广东木荷资源的基因型特征分析
2018-07-07汪迎利陈一群蔡燕灵连辉明何波祥曾令海
汪迎利 ,陈一群 ,蔡燕灵 ,连辉明,何波祥 ,曾令海 ,陈 琪 ,张 谦
(1.广东省林业科学研究院 广东省森林培育与保护利用重点实验室,广东 广州 510520;2.广东省林业科技推广总站,广东 广州 510173)
植物在适应生长环境的过程中,常进化产生复杂的种内遗传变异,形成丰富的遗传种质资源。这些变异在林木上表现为树体形态、结构、生理和生长等方面的差异[1-2]。木荷Schima superba是木荷属植物中分布最广的物种,在我国主要分布于北纬31°以南、东经105°以东的广大地区,自然生境纷繁复杂,进化形成了极其丰富的种内遗传变异[3-6]。研究表明,木荷苗木和幼林的生长、形态、木材基本密度等性状存在显著的种源、家系、个体差异[7-12];树高、胸径和冠幅受中等偏强的遗传控制,树干通直度和分枝性状受中等偏弱的遗传控制;以生长量和木材密度为选择标准,筛选出一批优良的木荷种源、家系和单株,为我国生物防火林带和速生珍贵阔叶用材林等建设提供了重要种植材料[9,11]。
广东省是木荷主要分布区和生产区,受复杂多样的地形、地貌和气候影响,木荷遗传资源十分丰富[3,6]。目前,参与早期遗传测定研究的广东材料仅有少量局部种源,全省木荷群体研究仅有叶片表型多样性分析[13],针对该区域木荷资源系统清查、遗传多样性以及生长与形质性状的分析仍鲜有报道。为此,本研究在广东省17个县市广泛开展了天然木荷资源普查和优树选择,收集保存了55株优树的种质资源。根据天然隔离物种进化形成独立基因型的特点[2],55株优树的天然种质资源即为55个基因型。为了研究木荷天然资源的基因型特征,建立各基因型自由授粉子代测定林,开展生长与形质性状的相关性分析、主成分分析和聚类分析等研究,挖掘可为木荷良种选育提供技术支撑的基础数据。
1 材料与方法
1.1 试验材料
从2003年开始,在广东省17个市县开展木荷天然林普查,运用“五株优势木”对比法进行优树选择[14],共选择优树55株,即55个基因型(图1),采集全部优树自由授粉种子,对照CK为广东本地普通生产种的种子,于2004年1月播种育苗。木荷基因型命名方式为采集地拼音首字母加数字编号,如“CZCA102”表示潮州潮安采集的102号基因型。

