Cry1Ab13杀虫基因的PCR诱变及功能验证
2018-05-24张宇婷代力强王丕武
闫 鸽,宋 阳,张宇婷,张 卓,代力强,王丕武
(1吉林农业大学 农学院,吉林 长春130118;2延边大学 农学院,吉林 延吉 133002)
虫害是影响我国农作物产量的重要因素之一。据国家统计局统计,因虫害的影响,2016年我国夏粮总产量较上年减产162.1万t,经济损失严重[1],其中玉米螟(Ostrinianubilalis)和大豆食心虫(LeguminivoraglycinivorellaMatsumura)等鳞翅目昆虫是主要为害害虫。为有效防治害虫,大量的化学农药被使用,据统计我国农药的单位面积使用量是世界平均水平的2.5倍[2],而大量广泛喷洒化学农药严重影响着人畜和生态环境的安全,同时也会导致害虫产生抗药性,而且会杀死害虫天敌及有益昆虫。因此利用基因工程技术培育具有抗虫性状的品种,是解决虫害问题的有效途径之一。
苏云金芽孢杆菌(Bacillusthuringiensis,Bt)是由日本学者石渡1901年从染病家蚕中首次分离得到,1915年被正式定名为Bacillusthuringiensis(Bt)[3],是一种革兰氏阳性土壤芽孢杆菌。苏云金芽孢杆菌在形成芽孢的同时会产生杀死昆虫幼虫的毒性晶体蛋白[4],这类杀虫晶体蛋白具有高度专一性,对害虫天敌和有益昆虫无毒害作用,只对鳞翅目、鞘翅目等靶标害虫具有杀虫效果[5-6],同时又不会对环境造成污染,因而被广泛应用于害虫防治。截至目前,已成功分离出Cry类Bt基因300多个[7],这些基因根据其编码氨基酸序列同源性,以45%,75%和95%为界限,被分成72类(Cry1-Cry72),其中Cry1A种类最多,共有113种。随着这些Cry类Bt基因及其制剂的大量应用,害虫对其产生了不同程度的抗性[8-9],野生型Bt基因mRNA不稳定也造成该基因表达量极低[10]。因此人们开始运用酶工程技术对Bt类基因进行改造。郭三堆等[11]通过分子设计技术,人工合成Cry1A基因并成功转入棉花中,使我国成为第2个成功培育抗虫棉的国家。王阳等[12]利用结构域交换法对杀虫基因Cry1Ab进行优化改造,获得新型杀虫基因CryFLIa,并且通过室内生测证实该基因对亚洲玉米螟具有明显抗性。现在有关强毒性和新型杀虫活性Cry类、Cyt类、Vip类Bt基因的发掘与克隆,已成为国内外Bt基因研究和利用的热点[13]。
Bt蛋白主要的改造方法有结构域转换法、密码子优化法、DNA Shuffling、易错PCR法等,其中易错PCR技术可以简便有效地向已知DNA序列中引入突变[14],其体系最早由Leung等[15]建立,Cadwell等[16]以该体系进行易错PCR,诱变了编码四膜虫核酶的基因,明显提高了酶的活性。Shelly等[17]通过易错PCR法体外进化芽孢杆菌脂肪酶基因,使其在5 ℃时仍具有活性,增强了该酶的适冷性。本研究运用易错PCR技术,向Cry1Ab13基因原始序列中随机引入突变并建立突变文库,构建3个原核表达载体并进行异源蛋白表达,利用表达的目的蛋白饲喂小菜蛾幼虫(Plutellaxyllostella)和亚洲玉米螟幼虫(Ostrinianubilalis),鉴定诱变基因的抗虫效果,以期为获得杀虫活性更高的新Cry类抗虫基因奠定基础。
