三江源地区高寒草地地上生物量时空动态变化
2018-05-26杨淑霞冯琦胜孟宝平高金龙梁天刚
杨淑霞,冯琦胜,孟宝平,高金龙,葛 静,梁天刚
(兰州大学草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州 730020)
草地是覆盖陆地表面最大的自然景观之一,约占陆地面积的1/5[1],是陆地生态系统最重要的组成部分之一。草地生物量可以判断草地的生长状况和生产潜力,草地生物量及其动态变化的准确估算在全球碳循环方面具有重要意义[2-3]。
三江源区作为长江、黄河和澜沧江的发源地,素有“中华水塔”之称。是我国重要的水源涵养和生态功能保护区,三江源区具有特殊的地理位置,丰富的自然资源以及显著的生态功能,三江源区成为青藏高原重要的自然保护区[4-5]。然而,三江源区具有海拔高、自然条件严酷等特点,致使该区生态系统极其脆弱。除此之外,近年来随着气候不断变暖及人类干扰活动的不断增强,三江源区生态系统发生了很大的变化。为了及时准确地获取该区域草地生物量信息,开展三江源区草地地上生物量动态变化的研究,可以为该区草地资源的可持续利用和生态恢复提供科学依据。
传统的生物量实地测量方法费时费力、成本高、效率低,且难以在大范围地区开展草地调查工作。遥感技术的发展,为草地植被信息的研究提供了新的方法[6]。遥感手段具有重复周期短、覆盖范围广、数据源丰富、节约成本等优势,得到了广泛的应用[7]。许多学者运用3S技术对草地地上生物量进行了大量研究,以植物叶面在可见光波段有强吸收特性而在红外波段有强反射特性为基础[8-9],利用植被指数,如NDVI (normalized difference vegetation index)等,结合地面样点的调查数据和遥感图像处理系统,构建地上生物量的统计模型,开展地上生物量动态监测研究。遥感监测方法已在世界各地,尤其是偏远地区被广泛应用于草地地上生物量的动态监测和评价研究中[10]。如姚兴成等[11]以云南省为研究区,结合2012-2014年草地生物量野外调查数据以及相应时期的MODIS(moderate-resolution imaging spectroradiometer)遥感数据,建立草地地上生物量的遥感监测模型。于惠等[6]基于MODIS SWIR(short-wave infrared)数据及地面实测样方数据,筛选出干旱区草地地上生物量(above ground biomass,AGB)的敏感性指标,构建了草地AGB反演模型。除此之外,也有学者对高寒草甸生物量动态变化与环境因子之间的关系进行探究[12-14]。刘艳书等[15]分析了三江源区不同剥蚀退化高寒草甸生物群落生物量的变化特征;李凤霞等[16]探索了放牧强度对三江源典型高寒生物量的影响,发现封育区的地上生物量和地下生物量最高,随着放牧强度的增加,草地地上生物量和地下生物量均呈减少趋势;曾纳等[17]结合遥感植被指数、海拔、气象观测资料构建了BP神经网络模型,对2001-2010年三江源区的草地AGB进行估算,并分析草地地上生物量动态变化。以上的研究采用不同模拟方法估算了草地地上生物量,并且探讨了影响生物量变化的相关因子,但是对草地地上生物量的精确反演一直是草地遥感监测研究的难点[18]。基于以上原因,本研究利用三江源区草地地上生物量实测值,结合同时段的遥感数据,采用BP-ANN(back propagation artificial neural network)的方法构建草地地上生物量估算模型,分别从草地类型和流域方面分析草地地上生物量的年际和月际变化,以揭示该区域草地地上生物量的动态变化特征。
1 材料与方法
1.1 研究区概况
三江源区地处青海省南部,其地理位置31°38′-36°20′ N,89°31′-102°14′ E,三江源区是青海省的主要牧区。该区是我国黄河、长江、澜沧江三大水系发源地,素有“中华水塔”之称。
三江源区土地总面积36.3万km2,约占青海省土地总面积的50.3%。该区隶属青海省南部的3个自治州,三江源区天然草原面积为2 098.97万hm-2,占三江源区土地总面积的59%,其中可利用草地面积1 787.44万hm-2。草地类型主要由高寒草甸类(alpine meadow steppe)和高寒干草原类(alpine steppe)组成,高寒草甸面积约占草地占面积的76%,高寒干草原面积约占草地占面积的23%,土壤主要以高山沼泽化草甸为主(图1)。其平均海拔在4 000 m以上,年平均气温介于-5.6~4.9 ℃,大部分地区低于1 000 ℃,年降水量介于391.7~764 mm,主要集中在6-9月。夏季短暂,牧草生长期短,植物长势低矮,产草量低,冷季长达7~8个月,为典型的高原大陆性气候[19]。
1.2 研究数据
1.2.1地面实测数据 本研究所用的草地外业观测数据包括:1) 2003-2014年青海省气象局在三江源区的15个牧业气象站的监测数据[19](图1a);2)2010-2014年青海省草原站草地观测数据[18](图1a);3)2015年本项研究人员在三江源区开展的草地调查数据(图1b)。