图1 木荷基因型采集地分布图Fig.1 Geographic location of collection sites for Schima superba genotypes
1.2 试验地概况
试验地广东省东江林场位于广东省河源市紫金县临江镇,东经 114°44′,北纬 23°38′,海拔100~150 m,坡度12°~17°,年均气温20.9~21.5 ℃,年均积温6 997.6 ℃,夏季最高气温38.0 ℃,冬季最低气温-2.0 ℃,年降水量1 600 mm,土壤为赤红壤,土层厚90 cm,腐殖土8 cm。
1.3 试验设计
测定林于2005年5月营建完成。测定林以家系/基因型为单位,采用完全随机区组设计,每小区4株,重复8次,55个家系栽种在山脉同一坡向试验地,栽种密度为2.5 m×2.5 m。
1.4 树木观测
分别于2008年(3 a)、2011年(6 a)、2014年(9 a)和2016年(11 a)对测定林进行生长与形质性状调查,生长性状包括树高(HT,m)和地径/胸径(CD/DBH,cm),形质性状包括主干通直度(STS)、分叉干数(STN)、分枝角度(AGL)、主侧枝基径(LBD,cm)和枝下高(HUB,m)。树干通直度按通直、一般弯曲、较弯曲和严重弯曲分为4级,分别记为1、2、3和4,即分值越低越通直。分枝角度分为4级,依次为0°<X1≤ 22.5°、22.5°< X2≤ 45°、45°< X3≤ 67.5°和67.5°<X4≤90°,分别记为1、2、3和4,即分值越低分枝角越小。分叉干数为1.2 m以下实测数。
因单株材积由树高和胸径计算得出,易影响后续相关性分析、主成分分析和聚类分析结果,进而影响遗传多样性分析结果,故本研究不考虑单株材积性状。
1.5 数据分析
采用SAS软件PROC GLM、CORR、MIXED和IML模块,分别进行方差分析和相关性分析(SAS Institute Inc.,Cary,NC)。分析分枝角度、主干通直度和分叉干数前,先对数据进行X-1/2转换[15]。方差分析模型为Yijk=μ+Fi+Bj+(FB)ij+ek(ij),式中μ为总均值,Fi为第i个家系的效益,Bj为第j个区组的效益,(FB)ij为家系与区组的交互作用,ek(ij)为剩余误差。变异系数、单株遗传力、家系遗传力的估算参照先前文献进行[15-17]。主成分分析与聚类分析涵盖全部13个性状,分别采用SAS软件PROC PRINCOMP和CLUSTER模块进行数据分析[2,18-19]。为了避免各性状的读数级差对聚类分析的影响,对各性状的数值进行Z值转换[2,19]。
