演化的过程中受到的人为干扰较少,使得空间距离上较为接近的温泉之间保留了各自的特有物种。同时,各温泉间的生境差异,如pH和温度的差异,也可能会导致生活在其中的菌种的不同。捕食线虫真菌的最适生长温度为25―28℃,而本次调查的温泉温度范围为29―64.2℃, 表明捕食线虫真菌的温度适应能力也存在菌株差异,这种差异由遗传物质导致。因此,对温泉捕食线虫真菌菌种资源的发掘可获得常规生境中捕食线虫真菌没有的基因资源。随着人类对温泉的开发加剧[35―37],分布其中的独特的菌种资源也必将受到影响,因此尽快发掘和保存温泉菌种资源是一项很迫切的研究工作,同时,应当从政府层面加强对温泉资源的保护。A.musiformis在大理温泉中的检出率最高,是大理温泉中捕食线虫真菌的优势种群;A.oligospora在其中检出率较低,且仅在大理市的大坝温泉中有检出,但已有文献报道A.oligospora是不同生境土壤中的优势种群[6, 38,39],表明不同生境中捕食线虫真菌的分布存在一定差异,产相同捕食器官的捕食线虫真菌也存在环境适应能力的差异。
据化石记录显示,食肉类真菌大约起源于1亿年前的中生代,在当时,这个古老的类群主要营腐生生活,但在漫长的进化历史中,其菌丝体为适应生存环境(如氮源缺乏),可变态为各种形态独特、结构精巧的捕食器官捕食线虫[40,41]。因此,捕食器官的进化也就成为该类真菌进化的主要特征之一,不同基于捕食器官形态、捕食率和分子系统发育学特性的进化假说也相继被提出[25,30,42―45]。但在这些假说中,各捕器类型的进化地位也各不相同,甚至完全相反,如:李天飞等[25]和Yangetal.,[43]提出黏性菌网是一种比较古老、进化地位低且捕食率也较低的捕器类型,但Lietal[30]、Rubner[44]和Enceetal.[45]则认为黏性菌网是一种进化地位比较高且捕食率也较高的捕器类型。林慧彪等[46]对产不同捕器类型的捕食线虫真菌进行捕食率的定量测定后发现,产黏性菌网的捕食线虫真菌的捕食率相较产其他捕器类型的捕食线虫真菌低,而本研究结果也显示产黏性菌网的捕食线虫真菌为大理温泉中的优势种群,考虑到温泉生境的特殊性,产黏性菌网的捕食线虫真菌可能更原始。
