褐菖鲉的听觉阈值研究
2018-05-16张旭光郭弘艺宋佳坤
张旭光 郭弘艺 宋佳坤
(1. 上海海洋大学海洋动物系统分类与进化上海高校重点实验室, 上海 201306; 2. 上海海大学科技部海洋生物学国际联合研究中心, 上海 201306; 3. 上海海洋大学水产科学国家级实验教学示范中心, 上海 201306)
水下噪声污染是近年来备受关注的环境污染问题, 尤其是来自航运中船舶马达和海上风电机的低频噪声污染日益严重[1]。鱼类可以通过内耳来感知水中的声音信号, 此外, 声信号在其捕食、避害、生殖、洄游、栖息地选择等行为中都有重要的作用[2]。而水下噪声不仅会遮蔽与鱼类上述行为相关的声音线索, 甚至会造成鱼类的听觉丧失和内分泌应激反应等负面影响[3,4], 从而影响其摄食、避害、生殖和洄游等行为, 长期以往可能会影响其种群补充量、群体结构和生态功能等。
褐菖鲉(Sebasticus marmoratus)是典型的近岸岩礁鱼类, 广泛分布于我国东南沿海, 具有较高的生态意义和经济价值[5]。由于航道开发和近岸工程等人为活动的影响, 褐菖鲉的活动范围受到了严重地缩减, 其野生资源量已急剧下降, 急需对褐菖鲉资源开展保护[6]。最近的研究表明, 褐菖鲉可通过鱼鳔肌收缩发出声音进行种间声讯交流[7,8]。这样,近岸水下工程产生的水下噪声以及船舶噪声则可能对其声讯交流造成一定的影响。确定褐菖鲉的听觉敏感性是评估水下噪声对其影响的重要前提。Lee等[9]曾利用行为驯化条件下的心电图技术研究了褐菖鲉的听觉阈值。然而, 这种方法需要实验鱼经过一段长期的训练以形成生理上对声音信号的条件反射, 并不能客观反映实验对象的听觉特性。目前, 听觉诱发位电位(Auditory Evoked Potential, AEP)作为一种微创伤性的电生理技术广泛应用于鱼类的听觉方面的各种研究中[10]。因此, 本文利用听觉诱发电位法研究褐菖鲉的听觉阈值, 以确定其听觉敏感性, 进一步理解其声讯交流机制和环境适应性机制, 并为水下噪声对经济鱼类的影响评估提供科学数据。
1 材料与方法
1.1 实验材料
实验用褐菖鲉为浙江省舟山枸杞岛(30°42′48″N,122°46′42″E)附近海域采用定置网具所捕的渔获物,全长7.3—13.2 cm, 平均(10.3±2.1) cm, 体重30—60 g, 平均(45.3±12) g。实验鱼由船运至实验室暂养, 暂养水槽规格为100 cm×40 cm×45 cm, 水温为22—24℃, 正常自然光照。根据褐菖鲉喜栖岩礁的特性, 暂养水槽内放置多个瓦砾块作为其栖息场所。暂养期间投喂人工配合颗粒饲料以及小杂鱼。实验前, 挑选无外伤, 有活力的褐菖鲉进行实验。实验前采用0.1%的MS-222麻醉3—5min, 并在其躯干背部注射加拉碘铵抑制肌肉活动, 剂量为2—4 μg/g(体重), 仅见鱼鳃盖轻微扇动。
1.2 AEP记录
实验鱼通过自制的夹持器固定在实验水槽中(r=15 cm), 并通过自重力富氧流水进行人工灌流,以保持实验鱼的呼吸[11,12]。水槽放置于位于隔音室(2.5 m×2.5 m×2 m, 上海声诺)内的防震台(ZDT07-07, 江西连胜)上。隔音室内的空气本底噪声为25 dB(上海声诺)。
实验时, 使鱼头部背侧露出水面1—2 mm, 记录电极(Rochester Electro-Medical, 美国)置于头骨的中缝线上, 延脑上方。参考电极置于记录电极前方1—2 cm处。2个电极穿过纸巾紧紧地固定于皮肤上。另有一根接地电极置于鱼尾附近的水体中。所有电极均直接连接至前置放大器(RA4LI,Tucker Davis Technologies TDT, 美国), 并通过光纤与TDT听觉生理工作站(RZ6, 内置数模转换和信号同步, TDT, 美国)相连。