2 结果与分析
2.1 生长与形质性状分析
表1列出了木荷55个家系/基因型生长性状与形质性状的统计数据。树高、胸径呈逐年增长趋势,到11 a时均值分别达7.81 m和9.69 cm;形质性状中,主干11 a时已出现明显整枝,枝下高均值达1.96 m;侧枝的分枝角指数达1.88,即角度在67.5°以上,说明分枝角度较大;主干通直度指数仅为1.40,属一般弯曲;分叉干数均值为1.17,说明多为单一主干;主侧枝基径均值仅为2.93 m,处于较为合理水平。方差分析发现,8个生长性状和4个形质性状(枝下高、通直度、分叉干数和主侧枝基径)在家系间均存在极显著差异(P< 0.01),表明参试家系蕴含丰富的遗传变异,进行遗传选育与改良的潜力较大。

表1 木荷基因型13个性状的统计信息†Table 1 General information of 13 traits studied for Schima superba genotypes
变异系数分析发现,全部13个目标性状均具有较高的遗传变异系数,幅度在6.90%~50.43%之间。生长性状中,径生长的变异系数在27.29%~43.78%之间,高生长变异系数在16.90%~34.36%之间,即径生长的变异系数大于高生长的变异系数;变异系数最大的是3 a基径(CD3),达43.78%,最小的为11 a树高(HT11),仅16.90%。年度间比较发现,高生长与径生长的变异系数整体上均呈逐年下降趋势,即3 a时最大,11 a时最小。与生长量性状相比,形质性状的变异系数较大,11 a时均大于32%,其中分叉干数和主侧枝基径变异系数分别高达45.50%和50.43%。这进一步说明,8个生长性状与5个形质性状存在丰富的家系遗传变异。
遗传力分析结果显示,木荷13个性状的单株遗传力变化幅度较大,整体上生长性状单株遗传力高于形质性状。生长性状单株遗传力在0.177~0.540之间,其中HT3最小,仅0.177,HT11最大,达0.540;形质性状单株遗传力在0.014~0.276之间,其中AGL11最小,仅0.014,LBD11最大,为0.276。受非加性效应存在的影响,木荷13个性状的家系遗传力明显高于单株遗传力,且具有较低的标准差,其中生长性状家系遗传力在0.763~0.875之间,形质性状遗传力在0.620~0.789之间。这些结果说明,木荷主要生长与形质性状受较强的遗传控制,且遗传控制较为稳定。