图1 2003-2015年三江源区采样点(a)、草地类型(b)和流域(c)分布图Fig. 1 The sampling sites (a), grassland types (b) and watersheds(c) in the Three River Headwaters Region (TRHR) from 2003 to 2015
在以上调查中,样区面积设置为(3~5) km×(1~2) km,在此范围内设置监测样方,在植被群落均一性较好的样地内设置样方,其大小为0.5 m×0.5 m,在植被均一性较差且分布不均匀的地区设置样方,其大小为1.0 m×1.0 m。每年在草地生长盛草期(7-8月)开展草地监测,主要记录样地和样方的经纬度、海拔、坡度、坡向、草地类型、草地植被盖度、草层高度、地上生物量鲜重,以及在65 ℃烘箱烘干48 h后测量的干物质产量等特征数据。
1.2.2地形数据 本研究使用DEM数据的空间分辨率为90 m,版本为V004的SRTM(Shuttle Radar Topography Mission)影像,从国际农业研究磋商小组空间信息联盟(The Consultative Group on International Agricultural Research Consortium for Spatial Information,CGIAR-CSI)的网站(http://srtm.csi.cgiar.org/)下载。数据格式为Geo-Tiff。为了和已有数据相匹配,将经过拼接的数字高程数据进行转投影和重采样处理,投影类型定义为WGS84,栅格单元重采样至500 m。为了开展后续的统计分析,使用ArcGIS软件提取与地面实测样点相对应像元的经度、纬度。
1.2.3MODIS植被指数数据 本研究使用了MODIS逐日地表反射率产品(MOD09GA),数据格式为EOS-HDF,空间分辨率为500 m。对2001-2016年三江源区每年5-10月逐日MOD09GA反射率数据进行投影转换及拼接等处理,得到MOD09GA 1-7通道的逐日反射率数据。结合质量控制文件,采用最大值合成方法计算得到研究区16年间逐月NDVI植被指数图像。
1.3 研究方法
1.3.1基于ANN的草地地上生物量模型构建 本研究选取草地盖度、高度、地理位置(经纬度)和NDVI作为输入参数,基于BP-ANN方法,构建了草地地上生物量反演模型,构建过程包括如下步骤:1)数据归一化。将所有的输入数据归一化到0~1,归一化方法为(x-xmin)/(xmax-xmin),其中x为某一参数的原始值,xmin和xmax分别为该参数的最大和最小值。2)将输入数据按75%、15%、15%的比例随机分成训练数据集(training)、验证数据集(validation)和测试数据集(testing)三部分。选择Levenberg-Marquardt函数作为ANN训练函数,平均误差(MSE)作为训练评价指标,当验证样本的平均误差增加时训练停止,并计算目标数据与输出数据之间的相关关系[20],通过多次训练,根据训练结果确定适当的隐藏层节点数据。
1.3.2草地地上生物量监测模型精度分析方法 以实测值与相应的模拟值之间的决定系数(R2)和均方根误差(root mean square error,RMSE)对模型进行精度评价。其中,R2取值范围0~1,其值越接近1,模型精度越高,可信度也就越高;RMSE用来衡量预测值和实测值之间的偏差,其值越小,表示构建的草地地上生物量模型拟合效果越好。RMSE和决定系数的计算公式如下:
(1)
(2)
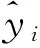
1.3.3三江源区高寒草地AGB变化趋势分析 采用Theil-Sen Median趋势分析和Mann-Kendall 趋势检验,研究三江源区地上生物量的时空变化动态[21]。
1)Theil-Sen median趋势分析。Theil-Sen Median趋势分析和Mann-Kendall检验结合分析,已成为长时间序列数据趋势判断的重要方法,并且已经成功应用到植被长时间序列分析中[22-25]。该方法的优点是不需要数据服从一定的分布,对数据误差具有较强的抵抗能力,对于显著水平的检验具有较为坚实的统计学理论基础,使得结果较为科学和可信[22]。其中,Theil-Sen Median 趋势分析是一种稳健的非参数统计的趋势计算方法,可以减少数据异常值的影响。Theil-Sen Median 趋势计算n(n-1)/2个数据组合的斜率的中位数,其计算公式[26-28]为:
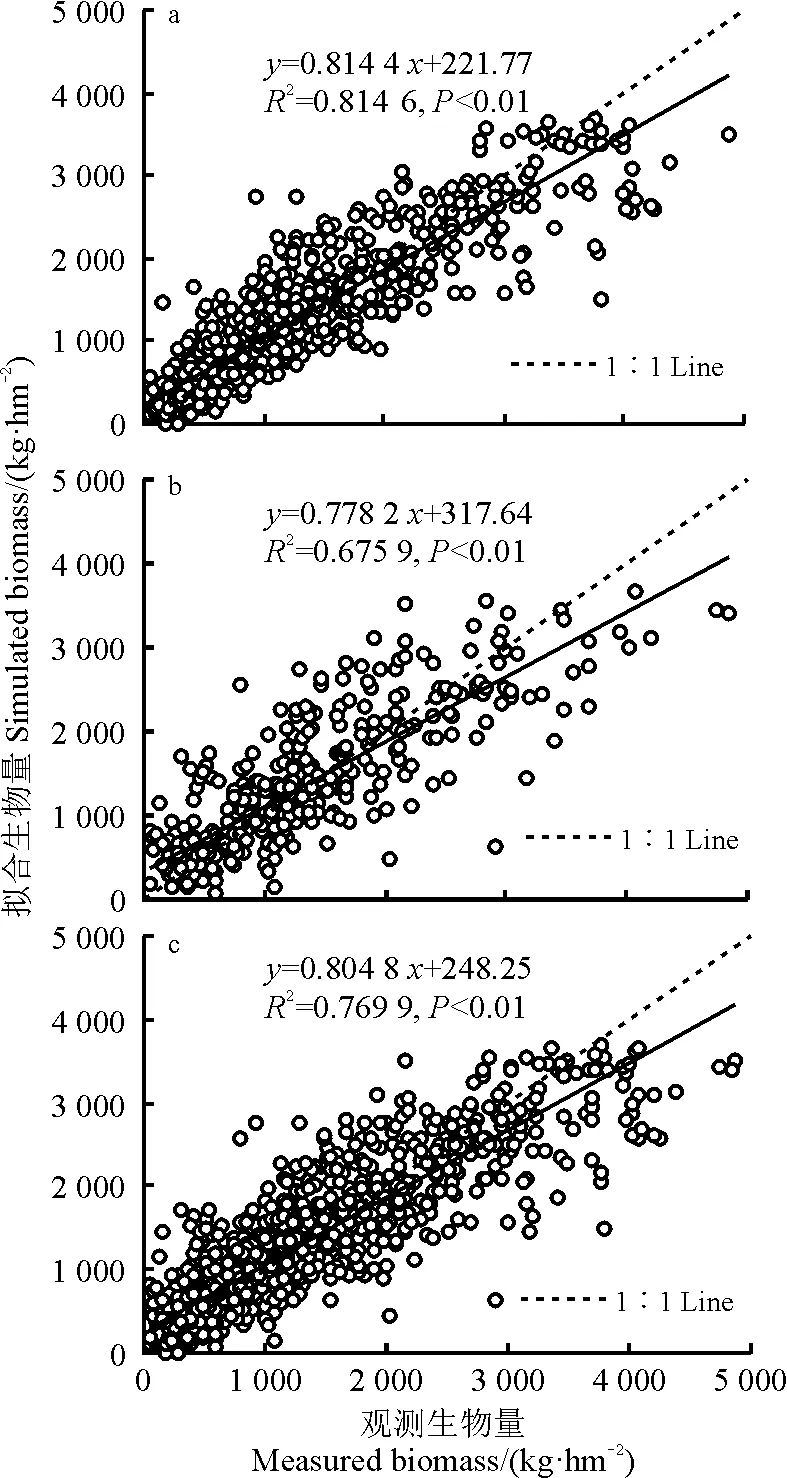
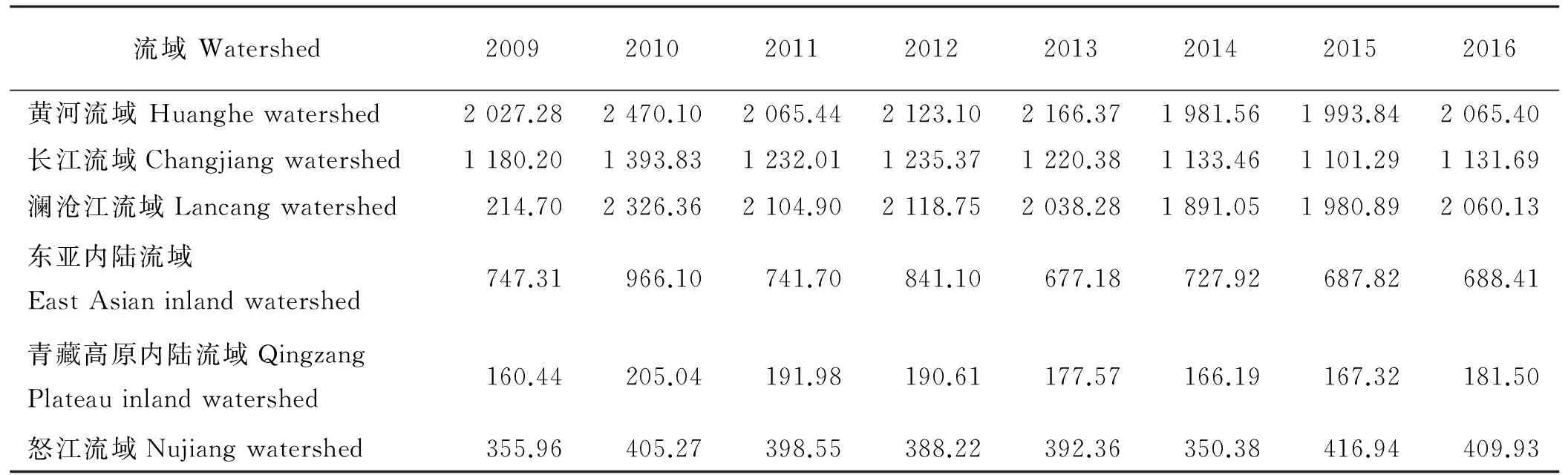
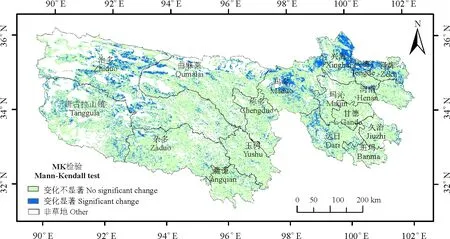
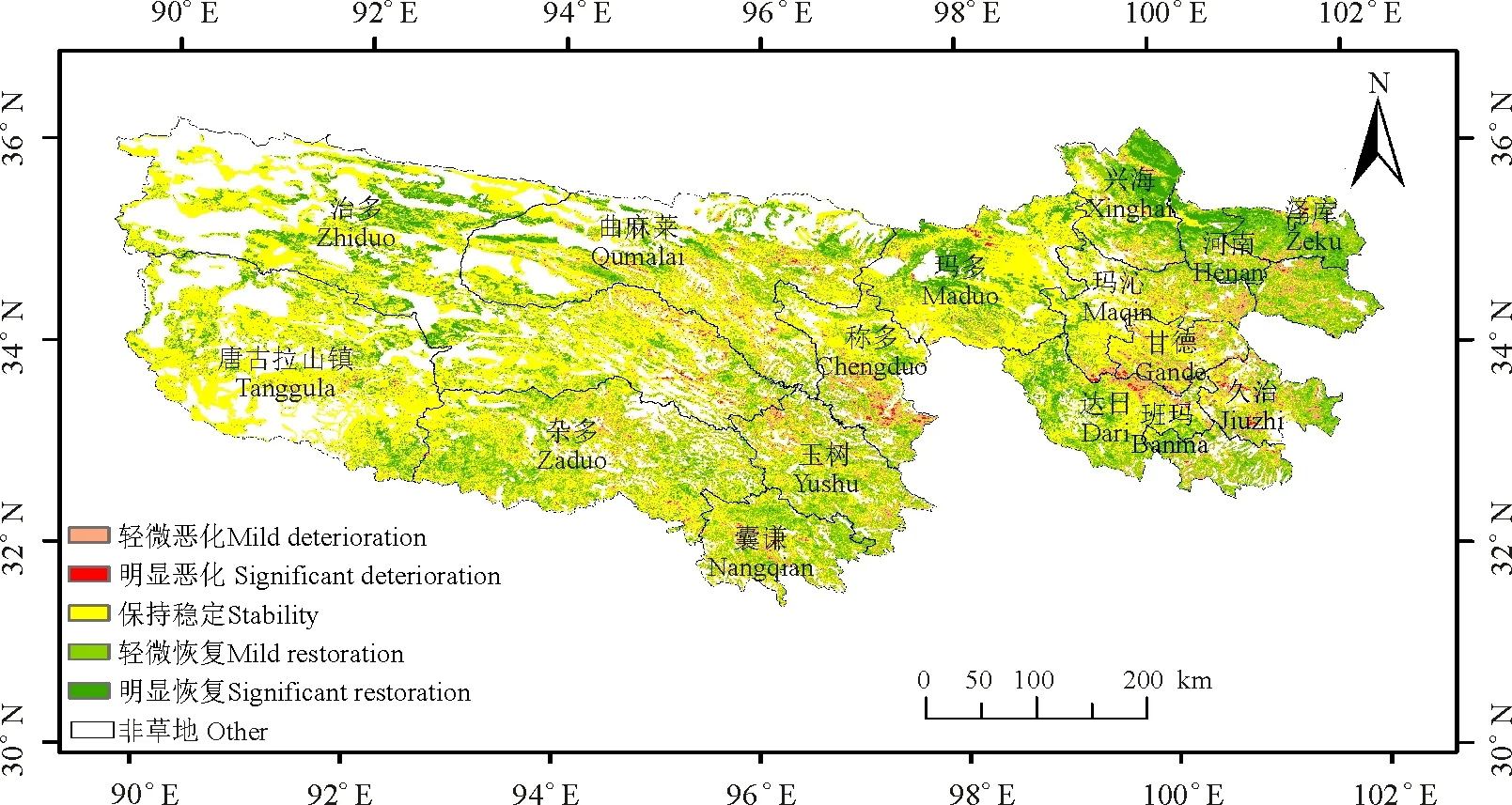
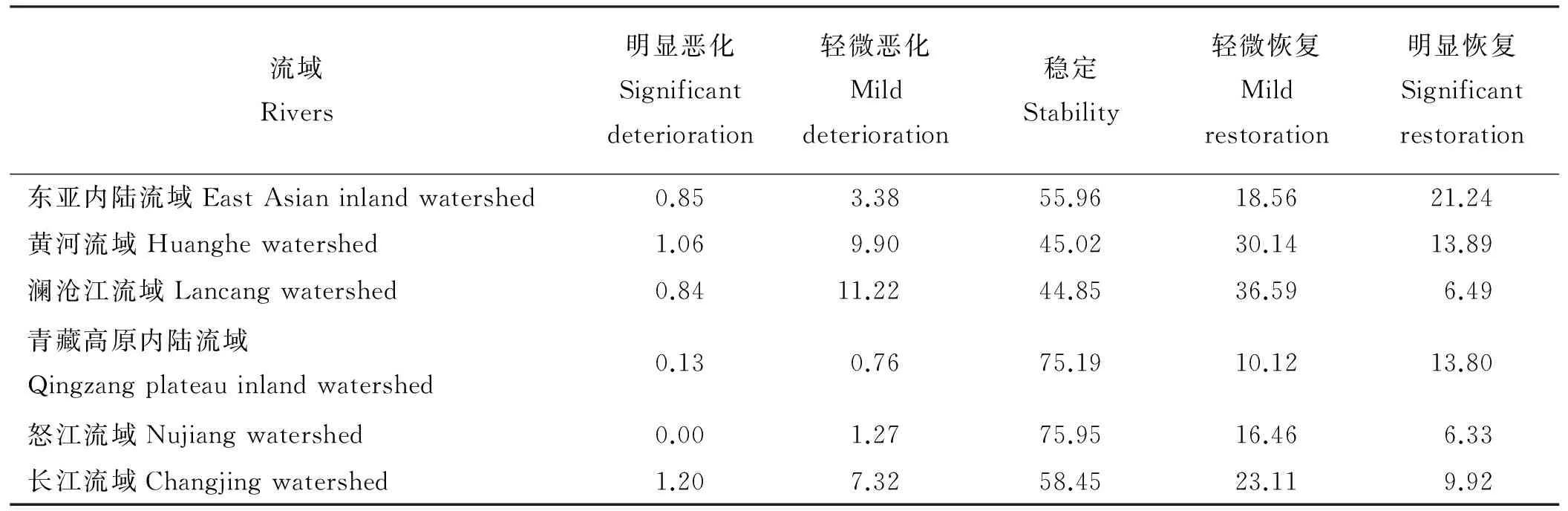