声音刺激文件由软件SigGenRZ (TDT, 美国)设置。频率设置为100、150、200、300、500、800、1000 Hz。声音时程(Duration)为12ms, 含2ms起落(rase/fall), 以减少声音输出的瞬变。声音信号通过功率放大器(D75A, Crown, 英国)输至位于水槽底部的水下扬声器(UW-30, University Sound, 美国)。实验鱼距离水下扬声器 18 cm, 水槽内的实际声音信号, 由置于鱼耳附近的水听器(8103, B&K, 丹麦)进行实时测量。测量的声音信号通过电荷放大器(2692, B&K, 丹麦)输入TDT数模转换模块。实验中采集的AEP信号经放大器, 也输入TDT数模转换模块, 并由软件BigSigRZ (TDT, 美国)同步保存声音信号与AEP信号。
实验鱼听觉阈值确定方法与Kenyon等相似[11,13]。实验时设置2个相位相反的刺激声音, 分别为90°和270°, 如果它们诱发的脑电位波形具有重合性, 即可视为正确的AEP反应。声音刺激强度设置为从110到60 dB, 每次以3 dB递减, 直至不可产生可重复的AEP波形, 此时对应的刺激声音的声压级即为该频率的听觉阈值[14,15]。实际刺激声的声压级通过水听器记录的声音波形和水听器校正系数共同确定。
2 结果
2.1 褐菖鲉AEP波形特征
实验对8条褐菖鲉进行了AEP测试。2个相位相反的刺激声音诱发的AEP波形可以基本重合(图1), 而且2个AEP波形之间的自相关系数(R值)也较高。但随着刺激强度的降低, AEP波形的自相关系数(R值)也逐渐降低。在刺激强度较大时, 其R值在0.7—0.9内波动, 当接近听觉阈值时, 其R值急剧下降, 但波形仍表现一定的反应幅度。AEP反应的波形幅度也随着刺激强度的降低而减弱, 通常反应幅度降至10—20 μV左右时, 基本也接近听觉阈值。由于AEP反应具有与声音相位无关的特性[10,16], 如果AEP波形不能重合时, 即无AEP反应, 其R值则可能接近或低于零。
根据波形形状特点, 褐菖鲉AEP波形可分为3类(图 2)。一类是100 Hz低频信号诱发的AEP波形, 波形相对于基线波动较平缓; 第二类是150—300 Hz诱发的AEP波形, 首先是产生1个相对于基线的负向峰, 在这个负向峰上可能出现多个小的负向峰, 紧接着1个相对基线的正向峰; 第三类是500—800 Hz诱发的AEP波形首先是一个相对于基线的正向起伏, 紧接着一个缓慢的负向峰。
褐菖鲉AEP波形的另一个基本特征是AEP反应频率几乎是刺激频率的两倍。如图 3所示, 150 Hz的声音信号在水槽中的实际频率为143 Hz, 其对应的AEP波形的频谱中则存在具有二倍关系的287 Hz频率峰, 而另一个597 Hz的频率峰则可能来自声音信号中262 Hz的声音畸变信号。这种两倍关系的特点也可简单的从两者波形疏密程度看出(图 2)。
2.2 褐菖鲉听力图
实验共记录了8尾褐菖鲉对100—1000 Hz短纯音刺激的AEP反应。将8尾褐菖鲉在各测量频率下获得的听觉阈值平均后, 可得出褐菖鲉的听力曲线图(图 4, AEP)。根据听力曲线图, 褐菖鲉的听力曲线在100—1000 Hz的频率范围内呈线性上升, 听觉阈值随声音频率的增加而增加, 即随着声音频率的增加, 褐菖鲉听觉敏感性降低。褐菖鲉对低频声音信号较敏感, 主要频率范围100—300 Hz, 其听觉阈值为70—79 dB, 其中最敏感的频率为150 Hz, 对应的听觉阈值为(70±1.1) dB, 对500 Hz以上的声音敏感性较差, 其听觉阈值已达90 dB以上。
3 讨论
