鲫幼鱼游泳运动能力的个体变异与表型关联
2018-05-16曾令清付世建
曾令清 付 成 付世建
(重庆师范大学进化生理与行为学实验室, 动物生物学重庆市重点实验室, 重庆 401331)
游泳运动是鱼类逃逸、捕食、繁殖等活动行为的重要实现方式, 为其生存生长及种群繁衍提供了基本保证。依据对氧气的需求, 鱼类游泳运动分为有氧运动和无氧运动2种类型, 前者反映有氧代谢能力, 与日常活动(如巡游、自发运动)和洄游等有关, 常以临界游泳速度(Ucrit)[1,2]或步法转换速度(Ugt)评价[2], 而后者则反映无氧代谢能力, 与捕食活动、逃逸敌害、穿越激流等相关, 常用匀加速游泳速度(Ucat)或爆发游泳速度评估[3]; 鱼类的有氧和无氧运动能力可能存在权衡[3—5]。自Ucat用于衡量鱼类无氧运动能力起, 不同研究采用加速度均是0.167 cm/s2[即10 (cm/s)/min][3—8]。采用单一加速度考察不同鱼类Ucat的方式可能具有局限性, 因为不同鱼类的生物学特征(如发育阶段、体重大小、游泳能力及性别等)差异较大, 将影响游泳能力的测定结果。因此, 不同加速度与鱼类Ucat关系的相关研究十分迫切。
鱼类表型存在种内个体变异并且在不同环境条件下维持稳定, 具有重要进化生物学和生态学意义[9]。鱼类处于静止状态、无食物消化吸收时的最低能量消耗称作为静止代谢率(Resting metabolic rate, RMR), 代表了机体基本功能的维持水平和有氧代谢能力的下限, RMR与标准代谢率(Standard metabolic rate, SMR)有相近的内涵, 本研究统一使用RMR; 高强度有氧运动对鱼类施加巨大的生理负荷, 使其对氧的需求达到最大, 表现出最大代谢率(Maximum metabolic rate, MMR), 代表了机体有氧代谢能力的上限, 二者的差值为鱼类的有氧代谢空间(Aerobic scope, AS=MMR–RMR), 它表示为其他生理功能提供的最大代谢潜能[10]。鱼类MMR和AS的测量方法通常包括两种, 一种是Ucrit法[11—14],别一种则是人为诱导运动力竭的过量耗氧(Excess post-exercise oxygen consumption, EPOC)法[11,15,16]。2种方法的本质都是通过高强度游泳运动对机体施加最大生理负荷, 诱导机体产生最强有氧代谢水平。然而, 由于实验方法的操作性及其相应的生态学意义不尽相同, 比较不同研究结果时可能面临一定的方法学局限性, 有关鱼类有氧代谢能力特征诱导方法的比较研究十分匮乏[11]。
