浙江沿岸海域春季螺类群落结构特征及其与环境因子的关系
2018-05-16卢占晖朱文斌徐开达周永东卢衎尔
卢占晖 朱文斌 徐开达 周永东 戴 乾 卢衎尔
(浙江省海洋水产研究所, 浙江省海洋渔业资源可持续利用技术研究重点实验室, 农业部重点渔场渔业资源科学观测实验站, 舟山 316004)
螺类是软体动物腹足纲的通称, 是软体动物门最大的一个纲, 占软体动物种类总数的80%左右,广泛栖息于海洋、淡水以及陆地等各种环境中[1]。在海洋生态系统中, 它们既可作为分解者分解动植物残体和有机碎屑, 又能作为消费者牧食大型藻类和维管植物, 还能作为次级生产者被鱼类和甲壳类等多种海洋动物所摄食[2,3]。因此, 螺类在海洋生态系统中属于消费者亚系统, 是该生态系统中物质循环与能量流动中积极的消费者与转移者, 同时也是海洋水体的环境指示生物, 在海洋生态系统中具有多种生态功能, 是食物网的重要环节[4]。目前, 有关螺类种类组成、数量分布以及群落特征方面的研究主要集中在河流、湖泊等淡水水域[5,6]以及海洋潮间带区域[7—9], 而海洋螺类方面的报道主要集中在种类组成的初步研究[10,11]以及系统分类学研究[12,13]等方向, 涉及到近海螺类群落方面的报到则主要是作为组成类群之一在大型底栖生物或软体动物的研究中得以体现[14—16]。
浙江近海海域水系分布复杂, 营养物质丰富,包括螺类在内的海洋大型底栖动物种类繁多[17], 分布广泛, 是该海域海洋生态系统的重要组成部分。然而, 随着近年来海洋捕捞作业, 尤其是大型底拖网的捕捞强度逐渐增强, 加之涉海工程和海洋污染的影响, 近海底栖生境受到了较为严重的破坏, 势必对近海螺类群落的结构及其分布造成一定程度的影响。本文通过对浙江近海海域螺类资源的调查, 对其种类组成、优势种、资源密度分布、群落多样性、外部因素对其群落结构的干扰程度及其与环境因子的关系进行了初步分析, 旨在探明该海域螺类资源的物种多样性, 丰富我国近海软体动物地理区系的基础资料, 同时也为浙江近海海洋生态和环境保护措施的制定提供参考依据。
1 材料与方法
1.1 数据来源
本文所用数据来自2015年春季(4月)进行的“浙江沿岸产卵场调查与选划”课题。共设置调查站位87个, 站位分布如图 1所示。调查船为单船底拖网渔船, 船长38 m, 总吨位150 t, 主机功率202 kW。调查网具网口拉紧周长50 m, 囊网网目尺寸25 mm,上纲长30 m, 下纲长38 m。各站位拖网时间均为1h, 平均拖速3 kn。各站位渔获物样品随机采样1箱(20 kg), 渔获物中螺类种类尽可能鉴定到最小分类单元, 并记录每种的重量和尾数, 螺类种类名称及分类地位以《中国海洋生物名录》[18]为依据, 上述调查采样及测定均按《海洋调查规范(GB/T 12763.6—2007)》[19]的有关标准进行。根据渔场环境条件及其生物种类分布特征, 将调查海域分为3个区域: 即将29°30′N以北(1—38号站位)水域称为北部海域, 28°30′N以南(63—87号站位)水域称为南部海域, 28°30′—29°30′N (39—62号站位)之间的水域称为中部海域。
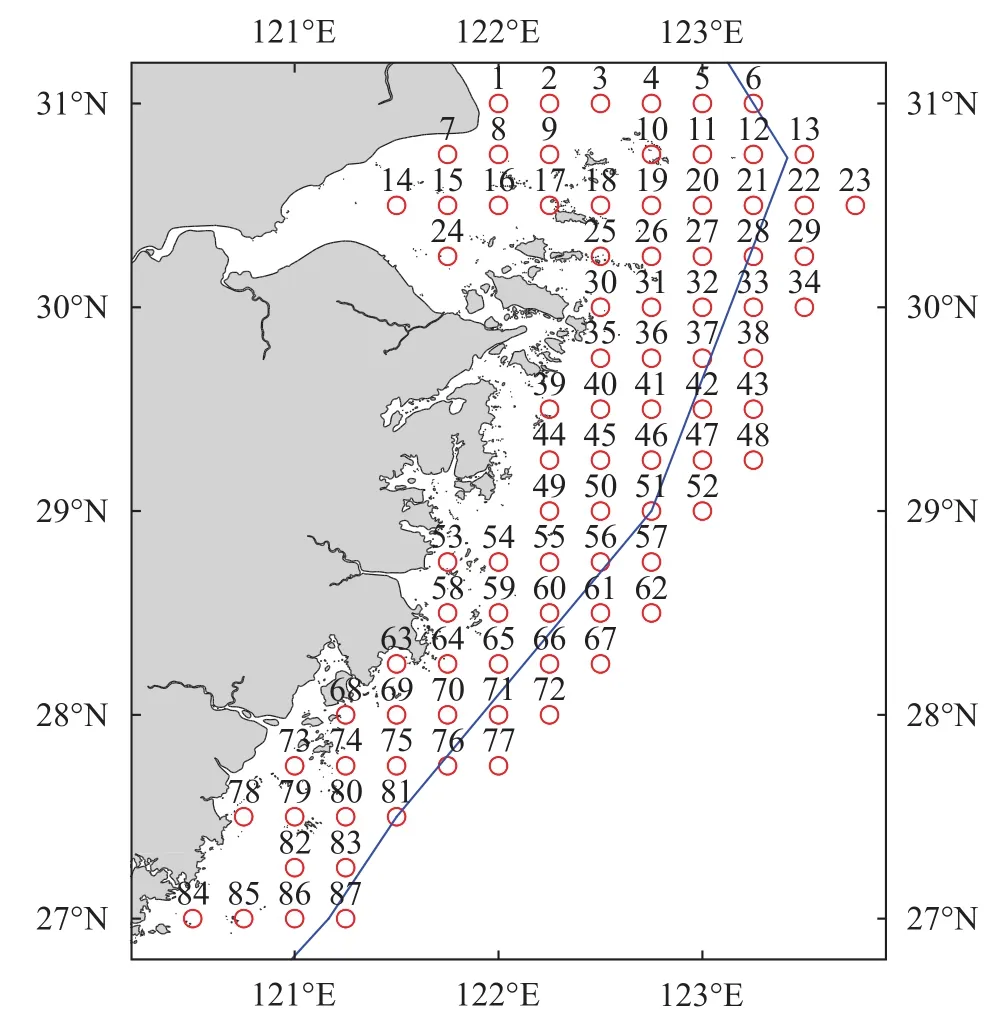
图 1 浙江沿岸海域调查站位分布图Fig. 1 Location of sampling stations along Zhejiang coast曲线代表拖网禁渔区线, 下同Blue curve indicates the boundary for the closure of fishing areas,the same applies below
使用多功能水质监测仪(JFE-AAQ171)对各站位海洋环境参数进行同步采集。采集环境参数包括: 表层水温(ST)、底层水温(BT)、表层盐度(SS)、底层盐度(BS)、表层溶解氧(SD)、底层溶解氧(BD)、表层叶绿素a(SC)、底层叶绿素a(BC)和水深(D)。
1.2 分析方法
优势种与常见种群落优势种与常见种采用Pinkas的相对重要性指数(Index of relative importance,IRI)进行划分[20]。其公式为:
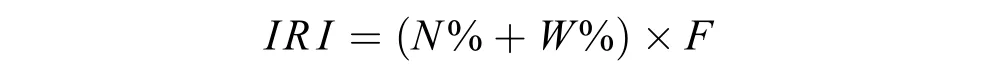
式中,N%为某一种类的尾数占总尾数的百分比;W%为某一种类的重量占总重量的分比;F为某一种类出现的站数占调查总站数的百分比, 根据不同螺类优势度(IRI)的大小, 确定不同种类在群落内的重要性。本研究将相对重要性指数(IRI)大于1000的种类定为优势种, 100—1000之间的种类定为常见种。
资源密度本文采用扫海面积法对调查海域螺类资源密度分布进行分析[21]。其计算公式为:
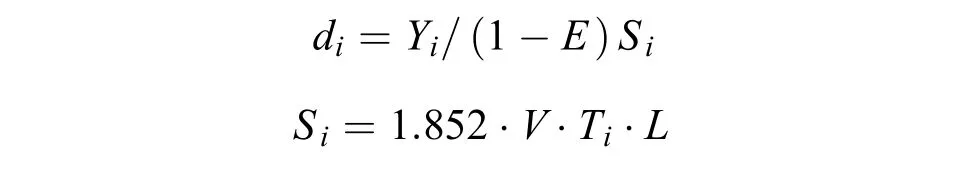
式中,di为i站位的资源密度(kg/km2或103ind./km2),Yi为调查船在i站位的渔获量(kg)或尾数(ind.);Si为调查船在i站位的扫海面积(km2);E为逃逸率(根据本次调查网具——单船底拖网的结构, 渔法特点以及螺类分布水层和生活习性(基本为底栖生活,游泳能力较弱), 综合相关研究报告逃逸率E的取值依据[22,23], 本研究选取的逃逸率为0.2);V为网具拖曳的平均拖速(kn)(本次调查拖网拖曳平均拖速为3 kn);Ti为拖网时间(本次调查均为1h);L为网口水平扩张宽度(km)(本次调查网具为5×10–3km)。