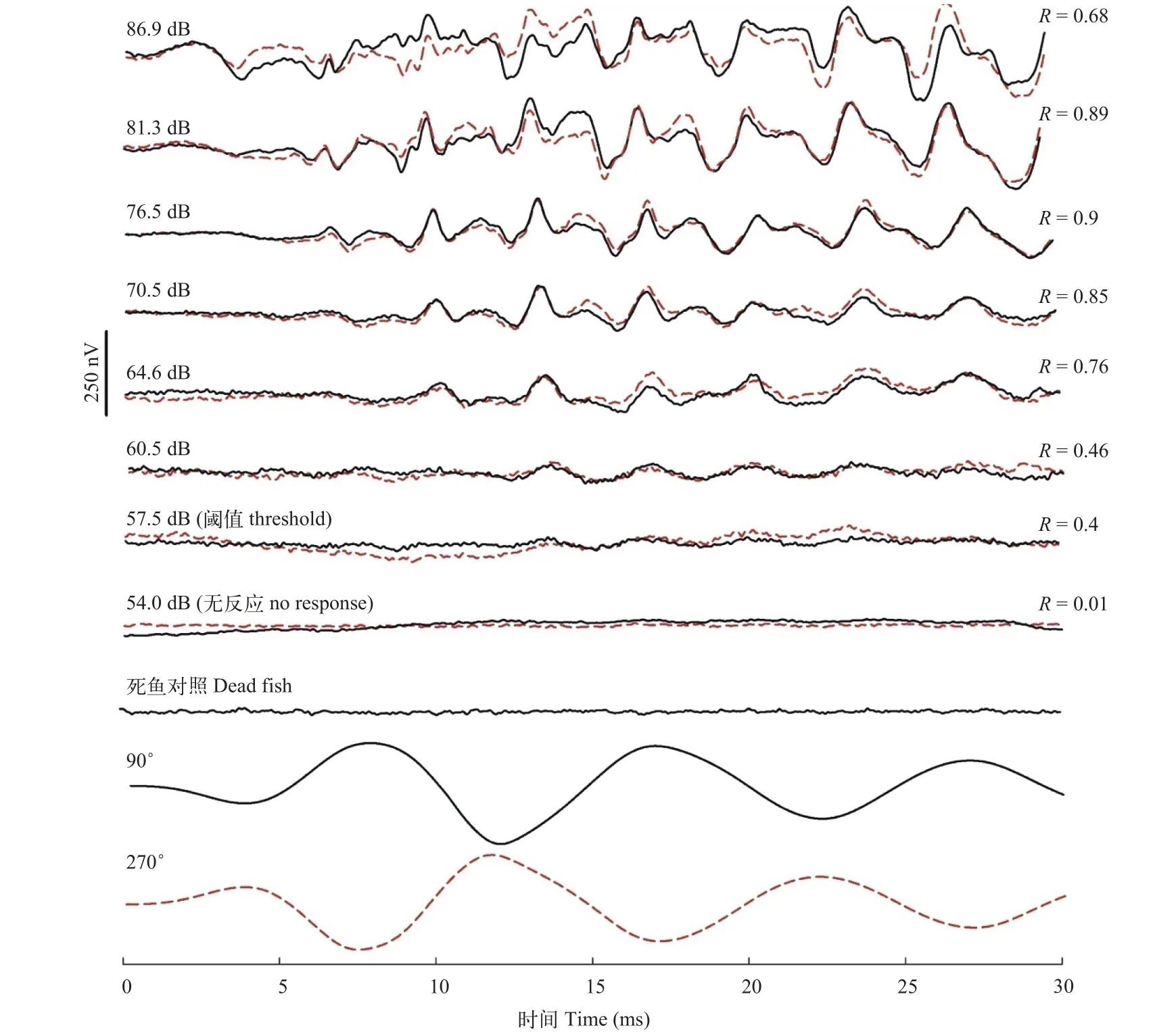
图 1 不同相位和刺激强度诱发的褐菖鲉AEP波形Fig. 1 AEP wave forms of marbled rockfish in response to 100 Hz tone bursts at stimulus levels from 86.9 dB to 54 dB with starting phases of 90° and 270°, respectively
目前, 听觉诱发电位法(AEP)是世界公认的最快、最准确的确定鱼类听觉阈值的方法[14]。相对传统的行为条件反射方法或其它创伤性电生理法,AEP方法无需对动物长期驯化或进行组织破坏性的解剖, 具有快速、可重复试验的优点[13—17]。本文首次应用听觉脑干诱发电位法研究了鲉形目鱼类褐菖鲉的听觉阈值。确定动物的电位变化是否为AEP反应的方法是采用相位相反的二组声音刺激,如果诱发波形表现出与刺激声音相位无关的波形重合性, 则表明该电位变化为正确的AEP波形。波形重合性可以通过第二实验人经肉眼检测或通过波形自相关系数进行检测。理论上来说, 相位相反的声音信号互相叠加后可消除AEP波形中声音信号的干扰[16]。然而, 由于实验条件限制, 如实验水槽的谐振, 声音的反射等, 声音信号叠加后并不能完全消除, 依然存在一些声音信号的残迹, 但声音伪迹的特点是波形起伏相对于基线是平行的。而AEP波形在叠加后, 表现出相对于基线起伏的正向或负向峰(图 2), 这与Kenyon等[13]在金鱼(Carassius auratus)听觉阈值研究中的结果相似。但是, 褐菖鲉在低频段的AEP波形相对基线起伏较平缓, 只有在高频段的AEP波形才有相对基线的负向或正向起伏。Lu等[17]在金鱼的听觉阈值研究中发现, 100 Hz的纯音诱发的AEP波形也相对基线较平缓, 只有500 Hz时, 才出现相对于基线起伏显著的波峰。在鱼类中, 一般只有高频短纯音可以诱发较好的听觉脑干电位, 这可能与听觉神经元的频率响应有关,但具体的神经机制还尚不清楚[18]。另外, 鱼类对声音或机械波的感受主要涉及声压感受或微粒子运动位移感受。对于听觉敏感型鱼类, 通过鱼鳔与内耳的连接(或相对靠近)提高了声压感受的敏感性,也提高的听觉阈值的上限[19]。因此, 听觉敏感型的鱼类的听觉上限可达数千赫兹。缺乏内耳与鱼鳔联系的鱼类则对低频的微粒子运动位移更敏感, 而无鳔鱼类则只对微粒子位移敏感[20,21]。可见, 听觉脑干诱发电位更适用于声压感受鱼类的听觉阈值研究, 而低频声音信号除声压信号外还涉及更多的粒子位移信号[21]。因此, 低频声音信号诱发的听觉脑干电位波形表现出非典型的AEP特征。AEP反应的另一特征是其反应频率接近刺激频率的二倍,这与听觉毛细胞成对排列有关[22]。然而, 在金鱼的听觉研究中也发现, 当声音刺激高于2000 Hz时, 这一规律并不是很显著, 其AEP的反应频率则小于刺激频率的二倍值[17], 其中的机制尚待进一步研究。
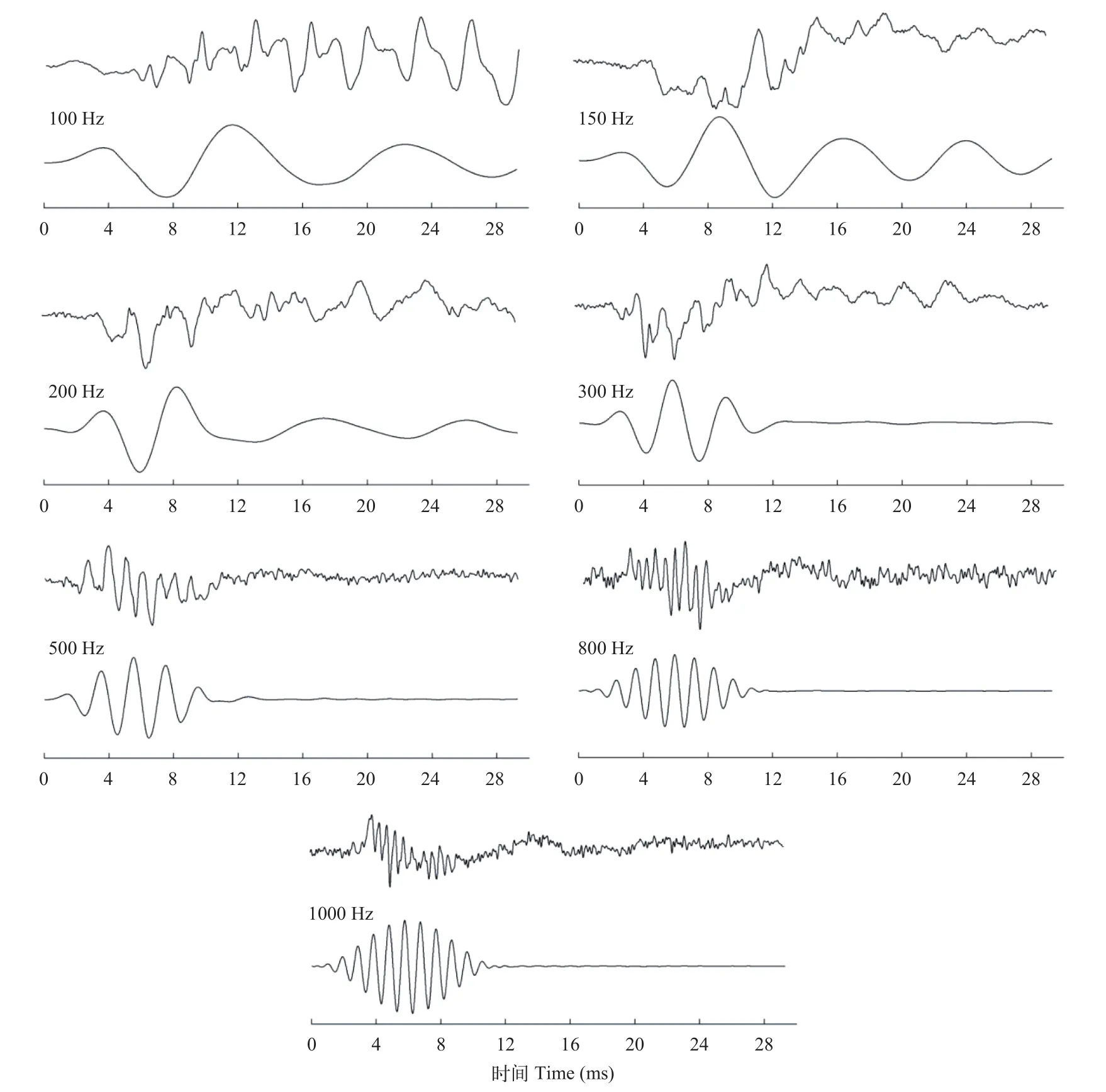
图 2 不同频率刺激声音的诱发的褐菖鲉AEP波形Fig. 2 AEP wave forms of marbled rockfish in response to tone bursts with different frequencies