1 材料与方法
1.1 材 料
1.1.1 菌株和质粒 本试验所用菌株E.coliDH5α、E.coliBL21和原核表达载体pET-28a,均由吉林农业大学植物生物技术中心保存;克隆载体pMD-18T,购自宝生物工程有限公司;模板质粒pUC57-Cry1Ab13,由吉林省农业科学院提供。
1.1.2 酶和试剂Taq酶等PCR相关试剂和DNA Marker购自TaKaRa公司,限制性内切酶购自Thermo公司,质粒提取试剂盒购自威格拉斯生物技术公司,DNA凝胶回收试剂盒购自爱思进生物技术有限公司,Seamless Assembly Cloning kit购自中美泰和生物技术有限公司,IPTG购自康为世纪生物技术有限公司,其余的药品与试剂均为国产分析纯。
1.1.3 供试昆虫 小菜蛾幼虫于2015年8月下旬在吉林农业大学植物生物技术中心试验田捕捉后,参照郭兆将等[18]的饲养方法室内饲养;亚洲玉米螟幼虫购自天津慧裕德玉米螟养殖场,参照袁志华等[19]的饲养条件饲养。
1.2 方 法
1.2.1 易错PCR扩增 使用软件Primer 5.0,根据Cry1Ab13原始碱基序列设计引物s:TGGACAACAATCCTAACATCAAC,As:TTATCAAA GTTCATCCTTCTCGG,其中下划线部分为随机置换点。以含有pUC57-Cry1Ab13的质粒为模板,参照秦秀林等[20]的方法进行易错PCR。
易错PCR反应体系50 μL,包含ddH2O 17.8 μL,Buffer 5 μL,MgCl220 μL,dNTP 1 μL,引物s 2 μL,引物As 2 μL,模板1 μL,TaqDNA聚合酶 1.2 μL。反应程序为:94 ℃预变性 3 min;94 ℃变性 40 s,40 ℃退火 40 s,72 ℃延伸 3 min, 70个循环;最后72 ℃延伸 10 min,4 ℃保存;不进行热启动。经1%琼脂糖凝胶电泳后纯化回收目的片段,以此片段为模板,以同样的条件进行第2轮易错PCR[21-22],产物经1%琼脂糖凝胶电泳后,纯化并测序。
使用软件DNAman 6.0对测序结果进行比对分析,根据外源蛋白质过敏性分析的国家标准(农业部1485号公告-18-2010),使用在线过敏源数据库(www.allergenonline.org)对其致敏性进行在线分析,利用蛋白质分析软件SOPMA对突变基因进行二级结构分析,通过SWISS-MODEL(www.expasy.org)对测序结果同源建模并进行物理性质分析。
1.2.2 突变体文库的构建 易错PCR产物经纯化后与载体pMD-18T连接,将连接产物转化至E.coliDH5α感受态细胞中,涂布在LB固体培养基(含100 mg/mL氨苄霉素)上,通过测序,筛选发生突变的转化子,组成突变体文库。
1.2.3 原核表达载体的构建 提取1.2.2筛选出突变菌株质粒的DNA,使用带有BamH Ⅰ和SalⅠ酶切位点的无缝引物进行PCR扩增,PCR产物经1%琼脂糖凝胶电泳纯化回收后,使用无缝连接试剂盒将其与载体pET-28a进行连接,再转化到E.coliDH5α感受态细胞中,次日挑取全部单菌落过夜培养,并提取其质粒DNA,进行PCR和酶切验证并测序。