群落多样性本文螺类群落多样性分析运用下列公式[24],
种类丰富度指数D:
式中,S为某站位的螺类种类数;N为该站位总个体数;Pi为该站位第i种螺类个体数占螺类总渔获个体数的比例。
群落ABC曲线本文根据ABC曲线中生物量和丰度的K-优势度曲线, 分析检验浙江沿海海域螺类群落受到污染和扰动的情况。若生物量曲线位于丰度曲线之上, 则表明群落处于未受干扰(稳定)的状态; 若2条曲线相交, 则表明群落处于中等干扰(或不稳定)的状态; 若生物量曲线在丰度曲线之下, 则表明群落处于严重干扰的(不稳定的)状态。用W统计量(W-statistic)作为ABC曲线方法的一个统计量, 其公式为:
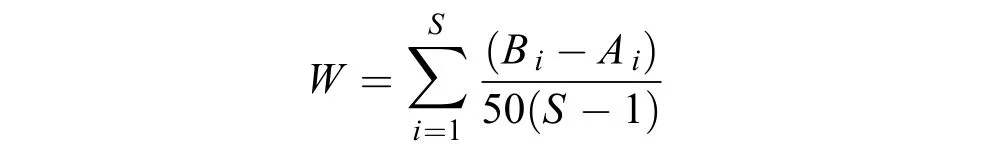
式中Bi和Ai为ABC曲线中种类序号对应的生物量和丰度的累积百分比,S为出现物种数。当生物量优势曲线在丰度优势度曲线之上时,W为正, 反之W为负[25]。
群落与环境因子的关系采用典范对应分析(Canonical Correspondence Analysis, CCA)对浙江沿岸海域春季的调查数据与环境因子的关系进行分析, 分析前先对螺类种类进行筛选, 排除出现频率少于1个站位的物种, 以减少机会种对分析的干扰。在数据分析之前, 物种数据和环境数据均进行log(x+1)转换, 以使它们的分布更接近正态分布[26]。
以上数据分析中, 群落的物种多样性指数、ABC曲线及W统计量的计算均使用PRIMER6.0软件进行处理; 典范对应分析(CCA)在CANOCO4.0软件进行运算, 排序结果用物种-环境因子关系的双序图表示, 站位图、种类分布图以及资源密度分布图利用surfer作图软件进行绘制。
2 结果
2.1 种类组成与优势种
春季拖网调查中共鉴定螺类种类数27种(表 1),隶属于2目, 15科, 21属。其中, 玉螺科出现种类数最多, 有4种; 其次为鹑螺科、塔螺科和织纹螺科,出现种类数均为3种; 骨螺科、盔螺科和衲螺科出现种类数均为2种; 蛾螺科、凤螺科、冠螺科、蛙螺科、涡螺科、细带螺科、衣笠螺科和锥螺科出现种类数均为1种。不同站位间出现的螺类种类数最多的为13种, 出现在南部海域的78号站位, 最少的仅有1种, 出现在北部海域的4、11、12和32号站位, 螺类种类数的空间分布特征呈现由北向南逐渐增多的趋势(图 2)。

表 1 浙江沿岸海域春季螺类群落种类组成及其优势度Tab. 1 The species composition and dominance of snail community along Zhejiang coast in spring
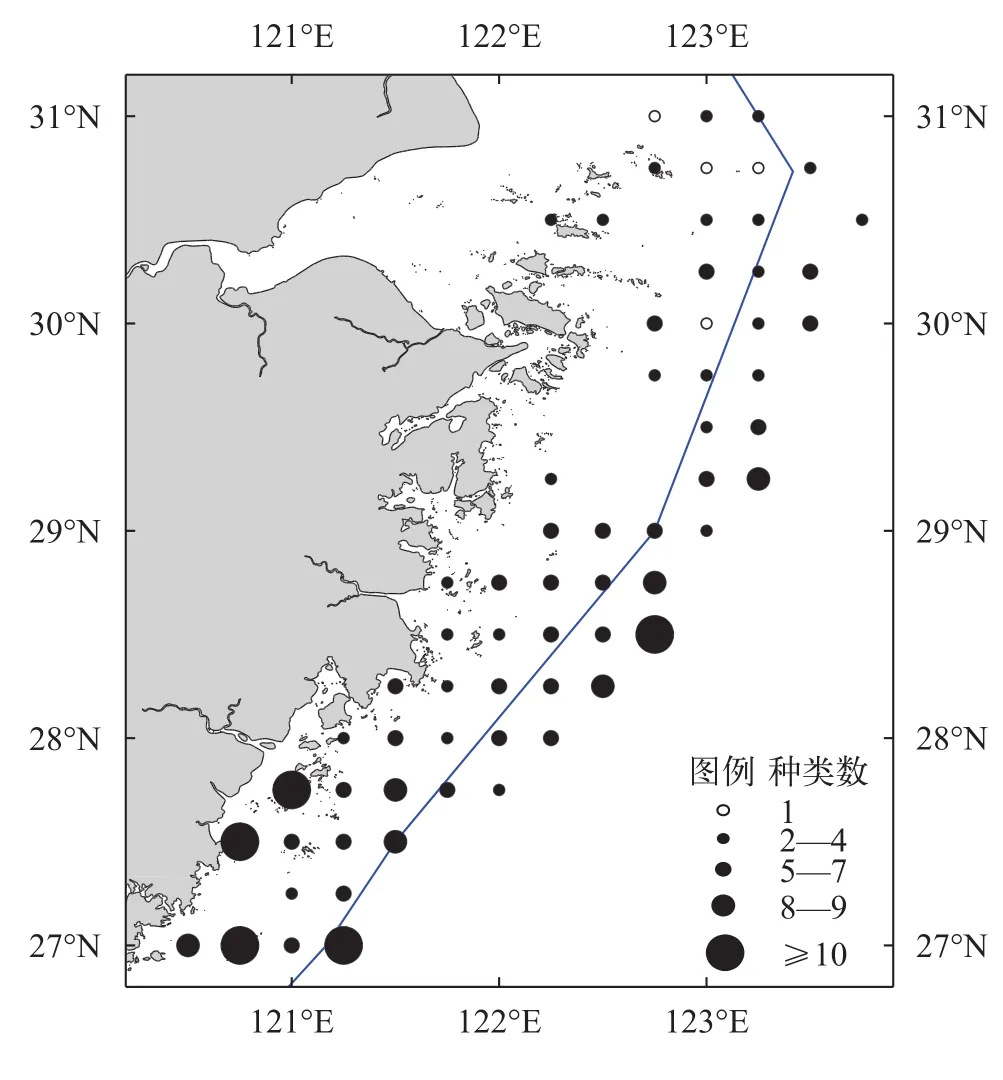
图 2 春季浙江沿岸海域螺类种类分布Fig. 2 The distribution of snail species along Zhejiang coast in spring
按照上述规定, 螺类群落的优势种为棒锥螺(Turritella bacillum)和褐管蛾螺(Siphonalia spadicea), 这2种螺类渔获质量占螺类总渔获物质量的80.76%, 渔获个体数占螺类总渔获物个体数的89.44%;常见种为习见蛙螺(Bursa rana)、长尾纺锤螺(Fusinus longicaudus)和扁玉螺(Nevetita didyma)3种(表 1)。
2.2 资源密度
春季浙江沿岸海域螺类出现站位为66个, 占调查总站位数的75.86%, 捕获螺类共计40696个,361.77 kg。螺类平均资源密度(重量)为246.64 kg/km2,资源密度(重量)最小值为0.45 kg/km2, 出现在位于舟山渔场的20号站位, 资源密度(重量)最大值为3067.49 kg/km2, 出现在位于温台渔场的85号站位;螺类平均资源密度(尾数)为27.75×103ind./km2, 资源密度(尾数)最小值为0.07×103ind./km2, 出现在位于长江口渔场的4号站位, 资源密度(尾数)最大值为350.97×103ind./km2, 出现在位于温台渔场的85号站位。
从整个调查海域螺类资源密度空间分布看, 浙江沿岸海域螺类重量资源密度和尾数资源密度的分布趋势基本一致: 出现2个高密度分布区, 一个为28°30′N至29°30′N的浙江中部海域, 一个为27°30′N以南的浙江南部海域。总体分布趋势呈现较为明显的斑块分布特征, 但北部调查海域资源密度低于中南部海域(图 3)。
2.3 群落多样性
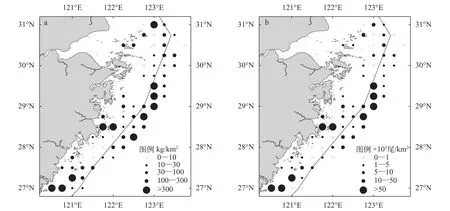
图 3 浙江沿岸海域春季螺类资源密度分布Fig. 3 The spatial distribution of snail density along Zhejiang coast in spring
浙江沿岸春季各站位螺类种类丰富度指数(D)的平均值为0.36, 其范围为0—1.30; 物种多样性指数(H′) 的平均值为0.56, 其范围为0—1.89; 种类均匀度指数(J′) 的平均值为0.50, 其范围为0.02—0.98。以上3种群落多样性指数均较小, 螺类群落多样性水平处于较低水平。从3个生物多样性指数空间分布来看: 物种丰富度(D)在调查海域整体较为均匀, 仅在30°N以北, 123°E以东以及南部海域的零星站位的指数相对较高; 物种多样性指数(H′) 在3个指数中空间分布特征相对较为显著, 其分布呈现拖网禁渔区线以外高, 以内低, 即近岸低,外海高的趋势; 均匀度指数(J)的趋势性分布性特征较为不明显, 仅在北部的外侧海域和中南部的零星站位存在较小的高值区。单因素方差分析表明: 以拖网禁渔区线为远近岸分界线,D、H′、J三种多样性指数远近岸差异均不存在显著差异(P>0.05)(图 4)。