2001≤i (3) 式中:AGBj和AGBi分别表示j年和i年的AGB值。Median为符号涵数。当SAGB>0时,表明AGB呈增长趋势,当SAGB<0时,表明AGB呈恶化趋势。 2)Mann-kendall检验。Mann-Kendall是一种非参数统计检验方法,用来判断趋势的显著性,它无需样本服从一定的分布,也不受少数异常值的干扰[29-30]。其计算公式如下: (4) (5) (6) S(S)=n(n-1)(2n+5)/18。 (7) 式中:AGBi和AGBj分别表示i年和j年的AGB值,n表示时间序列的长度;sgn是符号函数;统计量Z的取值范围为(-∞,+∞)。在给定显著性水平α下,当|Z|>u1-α/2时,表示研究序列在α水平上存在显著的变化。一般取α=0.05,本文判断在0.05置信水平上AGB时间序列变化趋势的显著性。 为了探索神经网络方法在高寒草地生物量反演中的作用,本研究采用BP ANN方法,选取与AGB关系密切且具有一定独立性的草地生物物理指标:盖度(C),地理位置:经度(X)、纬度(Y),及MODIS的NDVI(N)4变量作为自变量,构建基于BP ANN的AGB反演模型(CXYN的模型)。该模型的R2=0.769 9,RMSE=429.06 kg·hm-2(图2)。模型可以很好地拟合研究区草地地上生物量的变化趋势,且优于单因素模型[19]。 16年间各草地类型的生物量总体表现出从5月份开始上升,7月和8月达到峰值,9月开始下降的趋势(图3)。各草地类型盛草期的生物量介于40.59~2 600.04 kg·hm-2。其中,山地草甸类的生物量高于其他草地类型,7月和8月的生物量分别为2 657.30和2 600.04 kg·hm-2;其次为高寒草甸类,7-8月生物量介于1 489.43~1 496.09 kg·hm-2;温性草原类7月的生物量最大,为1 339.09 kg·hm-2;高寒草原类4-10月的生物量介于85.88~397.13 kg·hm-2;沼泽类8月的生物量最高,为483.80 kg·hm-2;高寒荒漠类的生物量最低,7月和8月的生物量均低于50 kg·hm-2。 16年间各流域生物量月际变化趋势与草地类型和气候区的变化基本一致(图4),即7月和8月的草地生物量高于其他月份。从各个流域看,黄河流域的生物量值均高于其他流域,生物量介于354.86~1 819.67 kg·hm-2,4-10月平均值为960.44 kg·hm-2,5月的生物量最低,7月的生物量最高;澜沧江流域的生物量略低于黄河流域,6-9月的生物量介于751.98~1 032.63 kg·hm-2;长江流域7月和8月的生物量高于其他月份,分别为978.80和1 006.39 kg·hm-2;东亚内陆流域生物量介于118.06~596.77 kg·hm-2, 7月和8月的生物量值较接近,分别为588.8和596.77 kg·hm-2;怒江流域和青藏高原内陆流域生物量的平均值分别为128.62和52.63 kg·hm-2,均低于其他流域。总体而言,各流域生物量从高到低依次为黄河流域、澜沧江流域、长江流域、东亚内陆流域、怒江流域和青藏高原内陆流域,不同流域的生物量存在差异。这种差异是由于不同流域所处地域范围不同,气候环境差异较大导致的。研究区内处于东部和南部的流域生物量高于西部和北部的流域。同时,虽然长江流域分布在三江源区的东部、南部和中部地区,范围最大,但其生物量要低于黄河流域和澜沧江流域。 图2 基于CXYN模型的人工神经网络模型模拟结果 Fig. 2 The ANN simulated results based on CXYN model a,训练集;b,测试集;c,全集。 a, Training set; b, Test set; c, Total. 图3 三江源区2001-2016年生长季不同草地类型草地生物量变化Fig. 3 The different grassland type biomass change at growing season in the Three-River Headwaters Region from 2001 to 2016 图4 2001-2016年三江源区不同流域生物量月际变化Fig. 4 The monthly variation of different river basin biomass in the Three-River Headwaters Region from 2001 to 2016 2016年1-12月地上生物量的空间变化趋势基本一致,即由西北向东南逐渐增加的趋势(图5)。1-5月东南部的生物量小于1 000 kg·hm-2。中部及西部地区的生物量主要集中在300~500 kg·hm-2及小于300 kg·hm-2。从6月开始,生物量从东南向西北部逐渐呈增加趋势,其中6月东南部的部分区域生物量最大值达3 000 kg·hm-2,中部部分区域的生物量最大值达2 000 kg·hm-2。7月和8月东南部的一些区域生物量最大值在3 000 kg·hm-2以上,中部区域的生物量较前5个月有所增加,生物量主要介于500~800 kg·hm-2以及800~1 000 kg·hm-2,且西北部的生物量值主要介于300~500 kg·hm-2以及500~800 kg·hm-2,只有少数区域的生物量值小于300 kg·hm-2。 基于构建的生物量反演模型,对三江源区草地地上生物量的动态变化进行分析,按草地类型、流域分别统计分析了三江源区年最大生物量16年来的变化状况,主要结果如下: 7种草地类型的生物量在16年间均呈现不同程度的上升趋势(图6)。