1.2.4 突变基因在大肠杆菌中的诱导表达 将构建好的重组质粒转化到E.coliBL21感受态细胞中,参照宋萍等[23]的方法,用IPTG进行诱导表达和超声波破碎,对表达产物进行SDS-PAGE电泳分析。
1.3 杀虫活性的生物鉴定
对1.2.4节重组菌诱导表达产物进行超声波破碎并回收,用于杀虫活性鉴定。分别于处理后24,48,72 h调查试虫死亡率,用软件Excel和DPS 9.5进行统计与分析。
1.3.1 小菜蛾幼虫杀虫活性的鉴定 采用浸叶法。在实验室中选取幼嫩一致的甘蓝叶片,用清水洗净晾干,放入待测样品中浸泡10 min后晾干,平铺在玻璃培养皿中,以含Cry1Ab13载体的破碎菌液为阳性对照,含空载体的破碎菌液为阴性对照。每个培养皿中接2~3龄小菜蛾幼虫11只,每个处理3次重复,放入人工气候箱中进行培养。培养箱内温度为(25±1.5) ℃,相对湿度为50%~60%,光/暗周期14 h/10 h。
1.3.2 亚洲玉米螟幼虫杀虫活性鉴定 采用人工饲料法。人工饲料的配制和亚洲玉米螟饲养参照乔利等[24]的方法,将破碎的菌液加入到人工饲料中混匀。以含Cry1Ab13载体的破碎菌液为阳性对照,含空载体的破碎菌液为阴性对照,每个处理接入30只3龄幼虫,3次重复。
2 结果与分析
2.1 Cry1Ab13基因的易错PCR扩增
在PCR扩增时高浓度的Mg2+可以降低PCR扩增特异性,稳定非互补碱基配对,增加碱基发生突变的概率。本研究分别以2.0,2.5,3.0,3.5,4.0,4.5,5.0,5.5,6.0,6.5 mmol/L的Mg2+进行PCR,结果(图1-A)表明,2.0~6.0 mmol/L的Mg2+均能扩增出条带,考虑到高浓度Mg2+会增加突变率,所以本研究选择6.0 mmol/L Mg2+进行易错PCR(图1-B)。
2.2 Cry1Ab13突变基因的序列分析
通过测序筛选出3个突变的转化子Cry1Ab13-1、Cry1Ab13-2和Cry1Ab13-3,片段大小分别为2 036,2 033和2 026 bp,与原始碱基序列的一致性分别为97.79%,97.93%和98.32%,其编码的氨基酸与原始氨基酸序列的一致性分别为96.97%,97.12%和97.68%。其中Cry1Ab13-1基因编码氨基酸序列第130位的Pro变成Ser、第383位Ser变成Thr;Cry1Ab13-2基因编码氨基酸序列第424位的Lys变成Thr;Cry1Ab13-3基因编码氨基酸序列第166位的Tyr变成Asn。根据外源蛋白质过敏性分析国家标准,使用在线过敏源数据库(www.allergenonline.org)对3个突变基因致敏性进行在线分析,表明这3个突变基因与已知过敏源序列的同源性均低于35%,不存在致敏性。

A.不同Mg2+浓度电泳:M.DL2000 DNA Marker;H.无菌水;1~10.