目前, 对鲉科鱼类听觉阈值的研究较少。Lee等[9]通过行为驯化条件反射的心电记录对褐菖鲉的听觉阈值进行了研究, 其行为听力曲线与听觉诱发电位法所得到的褐菖鲉听力曲线基本相似。行为学检测结果表明, 褐菖鲉其敏感的频率范围为80—300 Hz, 其中对100 Hz声音最为敏感, 对应的听觉阈值为90 dB。对300 Hz以上声音敏感性较差,阈值已达120 dB以上(图 4, ECG)。而AEP实验中,褐菖鲉的敏感频率范围为100—300 Hz, 其中150 Hz为最敏感频率, 对应阈值为70 dB, 比行为听觉阈值低20 dB。Kojima等[23]利用行为学和AEP方法对真鲷(Pagrus major)的听觉阈值进行了对比研究, 结果表明, 两种方法得到的听力曲线形状相同, 但行为学听觉阈值却比AEP听觉阈值高15 dB左右。同样,对听觉普通型鱼类眼斑星丽鱼(Astronotus ocellatus)的比较研究表明, 行为听觉阈值比AEP听觉阈值要高10 dB以上[13]。虽然, 不同实验室在不同条件下所得的听觉阈值存在差异, 但是不同方法所体现的频率敏感性却基本相近。根据行为学和AEP实验结果(图 4), 可知褐菖鲉对低频声音信号敏感, 对500 Hz以上的声音敏感性较差。
褐菖鲉可通过鱼鳔肌收缩发出主频为(172±2.5) Hz左右的低频信号, 用于种间的声讯交流[7,24]。AEP实验表明褐菖鲉听觉敏感性在100—300 Hz,这种发声频率与听觉敏感性的高度匹配性, 表明了其感觉系统协同进化的适应性[25]。然而, 由于海洋航运和水下工程使海洋中的低频噪声污染日益严重, 如海上风电场运营期的水下噪声主要频率为125 Hz[26], 而船舶马达声的水下噪声也以低频为主,声压级可高达150 dB以上[27,28]。这样, 水下噪声污染则可能对褐菖鲉的声讯交流造成影响。因此, 亟需开展海洋中近岸人为噪声对褐菖鲉个体或群体行为影响的相关研究, 以保护褐菖鲉的野生资源进而实现对该资源的持续利用。

图 3 150 Hz刺激声音(A)和褐菖鲉AEP波形(B)的频谱Fig. 3 The FFT of (A) 150 Hz sound stimulus and (B) AEP wave form of marbled rockfish

图 4 褐菖鲉听力曲线比较Fig. 4 Audiograms of the S. marmoratus from methods AEP and electrocardiogram (ECG), respectively[9]
参考文献:
[1] Hildebrand J A. Anthropogenic and natural sources of ambient noise in the ocean [J].Marine Ecology Progress Series, 2009, 395(5): 5—20
[2] Fay R. Soundscapes and the sense of hearing of fishes [J].Integrative Zoology, 2009, 4(1): 26—32
[3] Popper A, Hastings M. The effects of anthropogenic sources of sound on fishes [J].Journal of Fish Biology,2009, 75(3): 455—489
[4] Kight C R, Swaddle J P. How and why environmental noise impacts animals: an integrative, mechanistic review[J].Ecology Letters, 2011, 14(10): 1052—1061
[5] Fujita H, Kohda M. Timing and sites of parturition of the viviparous scorpionfish,Sebastiscus marmoratus[J].Environmental Biology of Fishes, 1998, 52(1): 225—229