由此表明,16年间三江源区草地地上生物量均呈现增加趋势,生态环境整体趋于好转。从不同草地类型来看,各草地类型的年生物量从高到低依次为山地草甸类、高寒草甸类、温性草原类、温性荒漠草原类、沼泽类、高寒草原类和高寒荒漠类。16年间,温性草原类和温性荒漠草原类草地的生物量有明显的增加趋势,如温性草原类2001年生物量为985.37 kg·hm-2,到2016年增加到1 479.17 kg·hm-2,平均每年增加约30.86 kg·hm-2。山地草甸类、高寒草甸类和沼泽类草地的生物量在16年间均有较大幅度的增加,而高寒荒漠类草地的生物量在16年间基本保持不变。 2010年各个流域的生物量值最大(表1)。黄河流域生物量值介于1 767.95~2 470.10 kg·hm-2,其平均值为2 014.30 kg·hm-2;澜沧江流域生物量平均值为1 969.43 kg·hm-2,其中2010年的生物量值最高,为2 326.36 kg·hm-2,2007年的生物量值最低,为1 690.18 kg·hm-2;长江流域和东亚内陆流域生物量的平均值分别为1 145.63和689.10 kg·hm-2;怒江流域和青藏高原内陆流域的生物量平均值分别为363.30和156.19 kg·hm-2。16年间6个流域的生物量均呈现出增加趋势,其中黄河流域生物量增加幅度最大,平均每年增加15.57 kg·hm-2,澜沧江流域和东亚内陆流域次之,平均每年分别增加12.40和11.49 kg·hm-2;青藏高原内陆流域和怒江流域最低,平均每年增加4.33 kg·hm-2。 图5 三江源区草地地上生物量月际变化动态Fig. 5 The dynamic of monthly variation of the grassland AGB in the Three-River Headwaters Region 图6 三江源区不同草地类型地上生物量年际变化动态Fig. 6 The dynamic of yearly variation of different grassland type AGB in the Three-River Headwaters Region 三江源区草地地上生物量呈现出从西北向东南逐渐增加的变化趋势,这与该区域的降水、气温、植被和草地类型等因素密切相关(图7)。东南部生物量主要集中在2 000~3 000 kg·hm-2,部分区域的生物量大于3 000 kg·hm-2,中部生物量主要集中在800~1 000和1 000~2 000 kg·hm-2,西北部生物量主要介于300~500 kg·hm-2,少部分区域的生物量小于300 kg·hm-2。 总体而言,三江源区16年间年最大生物量介于1 272.24~1 739.56 kg·hm-2,其中2003年生物量最低,2010年最高。 16年来三江源区年生物量主要以稳定和恢复为主(图8),恶化区域主要集中分布在三江源区东部和中部的部分区域。在三江源区西北部的部分区域,有少数区域的草地出现恶化趋势,大部分区域仍以稳定和恢复趋势为主。基于Mann-Kendall Test方法的2001-2016年三江源区草地地上生物量的变化趋势显著性检验得出,地上生物量的变化趋势从东北部到西北部主要以变化不显著为主,变化显著的主要分布在东部和西北部的部分区域(图9)。为了揭示草地地上生物量的变化趋势及其趋势检验,将生物量变化趋势结果与Mann-Kendall Test结果进行叠加,得到变化趋势与趋势检验的空间分布格局。将这种趋势检验分为5种情形,即明显变化、轻微变化、保持稳定、轻微恢复、明显恢复。 统计各情景的草地面积结果表明(表3、图10),16年间,三江源区各类草地生物量的变化趋势主要以稳定和恢复为主。分析不同草地类型生物量的变化趋势表明,高寒荒漠类的稳定区域所占比例为86.88%,明显恶化和轻微恶化所占比例较小,分别为0.02%和0.79%;高寒草原类有63.32%的草地呈稳定状态,34.55%的草地呈恢复状态;高寒草甸类51.83%的区域呈稳定状态,28.26%的草地呈轻微恢复趋势,8.81%的草地呈明显恢复趋势。48.62%的山地草甸类呈轻微恢复趋势,30.23%的草地保持稳定状态,同时有11.62%的草地出现轻微恶化;沼泽类有64.11%的草地呈恢复趋势,26.46%的草地保持稳定,9.43%的草地出现恶化现象;温性草原类和温性荒漠草原类主要以恢复为主,分别占86.81%和81.62%,而恶化趋势所占比例均较小。总体而言,三江源区16年间53.74%的草地保持稳定,37.31%的草地表现出不同程度的恢复趋势,有8.95%的草地呈现恶化趋势。综上,可以认为三江源区草地生物量表现出整体稳定,局部恶化的趋势。 表1 2001-2008年三江源区不同流域生物量年际变化Table 1 The yearly variation of different watershed biomass in the Three-River Headwaters Region from 2001 to 2008 kg·hm-2 表2 2009-2016年三江源区不同流域生物量年际变化Table 2 The yearly variation of different watershed biomass in the Three-River Headwaters Region from 2009 to 2016 kg·hm-2 图7 三江源区2001-2016年草地地上生物量空间分布图Fig. 