反应体系中Mg2+浓度依次为2.0,2.5,3.0,3.5,4.0,4.5,5.0,5.5,6.0,6.5 mmol/L;B.6 mmol/L Mg2+电泳:M.DL2000 DNA Marker;1~7.Cry1Ab13的易错PCR扩增A.Eectrophoresis at different Mg2+ concentrations:M.DL2000 DNA Marker;1-10.the concentrations of Mg2+ in the reaction system are 2.0, 2.5,3.0,3.5,4.0,4.5,5.0,5.5,6.0,and 6.5 mmol/L;B.6.0 mmol/L Mg2+ electrophoresis:M.DL2000 DNA Marker;1-7.Cry1Ab13 error-prone PCR amplification图1 Cry1Ab13基因的易错PCR扩增Fig.1 Error-prone PCR amplification of Cry1Ab13 gene
利用蛋白质分析软件SOPMA进行二级结构分析,结果(表1)表明,突变体的α-螺旋和无规则卷曲所占比例略微降低,β-折叠和β-转角所占比例略微升高。

表1 Cry1Ab13基因及突变体的二级结构预测Table 1 Prediction of secondary structure of Cry1Ab13 gene and mutants %
通过SWISS-MODEL服务器模拟突变基因编码蛋白三维空间模型如图2所示,其中红色圈表示发生变化的位置。

A.Cry1Ab13基因编码蛋白三维结构模拟(不同角度);B.从左至右分别为Cry1Ab13-1、Cry1Ab13-2、Cry1Ab13-3基因编码蛋白三维结构模拟,红色圈表示发生变化位置A.Three-dimensional structure simulation of Cry1Ab13 gene encoding protein(Different angles);B.Left to right are Cry1Ab13-1,Cry1Ab13-2,and Cry1Ab13-3;The red circle indicates where change occurred图2 Cry1Ab13基因突变体编码蛋白三维结构的模拟Fig.2 Simulation of three dimensional structure of proteins encoded by gene mutations FIG Cry1Ab13
2.3 突变菌株原核表达载体的构建
挑取全部单菌落过夜培养后提取质粒进行PCR验证,结果如图3-A所示,筛选出稳定的阳性克隆进行酶切验证,得到2个目的条带(图3-B)。测序分析表明,所得序列正确,说明目的基因片段成功连接到原核表达载体pET-28a(+)中,分别命名为Cry1Ab13-1、Cry1Ab13-2和Cry1Ab13-3。

A.PCR验证:M.DL2000 DNA Marker;1~7.重组质粒PCR验证;B.酶切验证:M.DL2000 DNA Marker;1~4.Cry1Ab13-1、Cry1Ab13-2、Cry1Ab13-3、Cry1Ab13基因重组质粒酶切验证 A.PCR validation:M.DL2000 DNA Marker;1-7.Recombinant plasmid was verified by PCR;B.Digestion:M.DL2000 DNA Marker;1-4.Cry1Ab13-1,Cry1Ab13-2,Cry1Ab13-3,and Cry1Ab13 gene recombinant plasmids图3 突变菌株原核表达载体的验证Fig.3 Verification of prokaryotic expression vector of mutant strain