2.2 性状间相关性分析
表2列出了木荷13个性状相互间的Pearson与遗传相关系数。Pearson分析结果显示,8个生长性状间均存正相关关系,全部28组相关系数中有24组相关系数达到显著(P<0.05)或极显著水平(P<0.01),未达到显著水平的系数均为3年生与其它年度的相关性。3年生地径(CD3)与树高(HT3)的相关系数虽高达0.78,但它们与6~11 a的生长性状间相关系数偏低,其中CD3与它们的相关系数在0.21~0.37之间,HT3与它们在0.21~0.40之间;6~11 a生长性状相互间正相关极显著(P<0.01),相关系数在0.62~0.92之间。5个形质性状间,仅有主侧枝基径(LBD11)与分枝角(AGL11)存在极显著负相关,相关系数为-0.36;其它形质性状间均不存在明显的相关性。生长性状与形质性状间的相关性存在明显分化。枝下高(HUB11)和主侧枝基径与6~11 a生长性状间均存在正相关,其中HUB11与其相关系数在0.20~0.36之间,主侧枝基径(LBD11)与其相关系数在0.16~0.34之间。剩余的3个形质性状与生长量性状间的相关性无明显规律。
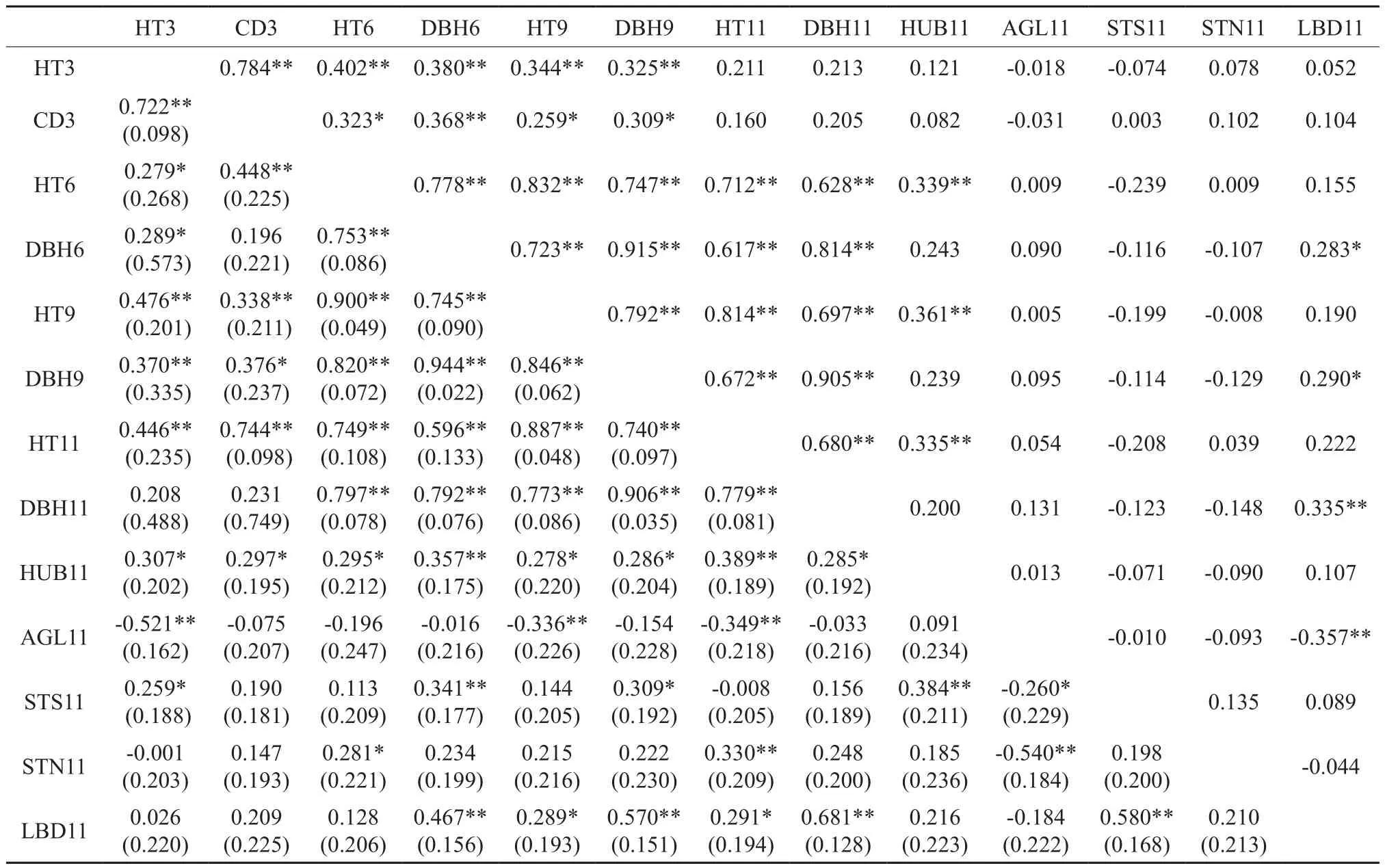
表2 木荷13个性状的Pearson与遗传相关系数†Table 2 Pearson and genotypic correlations of 13 traits for Schima superba
遗传相关分析发现,8个生长性状间亦均存在正相关关系,全部28组相关系数中有25组达到显著或极显著水平,未达到显著水平的系数亦为3 a生与其它年度的相关性,分别是HT3与DBH11、CD3与DBH6和DBH11。5个形质性状间,4组遗传相关达到显著或极显著水平,其中HUB11与STS11显著正相关,相关系数为0.384;AGL11与STS11和STN11均呈显著负相关,相关系数分别为-0.260和-0.540;LBD11与STS11亦呈显著负相关,系数为-0.580。生长与形质性状间,8个生长性状均与HUB11显著或极显著正相关,系数为0.278~0.289;6~11 a的5个生长性状亦与LBD11显著正相关,系数为0.289~0.681;其它性状间的遗传相关无明显规律。