[6] Zhang S Y, Wang L, Wang Z H,et al. Characteristics of dominant fish assemblages in kelp bed habitat of Gouqi Island and distribution diversity in coastal area [J].Journal of Fisheries of China, 2011, 35(9): 1399—1409 [章守宇, 王蕾, 汪振华, 等. 枸杞岛海藻场优势种鱼类群体特征及其在不同生境中的差异. 水产学报, 2011, 35(9):1399—1409]
[7] Zhang X, Guo H, Zhang S,et al. Sound production in marbled rockfish (Sebastiscus marmoratus) and implications for fisheries [J].Integrative Zoology, 2015, 10(1):152—158
[8] Zhang X, Guo H, Wang Z,et al. Noise-Dependent Fish Distribution in Kelp Beds [A]. In: Popper A N, Hawkins A (Eds.), The Effects of Noise on Aquatic Life II [C].Springer New York. 2016, 1259—1264
[9] Lee C, Park Y, Moon J,et al. The hearing ability of the scorpion fish sebastiscus marmoratus to audible sound, 1.the auditory threshold [J].Bulletin of the Korean Society of Fisheries Technology, 1999, 35(2): 156—160
[10] Kenyon T, Ladich F, Yan H. A comparative study of hearing ability in fishes: the auditory brainstem response approach [J].Journal of Comparative Physiology A,1998, 182(3): 307—318
[11] Zhang X, Herzog H, Song J,et al. Response properties of the electrosensory neurons in hindbrain of the white sturgeon,Acipenser transmontanus[J].Neuroscience Bulletin, 2011, 27(6): 422—429
[12] Zhang X, Song J, Fan C,et al. Use of electrosense in the feeding behavior of sturgeons [J].Integrative Zoology,2012, 7(1): 74—82
[13] Yan H, Curtsinger W. The otic gasbladder as an ancillary auditory structure in a mormyrid fish [J].Journal of Comparative Physiology A Sensory Neural & Behavioral Physiology, 2000, 186(6): 595—602