2.4 突变菌株在大肠杆菌中的表达
SDS-PAGE电泳结果(图4)显示,经IPTG诱导后,含CryAb13-1和CryAb13的工程菌(非可溶性组分)在79.5 ku左右分别可以表达出1条明显的特异条带,与预期结果相符合,表明Cry1Ab13-1和Cry1Ab13基因编码的杀虫蛋白在E.coliBL21中得到正确表达。
2.5 Cry1Ab13-1蛋白杀虫活性的生物鉴定
2.5.1 小菜蛾杀虫活性鉴定 试验结果(图5)表明,在试验开始的24 h之内,3组处理小菜蛾均继续大量取食叶片,随着处理时间延长试验组和阳性对照组小菜蛾取食叶片量逐渐减少(叶片孔洞减少),小菜蛾幼虫生长严重受阻,虫体萎缩出现死虫,而阴性对照组正常取食。图6显示,24,48和72 h时试验组小菜蛾的死亡率分别为18.18%,33.31%和87.88%,均高于阳性对照组,并且72 h时试验组小菜蛾的死亡率与阳性对照组和阴性对照组的差异均达到了显著水平,说明Cry1AB13-1基因编码蛋白对小菜蛾有杀虫活性。
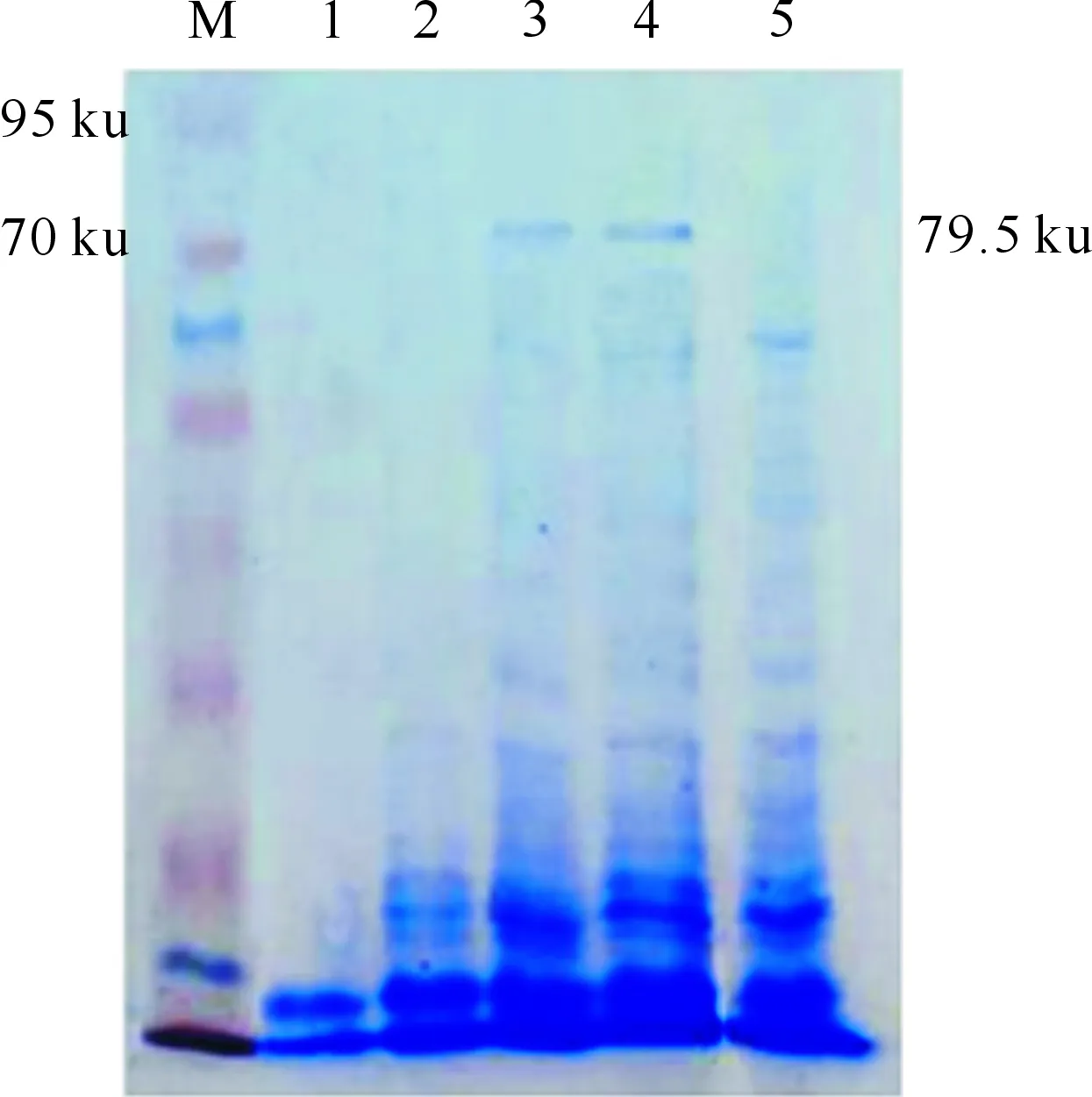
M.蛋白标准分子量;1.Cry1Ab13-3诱导;2.Cry1Ab13-2诱导;3.Cry1Ab13-1诱导;4.Cry1Ab13诱导;5.Cry1Ab13未诱导M.Protein Maker;1.Cry1Ab13-3 induced;2.Cry1Ab13-2 induced;3.Cry1Ab13-1 induced;4.Cry1Ab13 induced;5.Cry1Ab13 un-induced

A.处理后24 h;B.处理后48 h;C.处理后72 h;各图中左、中、右分别为Cry1Ab13、Cry1Ab13-1、空载体处理A.24 h after treatment;B.48 h after treatment;C.72 h after treatment;In the figure,the left,middle and right are Cry1Ab13,Cry1Ab13-1,and empty vector
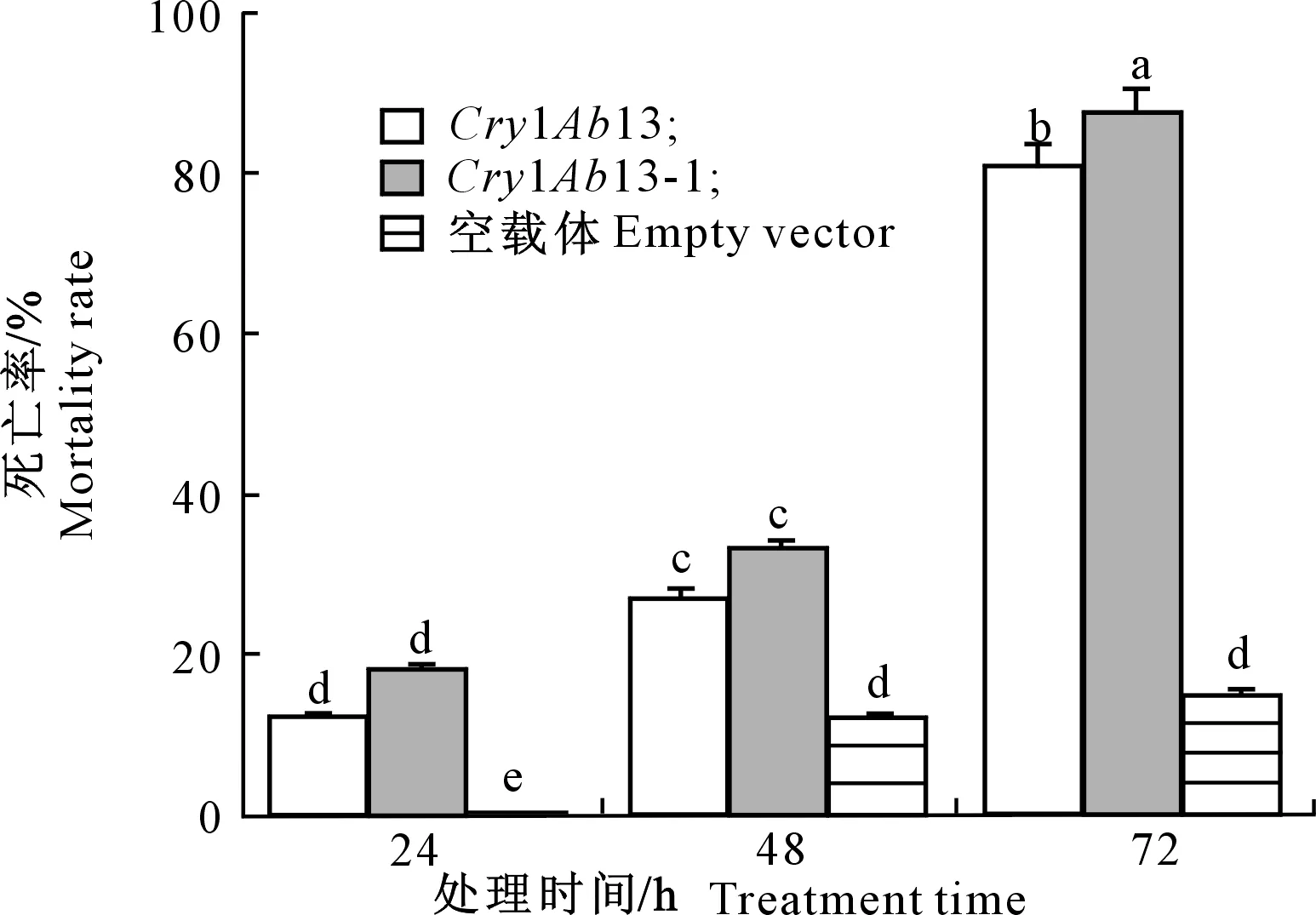
图柱上不同小写字母代表5%差异水平。下同Lowercase letters represent significant difference at 5% level.The same below图6 处理不同时间小菜蛾幼虫死亡率的比较Fig.6 Comparison of larvae mortality in Plutella xylostella at different times