2.3 主成分分析
为鉴别木荷最关键的表型性状,以便用最少的性状区分木荷不同基因型,并揭示各基因型之间的相互关系[2],本研究对木荷13个性状进行主成分分析,从表3可知,前3个主成分解释了总变异中的68.48%,说明木荷多数性状间存在紧密相关性。第1个主成分解释了总变异的45.00%,主要贡献性状为6~11 a生的高生长和径生长,包括 HT6、HT9、HT11、DBH6、DBH9 和 DBH11;第2主成分解释了总变异的13.05%,主要贡献性状为3 a生的生长性状(CD3和HT3)与11 a生通直度(STS11);第3主成分解释了总变异的10.43%,主要贡献性状为11 a生的形质性状,包括AGL11、STS11和STN11和LBD11。
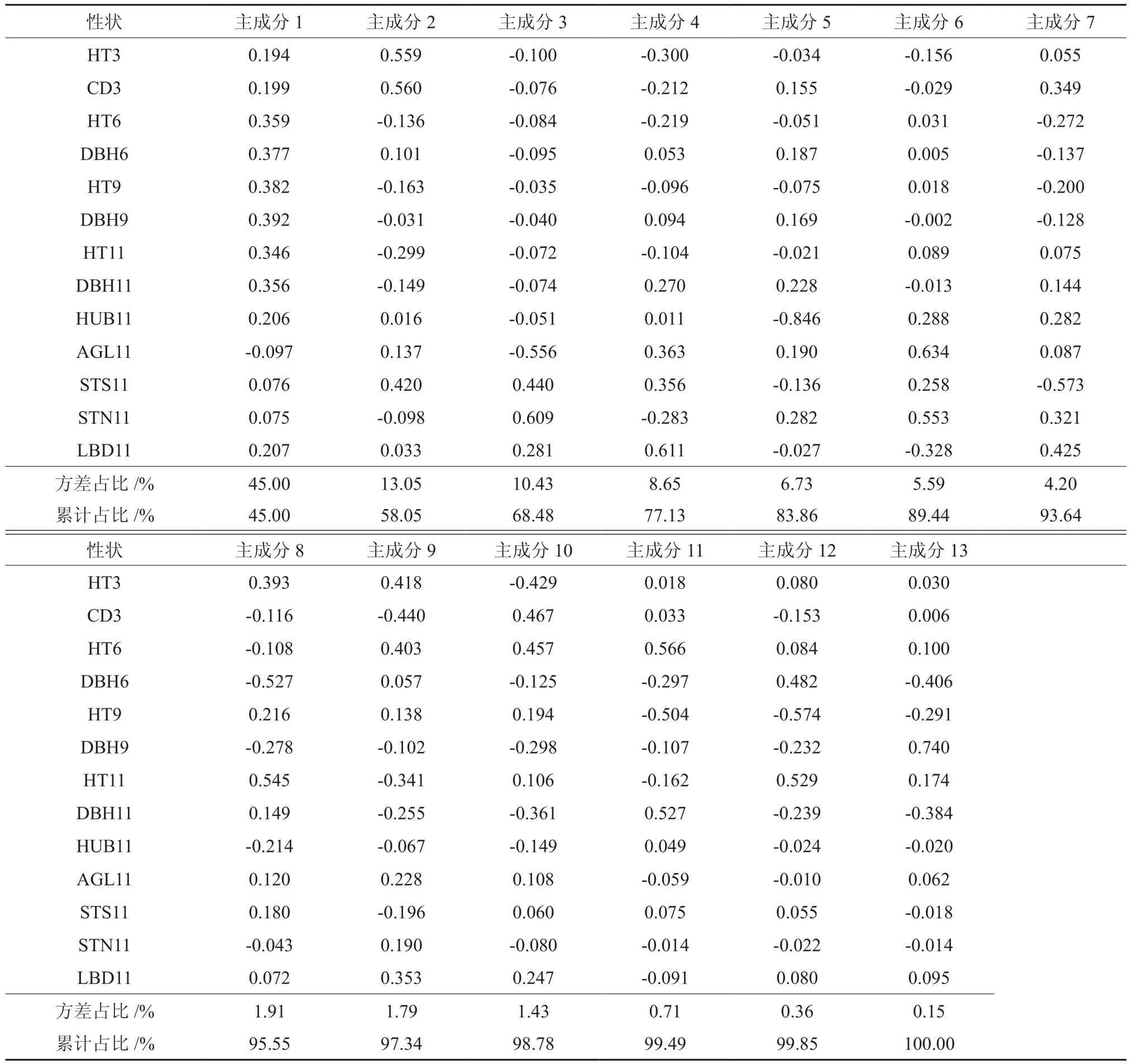
表3 木荷13个性状主成分分析的主成分轴特征值Table 3 Eigenvalues of the principle component axes from PCA of 13 variables in Schima superba
为分析生长性状与形质性状间的相互关系,以便鉴别不同木荷基因型,本研究构建了主成分分析散点图。从图2可知,木荷55个基因型13个性状的第1、2主成分二维散点图中,绝大部分木荷家系集中分布在一起,呈现出近似的生长性状和主干通直度;有2个家系(CZCA102和JMKP107)分布在左下部区域,即具有较差的3 a生长与适中的6~11 a生长;1个家系(HZLM105)分布在散点图右侧,即具有很高的6~11 a生长。第一、三主成分散点图显示,木荷不同基因型间的形质性状差异巨大,形质性状表型最优的有QYQX110、SGWY118和MMGZ101等家系,表现较差的有YJYC104、SGWY106和ZQHJ111等家系。
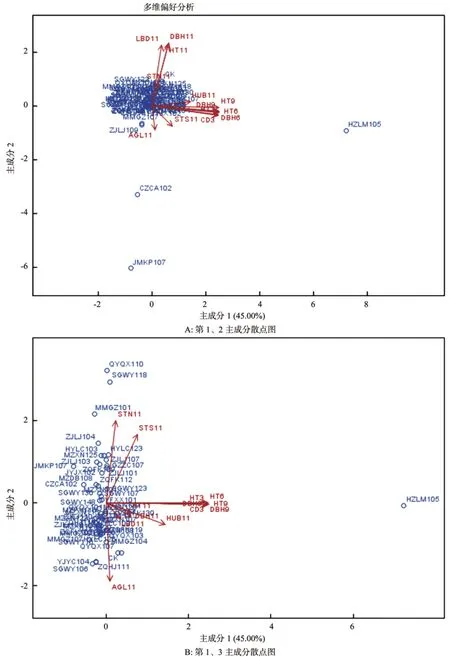