7 The Spatial distribution map of the grassland AGB in the Three-River Headwaters Region from 2001 to 2016 图8 2001-2016年三江源区生物量的变化趋势Fig. 8 The change trend of the grassland AGB in the Three-River Headwaters Region from 2001 to 2016 从2001-2016年三江源区各个流域草地地上生物量的变化趋势可以看出,不同流域均以稳定为主,怒江流域草地稳定趋势所占比例最大,为75.95%,未出现明显恶化;其次为青藏高原内陆流域,75.19%的草地保持稳定;长江流域的草地有8.52%的草地出现恶化,33.03%的草地呈恢复趋势;黄河流域和澜沧江流域草地稳定趋势所占比例差异不大,但是澜沧江流域有12.06%的草地出现恶化,其中轻微恶化的比例为1.22%,相比而言,轻微恢复的比例较大,为36.59%;东亚内陆流域有55.96%的草地保持稳定,明显恢复和轻微恢复所占比例分别为21.24%和18.56%(表4)。 图9 2001-2016年三江源区生物量的变化趋势检验Fig. 9 The change trend test of the grassland AGB in the Three-River Headwaters Region from 2001 to 2016 表3 2001-2016年三江源区不同草地类型生物量变化趋势Table 3 The change trend of different grassland types AGB in the Three-River Headwaters Region from 2001 to 2016 % 图10 2001-2016年三江源区生物量变化趋势分布图Fig. 10 The distribution of the grassland AGB change trend in the Three-River Headwaters Region from 2001 to 2016 表4 2001-2016年三江源区不同流域生物量变化趋势Table 4 The change trend of different watershed AGB in the Three-River Headwaters Region from 2001 to 2016 % 草地地上生物量遥感估算模型精度受到以下4个方面的影响:1)地形因子。三江源区特殊的地理位置,难以在整个研究区进行大范围且均匀的实地进行调查,这在一定程度上限制了采样数据的空间分布,如本研究的采样数据多集中在研究区的东部和中部地区,而西部地区几乎没有地面调查数据,这使得构建的草地地上生物量遥感估算模型在西部地区的代表性较低;2)地面实测数据源于不同采集人员,其采样时间不同,采样数据的标准不同,导致采样数据的质量存在一定的误差;3)三江源区特殊的地形地貌和草地资源空间异质性的影响,采样数据的空间代表性受到一定限制。MODIS的空间分辨率较低,本研究所用的MOD09GA空间分辨率为500 m,MODIS比较低的空间分辨率导致其像元与地面观测样点在空间范围上存在较大误差,像元普遍存在混合地物光谱问题,另一方面,受采样数据限制,并未针对不同草地类型分别构建模型,由此忽略了草地类型在空间分布及生物物理特征的差异;4)由于复杂的地形条件,坡度坡向等地形因子会对植被反射率有一定的影响,特别是在三江源地区,草地类型主要以高寒草甸为主,但是某些地区的草地受自然条件和人为因素的影响,草地呈恶化趋势,草地稀疏,出现裸斑,影响到草地地上生物量模型的精度。 BP神经网络因其对多变量信息的综合能力及对非线性问题的拟合等方面所具有的无可比拟的优势[26],已被广泛应用于基于高光谱的叶面积指数、地上生物量等参数的估算或反演研究领域。利用人工神经网络对草地地上生物量进行估算的研究结果表明,人工神经网络模型可以充分反应草地地上生物量与地理位置、生物物理指标之间的非线性关系,模拟精度优于其他传统的模型。 然而,在估算过程中仍然存在一些问题,如依赖学习样本的数量和质量的代表性,样本容量越大,模型精度越高。样本容量较小时,可能对某一组数据表现出很好的结果,但是对另外一组数据恰好相反。另一方面,ANN的最佳隐藏层和每层节点的数目取决于反复试验,如何确定合理的隐藏层节点数是草地地上生物量反演的关键问题。 虽然神经网络能够解决多因素线性及非线性问题,训练的模型精度比统计模型高,但是对于神经网络训练过程及构建的模型方程式却看不到。因此,从理论角度很难对其结果进行解释。 本研究以三江源区为研究区,主要研究了高寒草地地上生物量遥感反演模型构建、生物量时空变化特征分析等方面的工作。