2.5.2 亚洲玉米螟杀虫活性鉴定 试验中观察发现,亚洲玉米螟幼虫室内饲养24 h后,处理组出现变软、发黑和死亡的幼虫,而阴性对照组幼虫生长正常,随着处理时间延长,处理组幼虫逐渐停止活动,死虫数目逐渐增加。由图7-A可知,处理24,48和72 h后,试验组亚洲玉米螟幼虫死亡率分别为12.22%,23.89%和81.11%,与阳性对照组差异不明显,仅在72 h时略低于阳性对照组。图7-B表明,存活的亚洲玉米螟幼虫的生长受到严重抑制,72 h后试验组虫体质量仅为处理前的7%,阴性对照组虫体质量为处理前的98%,试验组和阴性对照组虫体质量差异达显著水平。上述结果表明,新诱变的基因可以编码出具有杀虫活性的毒蛋白,对小菜蛾和亚洲玉米螟具有很强的毒杀作用。
图7 亚洲玉米螟幼虫室内杀虫活性的鉴定Fig.7 Identification of insecticidal activity against Asian corn borer larvae
3 讨 论
苏云金芽孢杆菌(Bacillusthuringiensis,Bt)是农林业上应用最广也是最有效的微生物杀虫剂,但是随着大面积推广使用,在持续定向选择压作用下容易使昆虫产生抗性,并且来自微生物中的Bt基因碱基序列中含有大量AT碱基对,致使这类基因在植物中的表达量较低。因此对Bt基因进行优化改造,可以作为提高Bt基因蛋白表达量、增强杀虫效果的突破点,同时还可以丰富抗虫基因资源。岳润清等[25]对苏云金芽胞杆菌Cry1Ab野生型基因进行了改造,改造后的Cry1Ab基因编码蛋白对玉米螟幼虫的致死率提高到86.3%。任羽等[26]对Cry1Ca7基因作定点突变后发现,结构域Ⅰ中突变产生的毒性较Cry1Ca7基因提高了6倍,位于其他结构域的突变毒性均有所降低。
易错PCR是一种简单、方便的基因体外诱变技术,其突变频率明显高于自然突变,但该技术在使用过程中也存在弊端,例如突变频率过高、有害突变增加、很难筛选到带有目标性状的突变株等。有研究表明,合理控制突变频率是易错PCR成功的关键,一般每个基因有1~4个碱基发生突变,对应导致1~2个氨基酸发生改变最为适宜[27]。黄瑛等[28]通过调整易错PCR反应体系中的Mn2+浓度对BpL基因进行体外诱变,使YZ02脂肪酶活性提高了6倍。Drummond等[27]等在反应体系中加入丙醇进行易错PCR后发现,丙醇可以通过影响TaqDNA聚合酶活性提高突变频率。秦秀林等[20]通过改变反应体系中的模板浓度和反应循环数,经过3轮易错PCR诱变GAP启动子,获得了5个强度提高的启动子。目前该技术在改造Cry类Bt基因方面利用较少,本研究将反应体系中的Mg2+浓度提高到6.0 mmol/L,对Cry1Ab13基因进行2轮易错PCR诱变,获得了带有2个碱基突变位点的突变株Cry1Ab13-1。将Cry1Ab13-1基因构建重组表达载体后进行异源蛋白表达,利用表达的目的蛋白饲喂小菜蛾幼虫和亚洲玉米螟幼虫, 鉴定诱变基因的抗虫效果,结果表明,该表达蛋白对小菜蛾幼虫和亚洲玉米螟幼虫具有强毒杀作用,并可以抑制亚洲玉米螟幼虫生长,与余宗兰等[29]利用常规技术克隆的Cry类Bt基因具有相似的杀虫效果,表明易错PCR技术在改造Cry类Bt基因方面具有可行性。由于植物中的密码子偏好性与细菌中的密码子偏好性不同,所以本研究下一步计划将该基因转入玉米、大豆、水稻等作物中,进而在真核生物中进一步验证该基因的杀虫效果。
[参考文献]
[1] 国家统计局.国家数据 [DB].(2016-11-20).http://data.stats.gov.cn/easyquery.htm?cn=C01.