2.4 群落ABC曲线
浙江沿岸海域春季螺类群落生物量和丰度的K-优势度曲线相交, 而且W统计值(–0.024)为负值,说明春季调查海域螺类群落处于中等干扰(或不稳定)状态(图 5)。
2.5 群落与环境因子关系的CCA分析
CCA排序结果显示: 第一排序轴和第二排序轴的特征值分别为0.477和0.383, 共解释了种类变异的24.3%和70.5%, 前2个环境因子排序轴与种类排序轴之间的相关系数分别为86.4%和86.5%, 前2个种类排序轴近似垂直(相关系数–0.0549), 前2个环境排序轴的相关系数为0。以上均证实排序轴与环境因子间线性结合的程度较好地反映了群落与环境之间的关系, 排序的结果是可靠的[33,34](图 6)。
综合出现种类与环境因子的CCA排序图(图 5)和环境因子与排序轴的相关性大小(表 2)可得, 第一排序轴与底层溶解氧呈显著正相关(P<0.05), 相关系数为0.5020, 与水深呈极显著负相关(P<0.001),相关系数为–0.8320, 即沿CCA排序轴第一轴从左到右底层溶解氧逐渐升高, 水深逐渐降低, 所有9个环境因子中, 水深与第一轴的相关性最大; 第二排序轴与底层溶解氧呈显著正相关(P<0.05), 相关系数为0.6020, 与底层温度呈极显著负相关(P<0.001),相关系数为–0.7701, 即沿CCA排序轴第二轴从下到上, 底层溶解氧逐渐升高, 底层温度逐渐降低, 所有9个环境因子中, 底层温度与第二轴的相关性最大。综合前两轴相关性结果: 水深、底层温度和底层溶解氧等3个环境因子对螺类群落的分布起较大的作用。
3 讨论
3.1 种类组成和优势种
根据本次调查结果: 浙江沿岸海域春季螺类种类共出现27种。其中, 玉螺科种类最多, 有4种, 其次为塔螺科、织纹螺科和鹑螺科, 均为3种, 以上4科种类占总种类数的48.15%。不同海域种类分布的特征呈现出由北向南种类数逐渐增多的趋势, 这主要是由于随着水温由北向南逐渐升高以及外海黑潮暖流影响的逐渐增强, 除某些广布种以外, 中南部海域出现了更为典型的热带性种类(如沟鹑螺、管角螺等), 这一特征与以前的研究结果也较为一致[17]。从种类的区系特征来看, 浙江沿岸地处东海大陆架海域, 整个软体动物区系属于印度—西太平洋区系, 因此调查海域螺类种类绝大多数属于暖水性种类, 很少有暖温性种类渗入(本次调查中螺类种类均为暖水性, 未出现暖温性种类)。因此,浙江沿海海域螺类种类组成表现中典型的亚热带性质[17]。与其他海域种类组成相比, 南海的北部湾和大亚湾海域腹足类种类数分别为56种[27]和32种[28],黄海的胶州湾和莱州湾海域螺类种类分别为24种[29]和20种[30], 这也可以从一定程度上说明海洋螺类种类数随着纬度降低而逐渐增加。
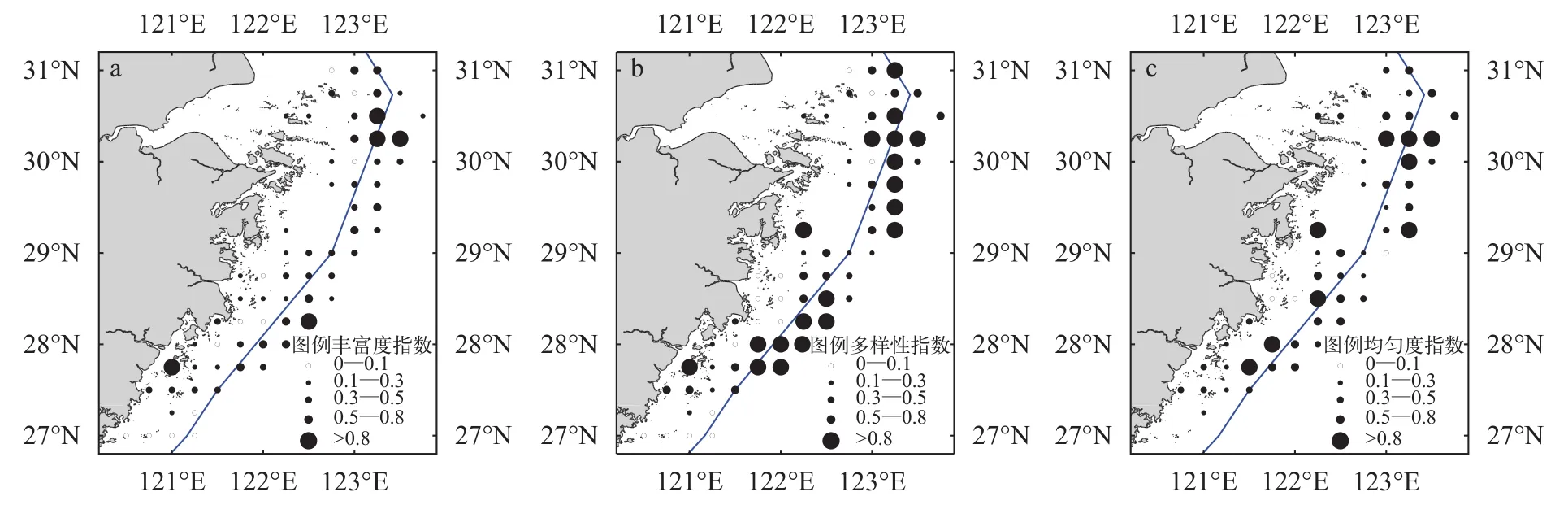
图 4 浙江沿岸海域春季螺类群落生物多样性指数分布Fig. 4 The distribution of biodiversity indices for snails community along Zhejiang coast in spring
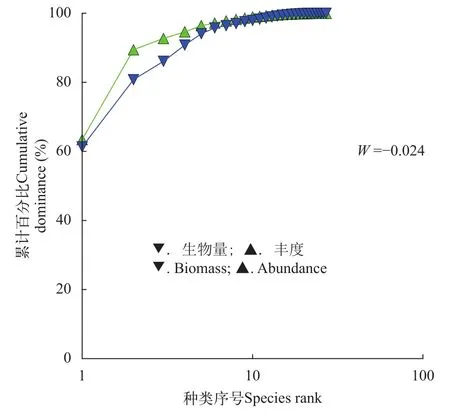
图 5 浙江沿岸海域春季螺类群落的ABC曲线及W统计值Fig. 5 ABC curve and W value of snail community along Zhejiang coast in spring
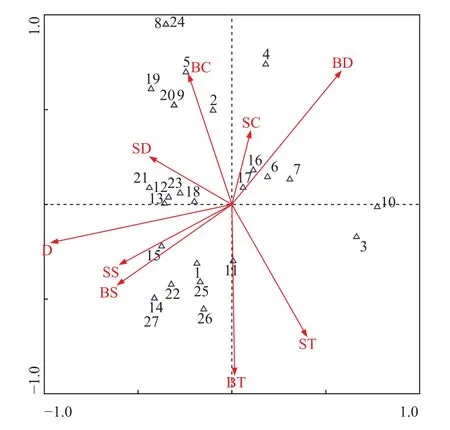
图 6 浙江沿岸海域春季螺类群落种类与环境因子的CCA排序图Fig. 6 Biplot of CCA ordination between different snail species and environmental factors along Zhejiang coast in spring
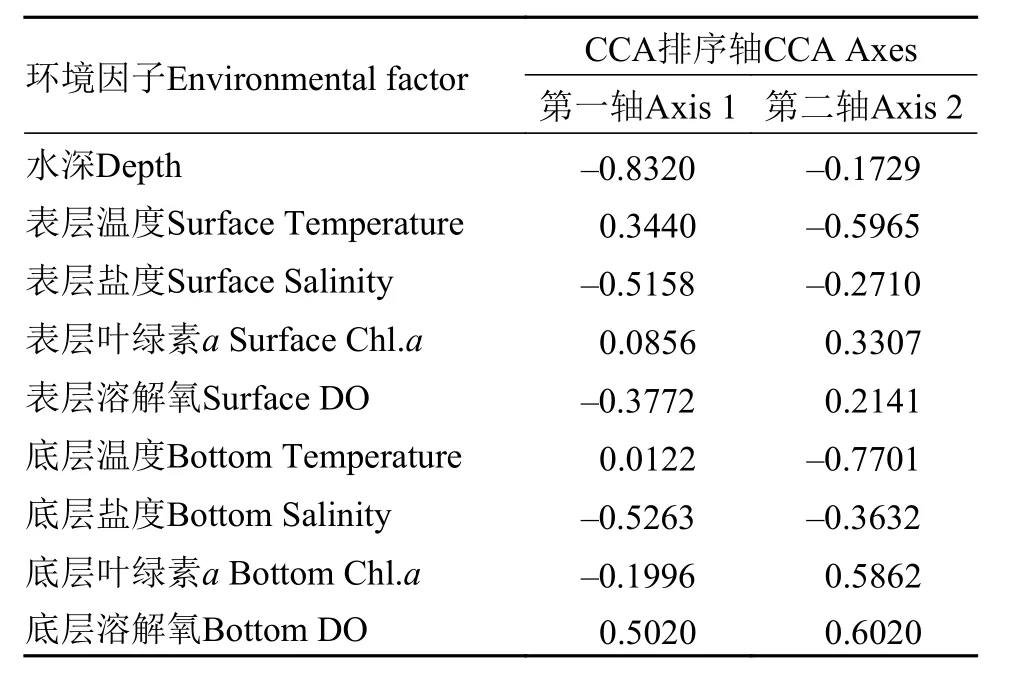
表 2 浙江沿岸海域春季螺类群落环境因子与CCA环境轴的相关性Tab. 2 Correlation between environmental factors for snail communities and CCA environmental axes along Zhejiang coast in spring
在本次调查中, 螺类群落的优势种为棒锥螺和褐管蛾螺两种。其中, 棒锥螺为春季螺类群落的主要优势种, 且主要分布在29°N以南的调查海域, 这也是由该种类的生态习性所决定的[17]。洞头列岛和椒江口海域的研究结果同样印证了此结论[31—32]。此外, 根据相对重要性指数(IRI)数值, 棒锥螺和褐管蛾螺的优势度明显高于其他种类, 且群落中偶见种较多(如: 白龙骨乐飞螺、斑鹑螺和假奈拟塔螺等)。综合这2种因素可见: 浙江沿岸海域螺类群落结构较为简单, 群落的性质和功能只是通过少数种类进行控制。究其原因主要是由于包括螺类在内的底栖动物与其生活的底质沉积生境有密切关系,人类活动常会导致底栖动物群落结构产生显著的变化[28], 而本次调查的浙江沿岸海域恰好为人类活动(主要是捕捞活动与涉海工程)较为频繁的海域,从而造成了调查海域螺类群落结构的简单化。这一点在浙江典型岛礁附近海域大型底栖动物的群落特征中同样得到印证[33]。