[14] Ladich F, Fay R R. Auditory evoked potential audiometry in fish [J].Reviews in Fish Biology and Fisheries, 2013,23(3): 317—364
[15] Yan H Y, Anraku K, Babaran R P. Hearing in marine fish and its application in fisheries [J].Behavior of MarineFishes: Capture Processes and Conservation Challenges,2010: 45—64
[16] Corwin J T, Bullock T H, Schweitzer J. The auditory brain stem response in five vertebrate classes [J].Electroencephalography & Clinical Neurophysiology, 1982,54(6): 629—641
[17] Lu Z, Tomchik S M. Effects of a red-tide toxin on fish hearing [J].Journal of Comparative Physiology A, 2002,188(10): 807—813
[18] Lu Z. Neural mechanisms of hearing in fishes [A]. In:Von Der Emde G, Mogdans G J, Kapoor B G (Eds.), The Senses of Fish: Adaptations for the Reception of Natural Stimuli [C]. Boston: Kluwer Academic Publishers. 2004,147—172
[19] Yan H Y. The Role of Gas-Holding Structures in Fish Hearing: An Acoustically Evoked Potentials Approach[A]. In: von der Emede G, Mogdans J (Eds.), The Senses of Fish [C]. New Delhi: Narosa Publishing House. 2003,189—209
[20] Fine M L, King C B, Cameron T M. Acoustical properties of the swimbladder in the oyster toadfishOpsanus tau[J].Journal of Experimental Biology, 2009, 212(21):3542—3552
[21] Radford C A, Montgomery J C, Caiger P,et al. Pressure and particle motion detection thresholds in fish: a re-examination of salient auditory cues in teleosts [J].The Journal of Experimental Biology, 2012, 215(19): 3429—3435