National Bureau of Statistics of China.National date [DB].(2016-11-20).http://data.stats.gov.cn/easyquery.htm?cn=C01.
[2] 陈晓明,王程龙,博 瑞.中国农药使用现状及对策建议 [J].农药科学与管理,2016,37(2):4-8.
Chen X M,Wang C L,Bo R.Current situation of Chinese pesticide application and policy suggestions [J].Pesticide Science and Administration,2016,37(2):4-8.
[3] Breed.伯杰氏细菌鉴定手册 [M].北京:科学出版社,1984.
Breed.Bergey’s manual of determinative bacteriology [M].Beijing:Science Press,1984.
[4] 郭三堆.植物Bt基因工程研究进展 [J].中国农业科学,1995,28(5):8-13.
Guo S D.Engineering of insect-resistant plant withBbacillusthuringiensiscrystal protein genes [J].Scientia Agricultura Sinica,1995,28(5):8-13.
[5] 叶恭银,陈 洋,田俊策,等.转Bt基因抗虫作物对非靶标生物的影响 [J].植物保护,2011,37(6):1-10.
Ye G Y,Chen Y,Tian J C,et al.The effects of transgenic Bt insect-resistant crops on non-target organisma [J].Plant Protection,2011,37(6):1-10.
[6] 杨 艳,李云河,曹凤勤,等.转Bt基因抗虫作物对鳞翅目非靶标昆虫生态影响的研究进展 [J].生物安全学报,2014,23(4):224-237.
Yang Y,Li Y H,Cao F Q,et al.Progress in the assessment of ecological effects of insect-resistant Bt crops on non-target of lepidopteran insects [J].Journal of Biosafety,2014,23(4):224-237.
[7] Ernest S.A relational database giving known insecticidal activities of the delta-endotoxins [J/OL].(2016-11-20).http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/.
[8] 林珠凤,陈海燕,吉训聪,等.害虫对转Bt基因作物的抗性及治理 [J].热带生物学报,2015,6(4):497-503.
Lin Z F,Chen H Y,Ji X C,et al.Insect resistance to Bt crops: possible mechanism and resistance management strategy [J].Journal of Tropical Biology,2015,6(4):497-503.
[9] 李怡萍,梁革梅,陈 豪,等.苏云金芽孢杆菌杀虫机理及害虫对其抗性机制的研究进展 [J].西北农林科技大学学报(自然科学版),2010,38(9):118-128.
Li Y P,Liang G M,Chen H,et al.Progress in insecticidal mechanism of Bt and resistance mechanism of pest insect to Bt [J].Journal of Northwest A &F University(Nat Sci Ed),2010,38(9):118-128.
[10] Barton K A,Whiteley H R,Yang N S.BacillusthuringiensisDelta-endotoxin expressed in transgenicNicotianatabacumprovides resistance to lepidoptern insects [J].Plant Physiol,1987,85:1103-1109.
[11] 郭三堆,王 远,孙国清,等.中国转基因棉花研发应用二十年 [J].中国农业科学,2015,48(17):3372-3387.
Guo S D,Wang Y,Suo G Q,et al.Twenty years of research and application of transgenic cotton in China [J].Scientia Agricultura Sinica,2015,48(17):3372-3387.
[12] 王 阳,杨 朔,尹悦佳,等.分子改造Cry蛋白基因的转基因玉米创制及其抗虫功能评价 [J].玉米科学,2015,23(6):27-34.
Wang Y,Yang S,Yin Y J,et al.Rational mutation of cry protein and the insect resistance assessment in the transgenic maize [J].Journal of Maize Sciences,2015,23(6):27-34.
[13] 关 雄,蔡 俊.我国苏云金杆菌研究60年 [J].微生物学通报,2014,41(3):459-465.
Guan X,Cai J.Bacillusthuringiensis:sixty years of research development and commercial applications in China [J].Microbiology China,2014,41(3):459-465.
[14] 高义平,赵 和,吕梦雨,等.易错PCR研究进展及应用 [J].核农学报,2013,27(5):607-612.
Gao Y P,Zhao H,Lü M Y,et al.Research progress and application of error-prone PCR [J].Journal of Nuclear Agricultural Sciences,2013,27(5):607-612.
[15] Leung D W,Chen E,Goeddel D V.A method for random mutagen of a defined DNA segment using a modified polymerase chain recation [J].Technique,1989,1:11-15.
[16] Cadwell C,Joyce G F.Randomization of genes by PCR mutagenesis [J].PCR Methods and Applications,1992,2:28-33.
[17] Shelly G,Arbind K,Jagdeep K.Disruption of N terminus long range non covalent interactions shifted temd.opt 25 ℃ to cold:evolution of point mutantBacilluslipase by error prone PCR [J].Gene,2015,10:29-31.