图2 主成分分析二维散点图Fig.2 Bi-plot for the studied Schima superb based two principle components
散点图箭头方向表示性状间的相互关系[20]。从第一、二主成分散点图可知,多数6~11 a生长性状趋于同一方向,HUB11与多数生长性状趋于同一方向,LBD11与DBH11和HT11亦趋于同一方向,表明它们相互间具有紧密的正相关关系;AGL11与DBH11、HT11和LBD11的箭头方向相反,表明它们之间具有负相关关系;这些结果与Pearson和遗传相关性分析结果高度吻合。
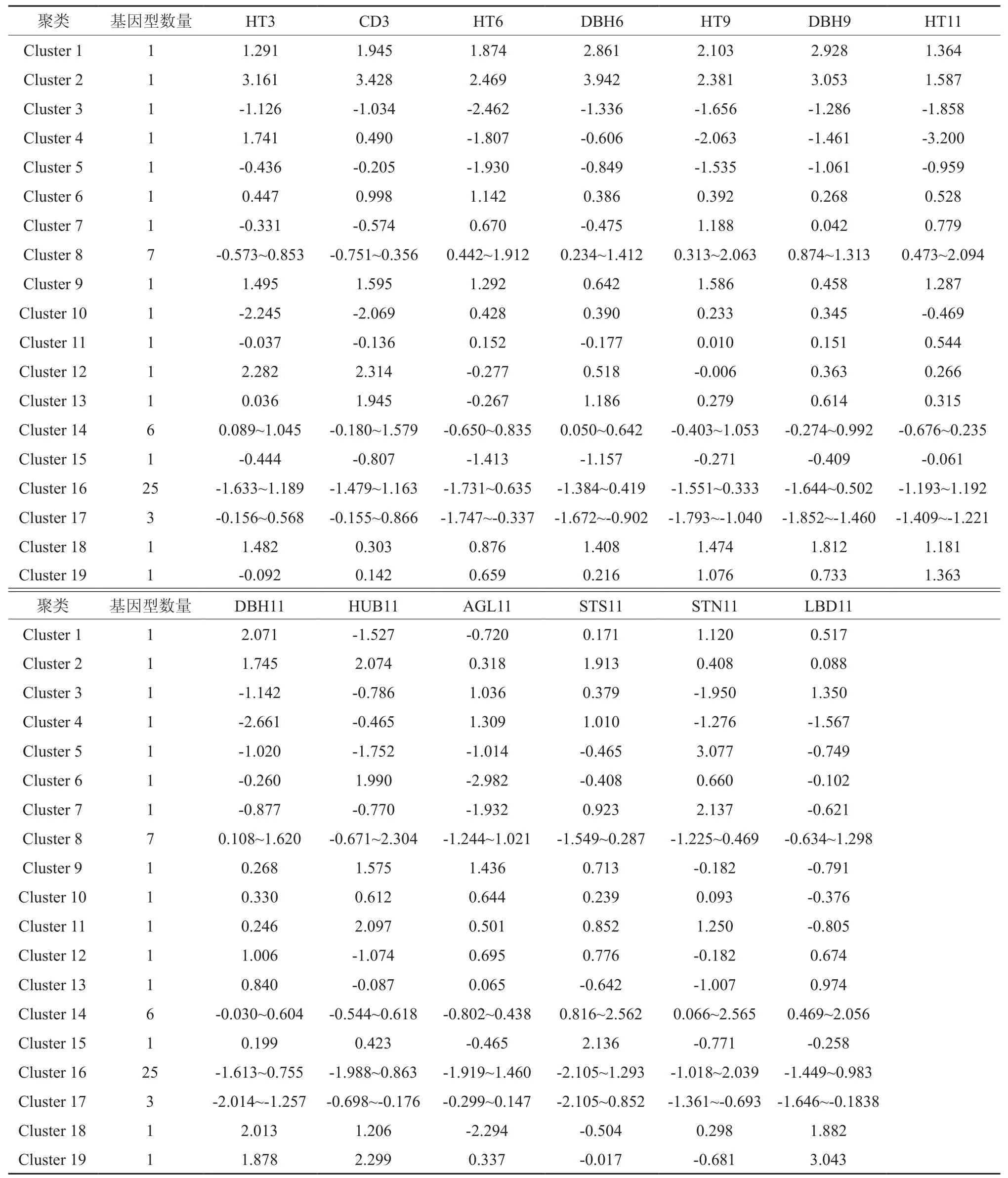
表4 木荷各聚类内13个性状的Z值Table 4 Z scores of 13 traits of Schima superba within each cluster
2.4 聚类分析
对木荷55个基因型13个性状的平均值进行聚类分析,生成如图3所示的系统树。按照聚类质心点距离(即表型相似性)为0.6的标准,55个木荷基因型被划分成18个聚类,对照为第19个聚类。各类含有不同数量的基因型,同一类中相邻家系大多来自相同的地理区域或临近区域。
表4列出了各类木荷的特征Z值。聚类内部基因型间具有近似的生长或形质性状,但聚类间的木荷表型差异巨大。聚类16为最大的集群,包含25个基因型,该类木荷具有中等或偏低的6~11 a生长量,但3 a生长量和11 a形质性状无明显特征;聚类8是第二大集群,含7个基因型,该类木荷具有中等3 a生长量、中等或偏高的6~11 a生长量、中等或偏低的通直度与分叉干数、中等或偏高的枝下高和主侧枝基径,分枝角度无明显特征;聚类14为第三大集群,含6个基因型,该类木荷具有中等的6~11 a生长量、较好的通直度以及中等枝下高和分枝角。聚类17为第四大聚类,含3个基因型,该类木荷具有中等的3 a生长、较低的6~11 a生长、较差的形质性状等。其余14个聚类均只含有1个基因型,且该木荷基因型具有明显不同的生长与形质性状。例如,聚类1为基因型SGWY123,具有较高的生长量性状、较多的分叉干数、较低的枝下高和分枝角、中等的通直度和主侧枝基径。

图3 基于13个生长与形质性状的55个木荷开放授粉家系聚类分析Fig.3 Cluster analyses of 55 open-pollinated families of Schima superb progenies based on 13 growth and form traits