鲫(Carassius auratusLinnaeus)是我国常见的鲤科鱼类, 广泛分布于江河湖泊、池塘沼泽等水域中。为考察鲤科鱼类运动能力的个体变异和表型特征的生态关联, 以及不同加速度对匀加速能力测定的影响, 本研究以鲫幼鱼为实验对象, 通过2种方法分别测定该种鱼的有氧代谢参数(RMR、MMR、AS及FAS)和有氧运动能力(Ucrit), 同时也测量在不同加速度(0.083、0.167、0.250、0.333 cm/s2)条件下鲫幼鱼的Ucat, 在Ucrit和Ucat测定过程中分别录像以分析获取相应的Ugt, 分析比较鲫幼鱼的有氧和无氧运动能力特征及表型关联, 比较匀加速游泳能力在不同加速度条件下的响应特征, 为鱼类匀加速游泳能力测定加速度的合理选择提供有益参考, 同时也为我国过渔设施(如鱼道)工程的设计开发提供重要的运动生理学资料。
1 材料与方法
1.1 实验鱼及其驯化
鲫幼鱼购自于重庆市北碚区歇马人工养殖基地, 运回后将鱼放置于3个实验室循环控温水槽(约250 L)中驯化3周。驯化期间, 每天以商业饲料(中国通威公司)饱足投喂1次, 投喂0.5h后用虹吸法清除残饵和粪便以维持水体环境。驯化水槽的日换水量约为水体的10%, 实验用水为曝气后控温的自来水, 用充气泵不断向驯化水槽充气使溶氧水平大于7 mg/L, 驯化水温为(25.0±0.5)℃(与当时季节水温相近), 光周期为光照14 L∶10 D。
1.2 实验设计
在驯化结束后, 挑选鱼体健康、体重相近的鲫幼鱼[n=30, 体重(13.08±0.41) g, 体长(7.52±0.08) cm]作为实验对象, 并将实验鱼移入已编号的30个养殖单元格中以便辨别实验鱼。实验之前, 对鱼禁食24h。在第1天先测量实验鱼的RMR, 随后采用游泳代谢仪测定鱼的Ucrit、Ugt和MMR并计算获取AS,测定完毕后将鱼放回至养殖单元格中并投喂一次约0.5%体重的饵料[17], 以维持实验鱼的体重和生理恢复; 第3天测定EPOC法诱导鲫幼鱼的MMR并在运动力竭后连续测定鱼的恢复代谢率, 共计算10min的过量耗氧(EPOC), 在测定完毕后将鱼放回至养殖单元格中并投喂一次约0.5%体重的饵料; 从第5天开始随后的连续4d中, 于每天上午只测定一个加速度条件下实验鱼的Ucat和Ugt, 本研究设定的加速度包括0.083、0.167、0.250、0.333 cm/s2, 每天测定完毕后对单尾鱼投喂一次约0.5%体重的饵料。对于单尾鱼而言, 在Ucrit和EPOC测定时均有48h机体恢复时间, 在不同加速度条件下测量Ucat的恢复时间也达24h, 并在每次饵料投喂过程中所有实验鱼全部表现快速摄食, 但由于体重控制要求仅对每尾鱼投喂约0.5%体重的饵料。