利用MODIS数据与野外实测资料,构建了基于神经网络方法的三江源区草地生物量反演的高精度模型,实现了对2001-2016年三江源区草地AGB反演分析;并探讨分析了不同草地类型和流域的草地AGB时空变化特征; 1)采用BP-ANN的方法构建的基于盖度、经纬度和NDVI的神经网络模型适用于天然草地生物量变化监测研究。 2)各草地类型盛草期的生物量介于40.59~2 600.04 kg·hm-2。山地草甸类的生物量高于其他草地类型,高寒荒漠类的生物量最低。7种草地类型的生物量在16年间均呈现不同程度的上升趋势。 3)三江源区黄河流域的生物量高于其他流域,4-10月生物量值介于354.86~1 819.67 kg·hm-2;各流域4-10月生物量从高到低依次为黄河流域、澜沧江、长江流域、东亚内陆流域、怒江流域和青藏高原内陆流域,不同流域的生物量存在显著的差异。 4)基于Theil-Sen Median和Mann-Kendall方法的趋势分析结果表明,16年间,三江源区草地生物量的变化趋势主要以稳定和恢复为主。 参考文献References: [1] Scurlock J M O,Hall D O. The global carbon sink:A grassland perspective.Global Change Biology,1998,4(2):229-233. [2] Mao D H,Wang Z M,Li L,Ma W.Spatiotemporal dynamics of grassland aboveground net primary productivity and its association with climatic pattern and changes in Northern China.Ecological Indicators,2014,41(6):40-48. [3] Toan T L,Quegan S,Davidson M W J,Balzter H,Paillou P,Papathanassion K,Plummer S,Rocca F,Saatchi S,Shugart H,Ulander L.The biomass mission: Mapping global forest biomass to better understand the terrestrial carbon cycle.Remote Sensing of Environment,2011,115(11):2850-2860. [4] Tong L,Xu X L,Fu Y,Li S.Wetland changes and their responses to climate change in the “Three-River Headwaters” region of China since the 1990s.Energies,2014,7(4):2515-2534. [5] Liu X,Zhang J,Zhu X,Pan Y,Liu Y,Zhang D,Lin Z.Spatiotemporal changes in vegetation coverage and its driving factors in the Three-River Headwaters region during 2000-2011.Journal of Geographical Sciences,2014,24(2):288-302. [6] 于惠,吴玉锋,金毅,张峰.基于MODIS SWIR数据的干旱区草地地上生物量反演及时空变化研究.遥感技术与应用,2017,32(3):524-530. Yu H,Wu Y F,Jin Y,Zhang F.An approach for monitoring the aboveground biomass of grassland in arid areas based on MODIS SWIR bands.Remote Sensing Technology and Application,2017,32(3):524-530.(in Chinese) [7] 张静宇,史击天,张群.遥感技术在北京市农业统计中的应用现状.宁夏农林科技,2012,53(4):110-112. Zhang J Y,Shi J T,Zhang Q.Application of remote sensing technology in agriculture-statistics in Beijing.Ningxia Journal of Agriculture and Forestry Science and Technology,2012,53(4):110-112.