以往对于螺类乃至大型底栖动物丰度及生物量分布的研究, 在潮间带海域较多。对于沿岸海域螺类等底栖群落的资源分布, 也多采用阿氏拖网或者采泥器进行样品取样, 前者属于定性/半定量采集网具, 最小网囊网目尺寸25 mm, 网口宽度1.5 m[33];后者虽为定量采样工具, 但其采样面积仅为0.1 m2,对于某些活动能力较强的大型螺类的捕获能力相对较差。本研究所采用的大型单拖网网具采样面积远高于以上两种网具, 最小网囊网目尺寸与阿氏拖网相同, 对大型螺类的捕获能力较强, 因而可以更为准确的反映调查海域螺类资源密度的分布特征。研究结果表明: 螺类平均资源密度(重量)为246.64 kg/km2, 略高于同海域的软体动物资源平均密度(236.00 kg/km2)[34]。从整个调查海域螺类资源密度空间分布看, 总体分布趋势呈现较为明显的斑块分布特征, 但北部平均资源生物量明显低于中南部海域(北部海域平均资源生物量仅为中南部海域的21.80%)。造成以上资源生物量分布特征的主要原因是由于浙江沿岸海域水系及地质类型较为复杂[34], 局部生境异质性较高, 从而造成资源生物量的斑块性分布明显; 而棒锥螺作为中南部海域螺类的最主要种类, 其对螺类平均资源生物量的贡献率达到75.90%, 北部海域主要螺类种类——长尾纺锤螺对对平均资源生物量的贡献率仅为5.49%, 从而造成了平均资源生物量呈现出南高北低的趋势。这一趋势在相同海域的其他生物类群的资源分布中也有所体现[35,36]。
3.2 群落多样性指数
根据研究结果, 春季浙江沿岸螺类群落的三个生物多样性指数均呈偏低水平, 群落处于不稳定状态。这主要是由于某一物种(棒锥螺)高密度集群导致研究水域螺类多样性降低。同时, 春季螺类群落中优势种较少, 且优势度相对群落内其他种类高,也是造成群落生物多样性指数不高的原因之一[37]。从多样性指数的空间分布来看, 3个指数远近岸间差异均不明显, 整体呈现在较低水平下的均匀分布、局部站位具有较高数值的空间特征。造成此种空间分布趋势的主要原因是: 群落多样性的变化不仅与各站位间出现的种类数有关, 还取决于种间个体的均匀性[38], 而生物多样性指数较高的站位中,不仅其种类数相对较多, 而且作为优势种的棒锥螺和褐管蛾螺在这些站位所占的丰度均不超过30%,种间个体分配较为均匀。
3.3 扰动对螺类群落的影响
综合调查海域春季螺类群落ABC曲线特征及W统计值, 根据Clarke和Warwick的划分标准, 表明调查海域春季螺类群落处于中等干扰状态, 而此时群落中生物量占优势的大个体消失, 在数量上占优势的种是个体较小的种, 在此情况下种内丰度的分布与生物量分布优势难分[39]。根据调查中出现的螺类个体大小, 平均体重50 g以上的大型螺类(如管角螺、沟鹑螺等)的生物量仅占螺类总生物量的4.56%, 而平均体重在5 g以下的小型螺类数量则占螺类总数量的93.93%, 充分验证了春季螺类群落处于中等干扰状态这一论断。究其原因, 主要是由于底质的稳定性是衡量其是否适宜包括螺类在内的底栖动物生存的基本条件之一, 底质任何形式的不稳定均可对螺类产生不利影响[40], 而春季(4月份)调查期间正处于浙江近岸渔场的捕捞汛期, 破坏底质较为严重的双拖、单拖以及桁杆拖虾等海洋捕捞作业活动频繁, 加之涉海工程建设和海洋污染等人为因素对底质的破坏[41], 严重影响了螺类群落的生境。
3.4 螺类群落与环境因子的关系
海洋生物的种类分布在时空上呈现一定的异质性, 这与复杂的理化因子和海底地貌关系十分密切[42]。本研究对浙江沿岸海域螺类与主要环境因子间的关系进行了典范对应分析(CCA), 结果表明:调查海域螺类分布受水深的影响最大, 其主要是通过控制螺类的主要食物来源(主要为有机碎屑)的分布来影响栖动物[43]。如分布于图 6左侧的爪哇斗螺、电光螺等种类与水深呈显著正相关, 而在实际调查站位中其恰好分布在水深60m以深的外侧调查, 沿岸调查站位则从未出现; 与之相反, 图 6右侧的浅缝骨螺、棒锥螺等种类与水深呈显著负相关,此类种类在沿岸20 m以浅站位出现个体量占二者总总个体量的百分比达87.63%, 在40 m以深站位则极少出现, 为典型的沿岸种。以上结果表明, 水深梯度是导致浙江沿岸海域群落结构空间变化的主要原因之一。其次, 底层溶解氧同样为影响螺类分布的较为重要的因子。有研究表明: 溶解氧的高低可间接或直接影响大型底栖生物的分布, 过低的溶解氧浓度会限制螺类乃至软体动物的活动范围, 其临界值约为6 mg/L左右[44]。本次调查也印证了此项论断: 所有出现螺类的66个站位中, 其底层溶解氧范围为6.14—9.53 mg/L, 均高于其临界值。如斑鹑螺、脉红螺和长尾纺锤螺等分布在图 6上方的种类, 其分布与底层溶解氧呈显著正相关, 以上种类出现站位的底层溶解氧均在8.50 mg/L, 有效的印证了螺类分布与水中溶解氧分布之间的关系。但是,由于调查项目的局限性, 诸如重金属特别是铜、营养盐、悬浮物、pH、沉积物粒径等环境因子对螺类分布与群落结构的影响未能涉及, 这也有待于进一步的研究。
参考文献:
[1] Guan Q, Liu J P, Wu H T,et al. Researach progress on the ecology of natural wetland snails (Mollusca Gastropoda) in China [J].Acta Ecologica Sinica, 2016, 36(9):2471—2481 [管强, 刘吉平, 武海涛, 等. 中国自然湿地螺类生态学研究进展. 生态学报, 2016, 36(9):2471—2481]
[2] Bronmark C. How do herbivorous freshwater snails af-fect macrophyte: A comment [J].Ecology, 1990, 71(3):1212—1215
[3] Elger A, Lemoine D. Determinants of macrophyte palatability to the pond snailLymnaea stagnalis[J].Freshwater Biology, 2005, 50(1): 86—95
[4] Herman P M J, Middelburg J J, van de Koppel J. Heip CHR (1999) Ecology of estuarine macrobenthos [J].Advances in Ecological Research, 2000, 29: 195—240
[5] Yan J Y, Huang X Y, Chen J K. Study on the species composition and habitats of freshwater snail in the littoral zone of Poyang lake [J].Acta Hydrobiologica Sinica,2014, 38(5): 407—413 [严家跃, 黄心一, 陈家宽. 鄱阳湖沿岸带螺类的物种组成及其生境. 水生生物学报, 2014,38(5): 407—413]
[6] Sun Y J, Wang W, Liu Q G,et al. Spatial distribution of snail (gastropod) community in Yangcheng lake, Jiangsu province, China [J].Resources and Environment in the Yangtze Basin, 2011, 20(6): 711—716 [孙月娟, 王武, 刘其根, 等. 江苏阳澄湖螺类群落的空间分布格局. 长江流域资源与环境, 2011, 20(6): 711—716]
[7] Hu C Y, Xu H, Zou L,et al. Community structure of molluscs in the rock intertidal zone of Zhejiang coast [J].Journal of Fisheries of China, 2015, 39(8): 1144—1154[胡成业, 徐衡, 邹莉, 等. 浙江沿岸岛礁区潮间带软体动物群落结构研究. 水产学报, 2015, 39(8): 1144—1154]
[8] Ji Y L, Zhao N, Wang Z Z,et al. Macrobenthic community structure of intertidal zone of Rushan Bay in spring [J].Chinese Journal of Applied Ecology, 2015,26(2): 609—615 [纪莹璐, 赵宁, 王振钟, 等. 乳山湾潮间带春季大型底栖动物群落结构. 应用生态学报, 2015,26(2): 609—615]
[9] Ke S, Shen Y C, Xie E Y,et al. Biodiversity of the benthic shellfish in the intertidal zone of the Liusha Bay,Leizhou Peninsula [J].Biodiversity Science, 2013, 21(5):547—553 [柯盛, 申玉春, 谢恩义, 等. 雷州半岛流沙湾潮间带底栖贝类多样性. 生物多样性, 2013, 21(5):547—553]
[10] Liu J X. A cataiogue of mollusca on Chuanshan Islands,Taishan City [J].Ecological Science, 1994, (1): 106—108[刘景旋. 台山川山群岛沿岸软体动物名录. 生态科学,1994, (1): 106—108]
[11] Zhang S P, Zhang F S. Species ofThaisfrom China coasts (Gastropoda: Muricidae) [J].Marine Sciences,2005, 29(8): 75—83 [张素萍, 张福绥. 中国近海荔枝螺属的研究(腹足纲: 骨螺科. 海洋科学, 2005, 29(8):75—83]