根据研究报道,在运动力竭1h后鱼类白肌的三磷酸腺苷和磷酸肌酸恢复到运动前水平[17], 力竭运动后8h后机体的乳酸水平也恢复运动前水平[18]。本研究实验参数测定时间的间隔明显长于上述重要能量物质恢复和产物清除时间, 不影响鲫幼鱼游泳能力的测定。实验期间所有环境条件(如温度、溶氧、光周期等)与驯化期间保持一致。
1.3 RMR的测定
本研究采用流水式呼吸代谢仪测定实验鱼的RMR。将鱼从养殖单元格转移至呼吸代谢仪的呼吸室驯化24h, 以消除转移过程带来的影响。驯化结束后, 采用溶氧仪(HQ30d, 美国哈希公司)在第2天的9:00、11:00、13:00、15:00、17:00、19:00和21:00测定实验鱼的耗氧率。呼吸代谢仪的环境条件(如水温、溶氧和光周期等)与驯化期间保持一致;测定过程中实验室保持安静, 以减少声音对实验鱼生理状态造成影响。将7个时间点的耗氧率取平均值用来表征实验鱼的RMR [mg O2/(kg.h)], 计算公式如下:
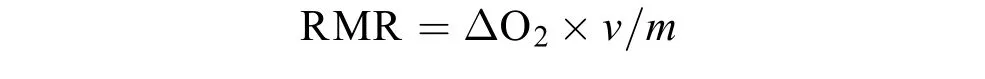
式中, ΔO2是实验呼吸室(有鱼)出水口与空白呼吸室(无鱼)出水口的溶氧差值(mg O2/L),v是呼吸室流速(L/h), 由呼吸室出水口装满50 mL容量瓶所需时间计算获得,m为实验鱼的体重(kg)。由于实验鱼的体重十分相近, 无需进行代谢率的体重校正。
1.4 Ucrit和MMR的测定
(1)Ucrit测定法: 采用鱼类游泳代谢仪同时测定实验鱼的运动代谢率和Ucrit, 同时通过摄像系统拍摄实验鱼的游泳行为。先将实验鱼装入游泳代谢仪的游泳管中驯化1h, 驯化流速为6 cm/s; 随后每隔20min增加6 cm/s, 直至实验鱼运动力竭。力竭标准: 实验鱼因运动力竭不能抵抗水流而后退停靠于游泳管的尾端筛板, 并且在尾端筛板的停靠时间达20s以上[19]。Ucrit(cm/s)的计算公式如下:
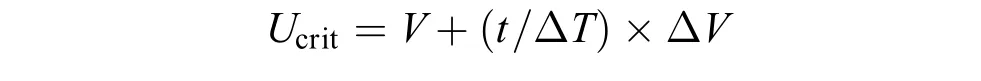
式中,V是实验鱼能够完成20min游泳运动的最大流速, ΔV是流速增量(6 cm/s), ΔT是时间间隔(20min),t是在最高流水速度下实验鱼的游泳历时(持续时间短于20min)。