[22] Tanimoto M, Ota Y, Inoue M,et al. Origin of inner ear hair cells: morphological and functional differentiation from ciliary cells into hair cells in zebrafish inner ear [J].The Journal of Neuroscience, 2011, 31(10): 3784—3794
[23] Kojima T, Suga T, Kusano A,et al. Acoustic pressure sensitivities and effects of particle motion in red sea breamPagrus major[J].Fisheries Science, 2010, 76(1):13—20
[24] Zhang X G, Xie W, Guo H Y,et al. Sound characteristics and morphology of sonic system of the Marbled rockfishSebasticus marmoratus[J].Journal of Shanghai Ocean University, 2013, 22(5): 672—677 [张旭光, 谢伟, 郭弘艺, 等. 褐菖鲉发声系统形态结构及其发声特征研究.上海海洋大学学报, 2013, 22(5): 672—677]
[25] Ladich F, Popper A N. Parallel evolution in fish hearing organs [A]. In: Manley G A, Fay R R, Popper A N (Eds.),Evolution of the Vertebrate Auditory System [C]. New York: Springer. 2004, 95—127
[26] Tougaard J, Henriksen O D, Miller L A. Underwater noise from three types of offshore wind turbines: Estimation of impact zones for harbor porpoises and harbor seals[J].Journal of Acoustical Society of America, 2009,125(6): 3766—3773
[27] Amoser S, Wysocki L, Ladich F. Noise emission during the first powerboat race in an Alpine lake and potential impact on fish communities [J].Journal of the Acoustical Society of America, 2004, 116(6): 3789—3797
[28] Picciulin M, Sebastianutto L, Codarin A,et al. In situ behavioural responses to boat noise exposure ofGobius cruentatus(Gmelin, 1789; fam. Gobiidae) andChromis chromis(Linnaeus, 1758; fam. Pomacentridae) living in a Marine Protected Area [J].Journal of Experimental Marine Biology & Ecology, 2010, 386(1-2): 125—132