[12] Zhang S P. Two new records ofNassarius(Gastropoda,Nassariidae) from China Sea [J].Oceanologia Et Limnologia Sinica, 2013, 44(3): 814—816 [张素萍. 中国海织纹螺属Nassarius(腹足纲, 织纹螺科)两新记录种. 海洋与湖沼, 2013, 44(3): 814—816]
[13] Zhang S Q, Zhang J L, Zhang S P. New records of a genus and a species of Buccinidae (Mollusca: Gastropoda) from East China Sea [J].Oceanologia Et Limnologia Sinica, 2015, 46(2): 295—297 [张树乾, 张均龙, 张素萍. 东海蛾螺科(软体动物: 腹足纲)一新记录属和一新记录种. 海洋与湖沼, 2015, 46(2): 295—297]
[14] Hu H Y, Tang J L, Li Q L,et al. Studies on benthic ecology in Zhejiang coastal waters [J].Journal of Marine Sciences, 2006, 24(3): 76—89 [胡颢琰, 唐静亮, 李秋里,等. 浙江省近岸海域底栖生物生态研究. 海洋学研究,2006, 24(3): 76—89]
[15] Zhang J H, Gao Y, Fang H D,et al. Ecological characteristics of macrobenthic communities in Pearl River Estuary [J].Acta Ecologica Sinica, 2009, 29(6): 2989—2999 [张敬怀, 高阳, 方宏达, 等. 珠江口大型底栖生物群落生态特征. 生态学报, 2009, 29(6): 2989—2999]
[16] Zhang P C, Xu Y, Li X Z,et al. Mollusc community of South Yellow Sea in summer [J].Oceanologia Et Limnologia Sinica, 2016, 47(4): 774—784 [张鹏驰, 徐勇, 李新正, 等. 南黄海夏季软体动物群落结构特征. 海洋与湖沼, 2016, 47(4): 774—784]
[17] Cai R X, Huang W H, Liu Y Y,et al. Fauna of Zhejiang Mollusks [M]. Zhejiang: Zhejiang Science and Technology Publishing House. 2001, 4—79 [蔡如星, 黄惟灏, 刘月英, 等. 浙江动物志-软体动物. 浙江: 浙江科学技术出版社. 2001, 4—79]
[18] Liu R Y. Checklist of Marine Biota of China Seas [M].Beijing: Science Press. 2008, 1—1267 [刘瑞玉. 中国海洋生物名录. 北京: 科学出版社. 2008, 654—983]
[19] General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China.Specifications for Oceanographic Survey-Part 6: Marine Biological Survey [S]. Beijing: China Standard Publishing House. 2008, 56—62 [中华人民共和国国家质量监督检验检疫局. 海洋调查规范第 6 部分海洋生物调查.北京: 中国标准出版社. 2008, 56—62]
[20] Pinkas L, Oliphant M S, Iverson L K. Food habits of albacore, bluefin tuna, and bonito in California waters [J].Fishery Bulletin, 1971, 152: 1—105
[21] Zhang H L, Wang Z M, Zhu Z J,et al. Analysis on the community characteristics of shrimps in the coastal spawning ground of South Zhejiang during spring [J].Acta Hydrobiologica Sinica, 2013, 37(4): 712—721 [张洪亮, 王忠明, 朱增军, 等. 浙江南部沿岸产卵场春季虾类群落结构特征分析. 水生生物学报, 2013, 37(4):712—721]
[22] Aglen A, Flyn L, Godflor,et al. Surveys of the marine fish resources of Peninsular Malaysia, June-July 1980[A]. Institute of Marine Research, Bergen, 1981, 9: 1—69