在测定Ucrit过程中, 同时测量实验鱼的运动代谢率。游泳代谢仪装有1个连接溶氧仪外部水循环系统(流量约为500 mL/min), 当溶氧仪探头连接该系统时游泳代谢仪形成封闭状态, 可监测游泳代谢仪内部水体溶氧水平。每隔20min, 打开游泳仪谢仪尾盖20s随后关闭, 该时间足以保证代谢仪内部与外部之间水体的充分置换, 溶氧水平可迅速恢复至正常状态。通过设定好的电脑程序控制目标流速, 用溶氧仪每隔2min测定一次游泳代谢仪内部水体的溶氧水平(mg/L), 连续测定10次。实验鱼的运动代谢率[MO2, mg O2/(kg.h)]的计算公式如下:
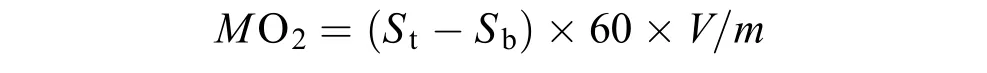
式中,St[mg O2/(L.min)]为每段期间(20min)代谢仪内部水体溶氧水平随时间变化的斜率, 当回归方程的指数R2>0.95时的斜率, 即可参与MO2的计算。60为60min,Sb[mg O2/(L.min)]为在1 h期间游泳代谢仪内部细菌耗氧的斜率。V为扣除实验鱼体积后的代谢仪内部的水体体积(3.50 L),m为实验鱼的体质量(kg)。在测定过程中, 获取的最大MO2即为实验鱼的, 其代谢空间=–RMR。
在Ucrit测定过程中, 在游泳管正上方安置一台高清摄像机, 录像单尾鱼Ucrit游泳行为。随流速的逐渐增加, 由于鱼类开始动用白肌(无氧代谢)提供游泳动力而打破体内乳酸平衡, 使得体内的乳酸积累一定程度时, 鱼类开始出现“爆发–滑行”的游泳行为变化并且摆尾频率变化明显, 该游泳行为被称为步法转换, 首次出现“爆发–滑行”游泳行为时的游泳速度称作为步法转换速度(Ugt), 评价最大有氧运动能力[2,20]。虽然Ucrit测定过程中涉及到无氧代谢且饱受争议, 但现有研究仍采用Ucrit或Ugt评价鱼类的有氧运动能力[2]。
(2) EPOC测定法: 通过力竭运动诱导实验鱼的MMREPOC[mg O2/(kg.h)]并计算代谢空间(ASEPOC=MMREPOC–RMR)。EPOC的简要步骤[15]: 将鱼从养殖单元格中转移至力竭运动驱赶装置(Φ外=56 cm,Φ内=33 cm, 水深15 cm)的环行水道(流速~60 cm/s)中; 在环形水道中实验鱼逆流快速游泳, 一段时间后鱼体因高流速无法保持相对稳定游泳而后退, 此时人工连续驱赶实验鱼向前游泳直至其运动力竭;处于力竭状态的鱼通常失去平衡且用手接触尾部时不再有明显应激反应, 整个过程一般在5min之内完成[21]; 然后将鱼迅速(10s之内完成)转移至呼吸代谢仪的呼吸室中进行30min的恢复代谢测定。溶氧数据采集时间分别为0、1、2、3、4、5、6、7、8、9、10、15、20、25、30min。在恢复代谢测定过程中, 呼吸室流速约为500 mL/min。经计算, 在此流速条件下呼吸室水体置换率达99%的历时小于1min[22]。