(in Chinese) [8] Craine J M,Nippert J B,Elmore A J,Skibbe A M,Hutchinson S L,Brunsell N A.Timing of climate variability and grassland prodctivity.PNAS,2012,109(9):3401-3405. [9] Eisfelder C,Kuenzer C,Dech S.Derivation of biomass information for semi-arid areas using remote-sensing data.International Journal of Remote Sensing,2012,33(9):2937-2984. [10] Lu D.The potential and challenge of remote sensing:Based biomass estimation.International Journal of Remote Sensing,2006,27(7):1297-1328. [11] 姚兴成,曲恬甜,常文静,易俊,李永进,孙振中,曾辉.基于MODIS数据和植被特征估算草地生物量.中国生态农业学报,2017,25(4):530-541. Yao X C,Qu T T,Chang W J,Yi J,Li Y J,Sun Z Z,Zeng H.Estimating of grassland biomass using MODIS data and plant community characteristics.Chinese Journal of Eco-Agriculture,2017,25(4):530-541.(in Chinese) [12] 付天新,刘正军,闫浩文.基于MapReduce模型的生物量并行反演方法.干旱区资源与环境.2013,27(1):130-136. Fu T X,Liu Z J,Yan H W.Remote sensing retrieval method for biomass based on MapReduce parallel model.Journal of Arid Land Resources and Environment,2013,27(1):130-136.(in Chinese) [13] 聂秀青,杨路存,李长斌,周国英.三江源地区高寒灌丛生物量空间分布格局.应用与环境生物学报,2016,22(4):538-545. Nie X Q,Yang L C,Li C B,Zhou G Y.Patterns of biomass partitioning across alpine shrubs in the Three-River Source Region. Chinese Journal of Applied and Environmental Biology,2016,22(4):538-545.(in Chinese) [14] 徐满厚,刘敏,翟大彤,薛娴,彭飞,尤全刚.青藏高原高寒草甸生物量动态变化及环境因子的关系:基于模拟增温实验.生态学报,2016,38(18):5759-5767. Xu M H,Liu M,Zhai D T,Xue X,Peng F,You Q G.Dynamic changes in biomass and its relationship with environmental factors in an alpine meadow on the Qinghai-Tibetan Plateau,based on simulated warming experiments.Acta Ecologica Sinica.2016,36(18):5759-5767.(in Chinese) [15] 刘艳书,樊江文,李愈哲,张良侠.三江源地区不同剥蚀退化高寒草甸群落生物量与多样性特征.草业学报,2014,23(3):1-7. Liu Y S,Fan J W,Li Y Z,Zhang L X.Plant community and diversity on alpine meadow steppe in the Three River Headwaters Region,Qinghai Province under different denudation levels.Acta Prataculturae Sinica,2014,23(3):1-7.(in Chinese) [16] 李凤霞,李晓东,周秉荣,祁栋林,王力,傅华.放牧强度对三江源典型高寒草甸生物量和土壤理化特征的影响.草业科学,2015,32(1):11-18. Li F X,Li X D,Zhou B R,Qi D L,Wang L,Fu H.Effects of grazing intensity on biomass and soil physical and climate characteristics in the source of Three Rivers.Pratacultural Science,2015,32(1):11-18.(in Chinese) [17] 曾纳,任小丽,何洪林,张黎,李攀,李志强,张林波.基于神经网络的三江源区草地地上生物量估算.环境科学研究,2017,30(1):59-66. Zeng N,Ren X L,He H L,Zhang L,Li P,Li Z Q,Zhang L B.Aboveground biomass of grassland in the Three-River Headwaters Region based on neural network.Research of Environmental Sciences,2017,30(1):59-66.