[23] Lu Z H, Xue L J, Zhang L,et al. Assessment of shrimp stock biomass in continental shelf waters of the East China Sea [J].Acta Hydrobiologica Sinica, 2013, 37(5):855—862 [卢占晖, 薛利建, 张龙, 等. 东海大陆架虾类资源量评估. 水生生物学报, 2013, 37(5): 855—862]
[24] Margalef R. Information theory in ecology [J].General Systematics Advancing Frontiers Plant Science, 1957, 3:37—71
[25] Clarke K R, Warwick R M. Change in Marine Communities: an Approach to Statistical Analysis and Interpretation (2nd ed) [M]. Plymouth: PRIMPER-E. 2001, 1—91
[26] ter Braak C J F. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis [J].Ecology, 1986, 67: 1167—1179
[27] Ye J Q, Cai L Z, Huang R J,et al. Species composition of trawling mollusk in Beibu Gulf and its environmental effect [J].Marine Science Bulletin, 2010, 29(6): 617—622[叶洁琼, 蔡立哲, 黄睿婧, 等. 北部湾底拖网软体动物的种类组成及其环境影响. 海洋通报, 2010, 29(6):617—622]
[28] Du F Y, Zhang H H, Li C H,et al. Species composition and diversity of macrobenthic fauna in Daya Bay [J].Journal of Fishery Sciences of China, 2008, 15(2):252—259 [杜飞雁, 张汉华, 李纯厚, 等. 大亚湾大型底栖动物种类组成及物种多样性. 中国水产科学, 2008,15(2): 252—259]
[29] Li B Q, Li X Z, Wang H F,et al. Species diversity of macrobenthic mollusk fauna in Jiaozhou Bay, Shandong[J].Biodiversity Science, 2006, 14(2): 136—144 [李宝泉,李新正, 王洪法, 等. 胶州湾大型底栖软体动物物种多样性研究. 生物多样性, 2006, 14(2): 136—144]
[30] Xu R H, He Z W. Notes on gastropods and bivalvula of Yantai coast and its vicinity [J].Xinxiang Normal College, 1984, (2): 73—79 [许人和, 和振武. 烟台沿海及其附近地区的螺类和双壳类. 新乡师范学院学报, 1984,(2): 73—79]
[31] Jia S H, Zeng J N, Liao Y B,et al. Research on macrozoobenthic community structure in Dongtou Islands and adjacent sea area [J].Journal of Marine Sciences, 2016,34(2): 83—92 [贾胜华, 曾江宁, 廖一波, 等. 洞头列岛及邻近海域大型底栖动物群落结构的研究. 海洋学研究,2016, 34(2): 83—92]
[32] Tian W, Xu Z L. Distribution of quantity and diversity of marrobenthos in the Jiaojiang Estuary [J].Chinese Journal of Applied and Environmental Biology, 2015, 21(2):358—365 [田伟, 徐兆礼. 椒江口海域大型底栖动物数量与多样性的分布特征. 应用与环境生物学报, 2015,21(2): 358—365]
[33] Liu X, Wang L, Gu X Y,et al. Characteristics of the community structure of macrobenthos around Zhejiang Tantoushan island waters in summer and autumn [J].Oceanologia Et Limnologia Sinica, 2017, 48(3): 553—567 [刘迅,王莉, 顾晓英, 等. 浙江檀头山岛周围海域夏、秋季大型底栖动物群落结构特征. 海洋与湖沼, 2017, 48(3):553—567]
[34] Hu H Y, Tang J L, Li Q L,et al. Studies on benthic ecology in Zhejiang coastal waters [J].Journal of Marine Sciences, 2006, 24(3): 76—89 [胡颢琰, 唐静亮, 李秋里,等. 浙江省近岸海域底栖生物生态研究. 海洋学研究,2006, 24(3): 76—89]