因此, 将随后数分钟出现的最大耗氧率作为实验鱼的MMR, 其计算公式与RMR一样。鱼类的EPOC是指其在运动恢复期内超过静止状态耗氧水平的额外耗氧量, 在一定程度上反映了鱼类的最大无氧代谢能力, 计算恢复期内高于运动前的差值在时间上的总和即为该时期的EPOC[15]。为便于比较不同鲫个体的代谢恢复特征, 本研究仅选取前10min的EPOC用于数据分析。
1.5 Ucat的测定
本研究共采用了4个加速度(0.083、0.167、0.250、0.333 cm/s2), 每天只测定1个加速度下实验鱼的Ucat。将鱼放入游泳代谢仪的游泳管中驯化1h, 管内流速为6.0 cm/s。驯化结束后, 流速从6.0 cm/s开始按照某一加速度(每秒种递增)逐渐增加游泳代谢仪的流速, 直至实验鱼运动力竭, 该力竭状态被认为是鱼类的最大匀加速游泳能力, 此时对应的流速即为实验鱼的Ucat。游泳代谢仪的流速控制由电脑软件完成, 可即时观察游泳管中即时流速及其变化特征。在测定过程中, 在游泳管正上方安装一台高清摄像机, 录像单尾鱼整个的匀加速游泳过程。对获取的视频资料分析确定实验鱼首次“爆发–滑行”游泳行为的出现时间并结合摆尾频率的变化特征, 以确定在Ucat测定过程中实验鱼的Ugt。
1.6 数据处理与统计分析
采用Excel (2003)软件对所有实验数据作常规计算, 统计软件为SPSS (19.0)软件。实验数据均以“平均值±标准误”(Means±SE)表示, 显著水平为P<0.05。Ucrit测定中的能量代谢参数与游泳能力的关系及Ucat与无氧代谢(Ucat–Ugt)的关系均采用一般线性回归方程。两种方法诱导MMR和AS的比较采用T-检验, 能量代谢参数之间的相关性采用Pearson相关分析。采用单因素方差分析考察不同加速度对游泳能力参数的影响, 若组间存在显著差异则采用最小显著性差异法(Least significant difference,LSD)检验组间数据差异。采用组内相关系数(Intraclass correlation coefficient, ICC)和肯德尔和谐系数(Kendall concordance coefficient, KCC)考察匀加速游泳能力参数在不同加速度条件下的重复性。
2 结果
2.1 Ucrit中能量代谢与游泳速度的关系
在Ucrit法中鲫幼鱼的MMR和AS均与Ucrit呈正相关, 但RMR与Ucrit不相关(图 1A)。此外, 鲫幼鱼的能量代谢参数(MMR、AS、RMR)与Ugt不相关(图 1B, 所有P>0.05)。
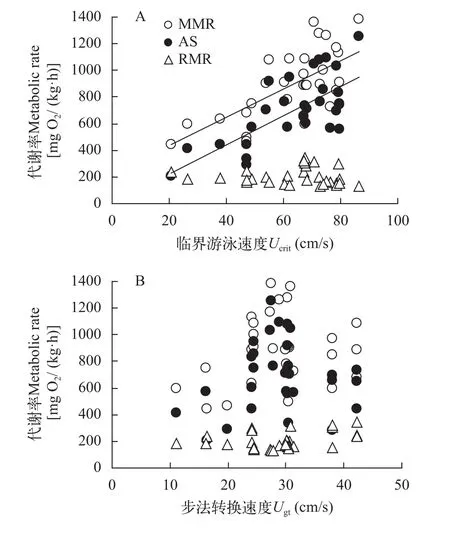