3 讨论与结论
(1)广东地处木荷中心种源区和中部种源区,其种质资源不仅最为速生,且遗传多样性最高[9]。本研究在全省17个市县选择、收集和保存了木荷优树材料55份,覆盖了省内全部天然分布区,储备了一批优良的基因资源,为今后的选育研究奠定了物种基础。
(2)采用变异系数分析、方差分析与主成分分析等方法,对自由授粉子代进行多年度观测分析,证实木荷基因型遗传品质对生长和形质性状具有显著影响。木荷表型差异在其它地理区域的生长、形质、材性等性状测试中亦有类似发现[8-11],其遗传多样性也在分子标记研究中得到进一步证实[5,12,21]。这些结果充分表明,广东木荷资源蕴含丰富的遗传变异,遗传改良潜力巨大,选育生长与形质性状优良的遗传材料是可行的。
(3)木荷基因型主要由3组关键因素决定,即6~11 a生长量、3 a生长量与11 a干形、11 a形质性状,它们解释了总变异中的68.48%。55个基因型分布在18个聚类中,聚类间木荷表型差异巨大,聚类内表型近似,这可能与木荷资源的自然生境有关[3]。同一聚类中相邻家系大多来自相同的地理区域或临近区域,具有近似的自然气候,易发生基因交流[22]。不同聚类的木荷多来自不同地理生境,彼此间存在自然地理隔离,常进化产生丰富的遗传变异[1]。
(4)本研究阐述了木荷55个基因型的表型性状特征,并据此进行了类型划分。鉴于表型性状是植物基因型与环境效应的综合体现,植物可通过改变基因表达、调控、个体发育等中间环节响应周边环境的变化[13]。因此有必要探明木荷表型变异产生的机制机理。这一方面需深入研究木荷基因型与环境的互作机制,另一方面需从细胞学、生理生化以及分子生物学等层次探索木荷遗传变异规律及其产生机制,同时探讨木荷表现型与基因型分析结果的吻合程度,为育种研究筛选适宜的亲本材料[12]。
[1]Pyakurel A, Wang J R.Leaf morphological variation among paper birch (Betula papyrifera Marsh.) genotypes across Canada.Open Journal of Ecology[J].2013, 3: 284-295.