(in Chinese) [18] 杨淑霞,张文娟,冯琦胜,孟宝平,高金龙,梁天刚.基于MODIS逐日地表反射率数据的青南地区草地生长状况遥感监测研究.草业学报,2016,25(8):14-26. Yang S X,Zhang W J,Feng Q S,Meng B P,Gao J L,Liang T G.Monitoring of grassland herbage accumulation by remote sensing using MODIS daily surface reflectance data in the Qingnan Region.Acta Prataculturae Sinica,2016,25(8):14-26.(in Chinese) [19] Liang T G,Yang S X,Feng Q S,Liu B K,Zhang R P,Huang X D,Xie H J.Multi-factor modeling of above-ground biomass in alpine grassland: A case study in the Three-River Headwaters Region,China.Remote Sensing of Environment,2016,186:164-172. [20] Ali I,Cawkwell F,Dwyer E,Barrett B,Green S.Satellite remote sensing of grasslands:From observation to management: Areview.Journal of Plant Ecology,2016,9(6):649-671. [21] 袁丽华,蒋卫国,申文明,刘颖慧,王文杰,陶亮亮,郑华,刘孝富.2000-2010年黄河流域植被覆盖的时空变化.生态学报,2013,33(24):7798-7806. Yuan L H,Jiang W G,Shen W M,Liu Y H,Wang W J,Tao L L,Zheng H,Liu X F.The spatio-temporal variation of vegetation cover in the Yellow River Basin from 2000 to 2010.Acta Ecologica Sinica,2013,33(24):7798-7806.(in Chinese) [22] 蔡博峰,于嵘.基于遥感的植被长时序趋势特征研究进展及评价.遥感学报,2009,13(6):1170-1186. Cai B F,Yu R.Advance and evaluation in the long time series vegetation trends research based on remote sensing.Journal of Remote Sensing,2009,13(6):1170-1186.(in Chinese) [23] Tucker C J,Newcomb W W,Los S O,Prince S D.Mean and inter-year variation of growing-season normalized difference vegetation index for the Sahel 1981-1989.International Journal of Remote Sensing,1991,12(6):1133-1135. [24] Milich L,Weiss E.GAC NDVI interannual coefficient of variation (CoV) images:Ground truth sampling of the Sahel along north-south transects.International Journal of Remote Sensing,2000,21(2):235-260. [25] Lunetta R S,Knight J F,Ediriwickrema J,Lyon J G,Worthy L D.Land-cover change detection using multi-temporal MODIS NDVI data.Remote Sensing of Environment,2006,105(2):142-154. [26] Ostendorf B,Hilbert D W,Hopkins M S.The effect of climate change on tropical rainforest vegetation pattern.Ecological Modelling,2001,145(2-3):211-224.2 结果
2.1 基于ANN模型的生物量反演
2.2 三江源区草地地上生物量月际变化


2.3 三江源区草地地上生物量的年际变化

2.4 三江源区草地地上生物量变化趋势分析




3 讨论
3.1 草地地上生物量遥感估算模型精度的影响因素
3.2 人工神经网络在草地AGB模拟中的优缺点
4 结论