图 1 鲫幼鱼能量代谢与游泳速度的关系Fig. 1 The relationships between metabolic rates and swimming velocity in juvenile crucian carp
2.2 两种方法中鲫幼鱼能量代谢参数的比较及相关性
本研究采用了2种常规诱导鱼类最大代谢率的方法, 分别为Ucrit法和EPOC法。结果发现, 两种方法获取鲫幼鱼的MMR和AS均无显著性差异(图2A, MMR:T=0.200,P=0.843; AS:T=0.220,P=0.827); 同样, 2种方法获得的FAS (Ucrit法:4.84±0.38; EPOC法: 4.98±0.36)也无明显差异(T=0.593,P=0.558)。虽然2种方法获得能量代谢参数的平均值无差异, 但2种方法的能量代谢参数(MMR、AS和FAS)仍保持较高的重复性(图 2B,MMR:r=0.434,P=0.017; 图 2C, AS:r=0.493,P=0.006; 图 2D, FAS:r=0.808,P<0.001)。
在Ucrit法中, 鲫幼鱼的RMR与MMR及AS均不相关, 但与FAS呈负相关; MMR与AS及FAS呈正相关, AS与FAS也呈正相关(表 1)。在EPOC法中, 鲫幼鱼的RMR与MMR不相关, 但与AS、FAS、EPOC均呈负相关, MMR与AS、FAS、EPOC均呈正相关(表 1)。AS、FAS、EPOC三者之间呈正相关(表 1)。
2.3 鲫幼鱼的有氧和无氧运动能力
鲫幼鱼的Ucrit低于不同加速度条件下的Ucat, 但加速度对Ucat测定无显著影响(图 3A,F=9.333,P<0.001)。然而, 随加速度的增加,Ugt呈现先增加后下降的变化规律(图 3B,F=87.386,P<0.001)。与Ugt类似, 运动能力比例(Ugt/Ucrit或Ugt/Ucat)也呈现先递增后降低的变化趋势(图 3C,F=58.797,P<0.001)。
不论是Ucrit法还是Ucat法, 鲫幼鱼在两方法中的Ugt均与Ucrit或Ucat呈现正相关, 并且在不同加速度条件下Ugt与Ucat呈现的正相关方向保持不变(表2)。此外, 鲫幼鱼的匀加速游泳能力参数[Ucat,Ugt,Ugt/Ucat,Ucat–Ugt, (Ucat–Ugt)/Ucat]在不同加速度条件下均保持较高的重复性(表 3)。