[2]Khadivi-Khub A, Ebrahimi A, Mohammadi A, et al.Characterization and selection of walnut (Juglans regia L.) genotypes from seedling origin trees.Tree Genetics and Genomes, 2015, 11: 54.
[3]倪 健.中国木荷及木荷林的地理分布与气候的关系[J].植物资源与环境学报, 1996, 5(3): 28-34.
[4]周志春, 范辉华, 金国庆, 等.木荷地理遗传变异和优良种源初选[J].林业科学研究, 2006, 19(6): 718-724.
[5]金则新, 李钧敏, 蔡琰琳.不同海拔高度木荷种群遗传多样性的ISSR分析.生态学杂志[J].2007, 26(8): 1143-1147.
[6]王秀花, 马雪红, 金国庆, 等.木荷天然林分个体类型及材性性状变异[J].林业科学, 2011, 47(3): 133-139.
[7]张 萍, 周志春, 金国庆, 等.木荷种源遗传多样性和种源区初步划分[J].林业科学, 2006, 42(2): 38-42.
[8]林 磊, 周志春, 范辉华, 等.木荷优树子代苗期生长遗传和变异研究[J].林业科学研究, 2009, 22(2): 155-160.
[9]王秀花, 陈柳英, 马丽珍, 等.7年生木荷生长和木材基本密度地理遗传变异及种源选择[J].林业科学研究, 2011, 24(3):307-313.
[10]楚秀丽, 王 艺, 金国庆, 等.不同生境、初植密度及林龄木荷人工林生长、材性变异及林分分化[J].林业科学, 2014,50(6): 152-159.
[11]辛娜娜, 张 蕊, 范辉华, 等.5年生木荷生长和形质性状的家系变异和选择[J].林业科学研究, 2014, 27(3): 316-322.
[12]辛娜娜, 张 蕊, 徐肇友, 等.木荷1代育种群体遗传多样性分析[J].林业科学研究, 2015, 28(3): 332-338.
[13]闫莉萍, 张方秋, 潘 文, 等.广东省木荷群体叶片表型多样性研究[J].广东林业科技, 2013, 29(5): 35-38.
[14]Ledig F T.Analysis of methods for the selection of trees from wild stands[J].Forest Science, 1974, 20: 2-16.
[15]张 谦, 曾令海, 蔡燕灵, 等.樟树自由授粉家系生长与形质性状的遗传分析[J].中南林业科技大学学报, 2014, 34(1):1-6.
[16]张 谦, 曾令海, 何波祥, 等.马尾松自由授粉家系产脂力的年度变化及遗传分析[J].林业科学, 2013, 49(1): 58-62.
[17]Zeng L H, He B X, Lian H M, et al.Age trends in genetic parameters for growth and resin-yielding capacity in masson pine[J].Silvae Genetica, 2013, 62: 7-18.
[18]Iezzoni A F, Pritts M P.Applications of principal components analysis to horticultural research[J].HortScience, 1991, 26: 334-338.
[19]陈 欢, 曹承富, 张存岭, 等.基于主成分-聚类分析评价长期施肥对砂姜黑土肥力的影响[J].土壤学报, 2014, 51(3):609-617.
[20]Gitonga V W, Koning-Boucoiran C F S, Verlinden K, et al.Genetic variation, heritability and genotype by environment interaction of morphological traits in a tetraploid rose population[J].BMC Genetics, 15:146.
[21]杨汉波, 张 蕊, 周志春.木荷种子园的遗传多样性和交配系统[J].林业科学, 2016, 52(12): 66-73.
[22]Sefc K M, Lopez M S, Lefort F, et al.Microsatellites variability in grapevine cultivars from different European regions and evaluation of assignment testing to assess the geographic origin of cultivars[J].Theoretical and Applied Genetics, 2000, 100:498-505.