[18] 郭兆将,康 师,吴青君,等.小菜蛾饲养技术与规范 [J].应用昆虫学报,2015,52(2):492-497.
Guo Z J,Kang S,Wu Q J,et al.Technical specifications for the mass-rearing of the diamondback moth,Plutellaxylostella(L.) [J].Chinese Journal of Applied Entomology,2015,52(2):492-497.
[19] 袁志华,王文强,王振营,等.亚洲玉米螟的寄主植物种类 [J].植物保护学报,2015,42(6):957-964.
Yuan Z H,Wang W Q,Wang Z Y,et al.Host plants of the Asian corn borer,Ostriniafurnacalis(Guenée) (Lepidoptera:Crambidae) [J].Journal of Plant Protection,2015,42(6):957-964.
[20] 秦秀林,钱江潮,储 炬,等.优化易错PCR条件以提高毕赤酵母GAP启动子文库突变效率 [J].生物技术通报,2016,10(6):211-217.
Qin X L,Qian J C,Chu J,et al.High-error-rate random mutagenesis of GAP promoter in pichia pastoris using an optimited error prone PCR [J].Biotechnology Bulletin,2016,10(6):211-217.
[21] 曾家豫,郑 群,廖世奇,等.大豆过氧化物酶基因随机突变文库的构建 [J].大豆科学,2011,30(5):731-737.
Zeng J Y,Zheng Q,Liao S Q,et al.Construction of soybean peroxidase gene random mutation library [J].Soybean Science,2011,30(5):731-737.
[22] 刘卫晓,钱世君.酶分子体外定向进化的研究方法 [J].微生物学报,2004,31(2):100-104.
Liu W X,Qian S J.Methods of the directed molecular evolution of enzymeinvitro[J].Microbiology China,2004,31(2):100-104.
[23] 宋 萍,南宫自艳,张 杰,等.苏云金芽胞杆菌Cry1Ia32基因克隆和原核表达的研究 [J].河北农业大学学报,2015,38(5):48-52.
Song P,Nangong Z Y,Zhang J,et al.Cloning and expression ofCry1Ia32 gene fromeBaciliusthuringiensisisolate [J].Journal of Agricultural University of Hebei,2015,38(5):48-52.
[24] 乔 利,郑坚武,成卫宁,等.不同饲料配方对亚洲玉米螟生长发育和繁殖的影响 [J].西北农林科技大学学报(自然科学版),2008,36(5):109-112.
Qiao L,Zheng J W,Cheng W N,et al.Impact of 4 different artificial fodders on life span of asian corn borer,Ostriniafurnacalis(Guenee) [J].Journal of Northwest A &F University (Nat Sci Ed),2008,36(5):109-112.
[25] 岳润清,铁双贵,陈小洁,等.人工改造基因Cry1Abm的合成、原核表达及其抗虫鉴定 [J].玉米科学,2013,21(6):21-25.
Yue R Q,Tie S G,Chen X J,et al.Synthesis of the modifiedCry1Abmgene,expressioninE.coliand bioassay [J].Journal of Maize Sciences,2013,21(6):21-25.
[26] 任 羽,刘华梅,宋福平,等.苏云金芽孢杆菌Cry1Ca7蛋白定点突变对甜菜夜蛾杀虫活性的影响 [J].微生物学报,2008,48(6):733-738.
Ren Y,Liu H M,Song F P,et al.Effect ofCry1Ca7 protein modified by site-directed mutagenesis on inhibitingSpodopteraexiguaHübne [J].Acta Microbiologica Sinica,2008,48(6):733-738.
[27] Drummond D A,Iverson B L,Georgiou G,et al.Why high-error-rate random mutagenesis libraries are enriched in functional and improved proteins[J].Journal of Molecular Biology,2005,350(4):806-816.
[28] 黄 瑛,蔡 勇,杨江科,等.基于易错PCR技术的短小芽孢杆菌YZ02脂肪酶基因BpL的定向进化 [J].生物工程学报,2008,24(3):445-451.
Huang Y,Cai Y,Yang J K,et al.Directed evolution of lipase ofBacilluspumilusYZ02 by error-prone PCR [J].Chinese Journal of Biotechnology,2008,24(3):445-451.
[29] 余宗兰,贺利业.苏云金芽胞杆菌cry1Ie新基因的克隆、表达与生物活性测定 [J].植物保护学报,2016,43(2):201-206.
Yu Z L,He L Y.Cloning,expression,and biological activity of a newcry1Iegene fromBacillusthuringiensisstrain [J].Journal of Plant Protection,2016,43(2):201-206.