从加速度0.083 cm/s2逐渐增加至0.250 cm/s2,鲫幼鱼Ucat的无氧代谢能力(Ucat–Ugt)与Ucat之间均呈现正相关(图 4A-C, 所有P<0.01), 而在加速度0.333 cm/s2下二者不相关(图 4D,r=0.342,P=0.069)。在匀加速能力测定中, 鲫幼鱼的有氧代谢能力(Ugt)与无氧代谢能力(Ucat–Ugt)呈负相关, 即二者存在权衡(图 5,r= –0.607,P<0.001)。
3 讨论
3.1 鲫幼鱼有氧运动能力与能量代谢的关系
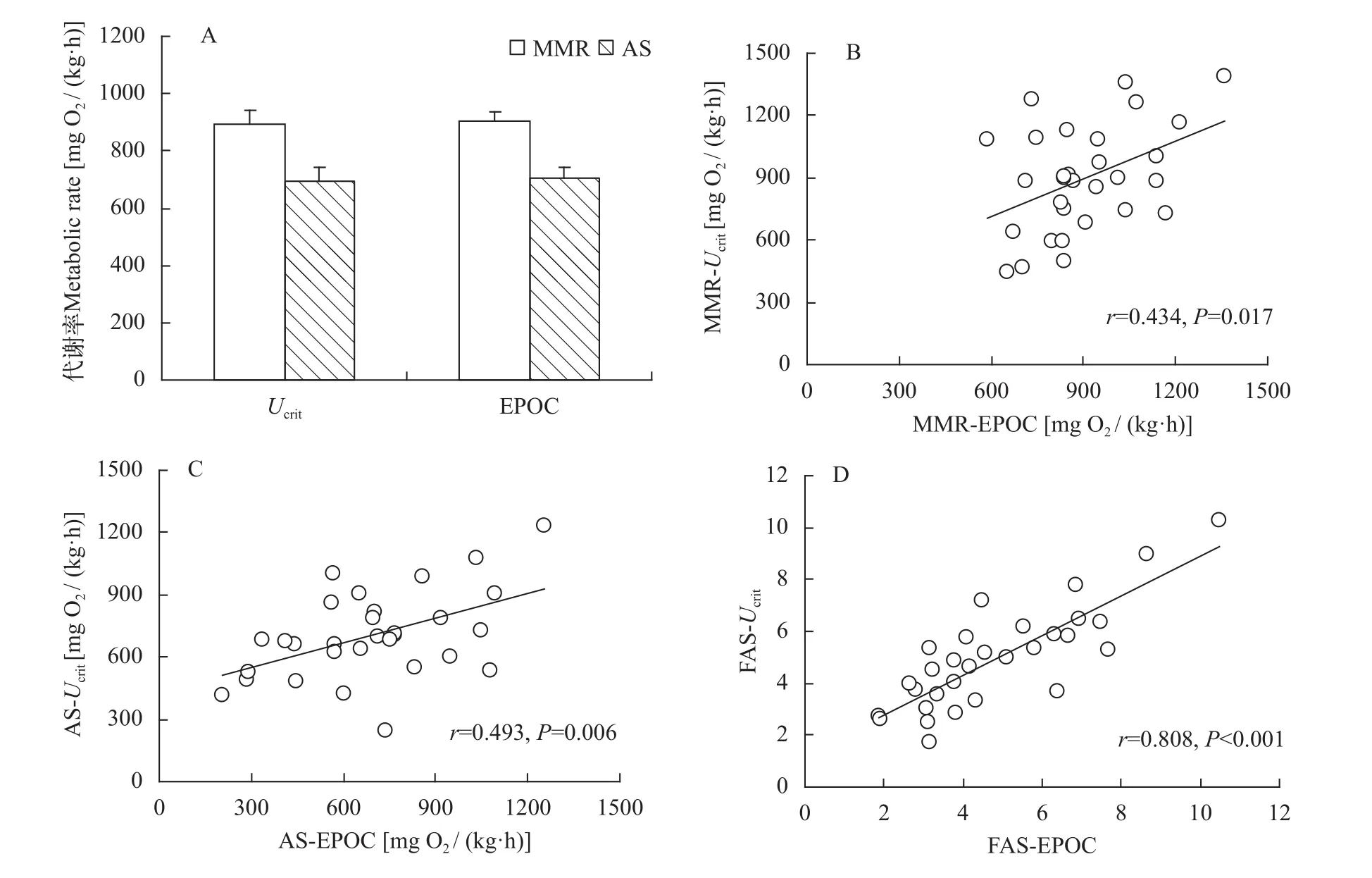
图 2 两种运动方式诱导鲫幼鱼有氧代谢能力的比较及相关性Fig. 2 The comparison and relationship in aerobic capacities induced by two methods in juvenile crucian carp
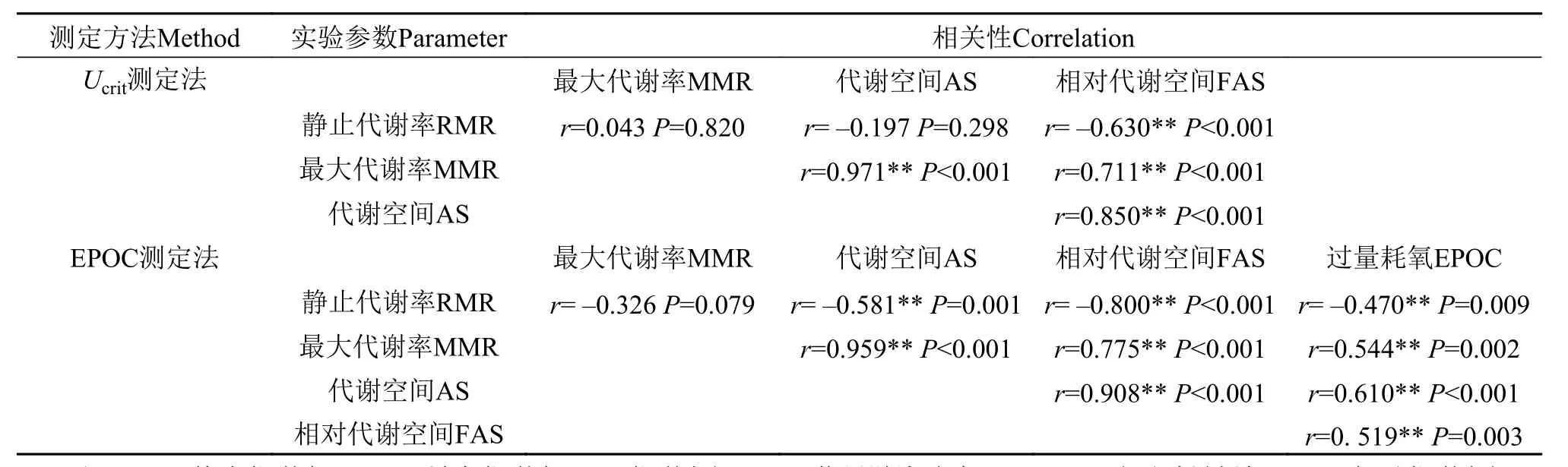
表 1 两种方法中鲫幼鱼能量代谢参数之间的相关性Tab. 1 Correlations among parameters of metabolic rate induced by two exercised methods in juvenile crucian carp
随着游泳速度逐渐增加至Ucrit时, 鱼类呼吸循环系统的生理功能处于最大负荷状态, 表现出最大有氧代谢水平, 常用来评价鱼类的有氧运动能力[23]。本研究发现, 鲫幼鱼的运动代谢率随游泳速度的增加而呈指数性上升, 并且其MMR和AS与Ucrit均呈现正相关, 即有氧代谢能力越强的个体可以支撑更强水平的有氧运动, 反之亦然。这种现象不仅存在于鲤科鱼类的种间水平[13], 而且也发现于鲤科鱼类的种内水平[7,24]。本研究还发现鲫幼鱼的RMR与Ucrit不相关, 这与鲤(Cyprinus carpioLinnaeus)[7]和青鱼(Mylopharyngodon piceusRichardson)[24]的正相关报道并不一致。在Ucrit测定过程中, 鲫幼鱼的Ucrit与Ugt呈正相关, 但Ugt与该种鱼的MMR、AS及RMR均不相关, 这可能是因为鲫幼鱼首次出现“爆发–滑行”游泳行为时(Ugt)的有氧代谢水平低于Ucrit时的MMR, 并且这两个代谢水平间的差距因个体而异, 具有明显个体变异; 另一方面由于游泳速度与有氧代谢之间的拟合方程指数不尽相同, 即相同游泳速度的增加, 指数越大的鲫个体其有氧代谢上升更为明显[11,19]。
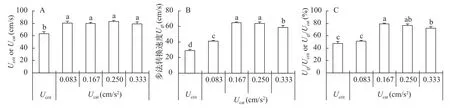