[35] Lu Z H, Xue L J, Zhang Y Z. Species composition and quantitative distribution of stomatopod in the East China Sea [J].Journal of Natural Resources, 2013, 28(2):2159—2168 [卢占晖, 薛利建, 张亚洲. 东海口足类(Stomatopod)种类组成和数量分布. 自然资源学报, 2013,28(2): 2159—2168]
[36] Yu C G, Song H T, Ding Y P,et al. A primary estimation on shrimp resource of Zhejiang coastal area [J].Journal of Zhejiang College of Fisheries, 1994, 13(3): 149—155[俞存根, 宋海棠, 丁跃平, 等. 浙江近海虾类资源量的初步评估. 浙江水产学院学报, 1994, 13(3): 149—155]
[37] Ryder R A, Kerr S R, Taylor W W,et al. Community consequences of fish stock diversity [J].Canadian Journal of Fisheries & Aquatic Sciences, 1981, 38(12):1856—1866
[38] Yang G, Li F, Wang X X,et al. Community structure of crabs in the coastal waters along the southern Shandong Peninsula [J].Journal of Fishery Sciences of China, 2017,24(4): 862—874 [杨刚, 李凡, 王秀霞, 等. 山东半岛南部近岸海域蟹类群落结构特征. 中国水产科学, 2017,24(4): 862—874]
[39] Warwick R M. A new method for detecting pollution effects on marine macrobenthic communities [J].Marine Biology, 1986, 92(4): 557—562
[40] Li Y F, Du F Y, Gu Y G,et al. Relationship between macrobenthic fauna community and environmental factors in Southeast Leizhou Peninsula of the South China Sea [J].South China Fisheries Science, 2016, 12(6):33—41 [李亚芳, 杜飞雁, 谷阳光, 等. 雷州半岛东南部海域春季大型底栖动物群落特征及其与环境因素的关系. 南方水产科学, 2016, 12(6): 33—41]
[41] Lin L S, Cheng J H, Ling J Z. Analysis on recent status of the bottom trawl fishery resources in the East China Sea region [J].Marine Fisheries, 2007, 29(4): 371—374 [林龙山, 程家骅, 凌建忠. 东海区底拖网渔业资源变动分析. 海洋渔业, 2007, 29(4): 371—374][42]Ren Y P, Xu B D, Ye Z J,et al. Preliminary study on community structure of fishery resources during spring and autumn in the coastal waters of Qingdao [J].Periodical of Ocean University of China, 2005, 35(5): 792—798[任一平, 徐宾铎, 叶振江, 等. 青岛近海春、秋季渔业资源群落结构特征的初步研究. 中国海洋大学学报, 2005,35(5): 792—798]
[43] Allard M, Moreau G. Effects of experimental acidification on a lotic macroinvertebrate community [J].Hydrobiologia, 1987, 14: 37—49
[44] Chen L Y, Heath A G, Neves R J. Comparison of oxygen consumption in freshwater mussels (Unionidae) from different habitats during declining dissolved oxygen concentration [J].Hydrobiologia, 2001, 450: 209—214