图 3 鲫幼鱼游泳能力参数的比较Fig. 3 The comparisons between parameters of swimming performance in juvenile crucian carp under two exercised methods
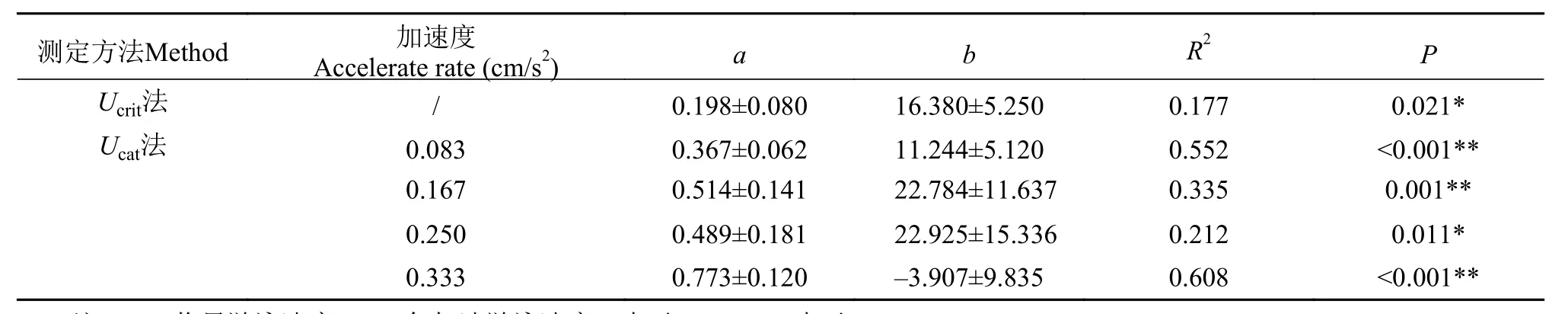
表 2 鲫幼鱼最大游泳能力与步法转换速度的回归方程Tab. 2 Regressions (±SE) of gait transition speed (x) as a function of swimming velocity (y, Ucrit or Ucat), y=a x+b
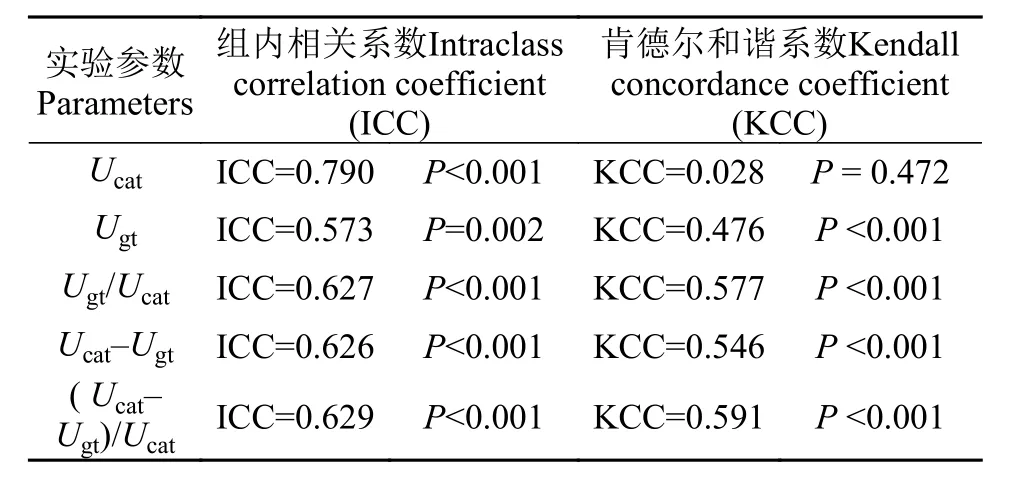
表 3 鲫幼鱼匀加速游泳能力参数的重复性Tab. 3 Repeatability of parameters of acceleration swimming performance in juvenile crucian carp
本研究采用了Ucrit法和EPOC法诱导鲫幼鱼的MMR和AS, 发现Ucrit法中MMR和AS的平均值与EPOC法的十分相近且无明显差异。然而, 这只是在组间水平的平均值差异检验, 还可以在组内个体水平上对两种方法进行比较。为此, 本研究将Ucrit法的能量代谢参数(MMR、AS和FAS)分别与EPOC法的进行重复性分析。结果发现, 3个能量代谢参数在两种方法之间保持较高的重复性, 即个体的生理表型保持较强的可重复测量水平并且在种群中的表型排序维持稳定, 表明在排除环境因素(如水温、体重和溶氧)后, 不论Ucrit法和EPOC法诱导鱼类的最大有氧代谢能力均可采用并进行比较。值得注意的是, 不论Ucrit法还是EPOC法均是排除实验鱼消化道中食物消化吸收的影响, 但先前研究发现鱼类最大代谢率的诱导与其代谢模式有关,食物的消化吸收和游泳运动同时进行的方式可诱导某些鱼类(如鲤)的最大代谢率[23], 这种由2种生理活动叠加诱导的最大代谢率明显大于单一生理活动(运动或消化)的诱导值。因此, 在方法采用时还需要考虑实验鱼的代谢模式类型。
3.2 鲫幼鱼RMR与MMR或AS的关系
RMR是鱼类机体的最小维持代谢水平, 而MMR或AS代表了机体的最大有氧代谢能力, 二者的关系引起研究者的广泛关注[25]。如果自然选择倾向于动物既具有较低的能量维持消耗又具有较强的最大代谢能力的表型特征组合, 那么拥有这种最佳表型组合的动物在自然界中具有极高的适合度[25],然而这种最佳表型关联的进化结果可能是不存在的[26,27]。根据内温动物有氧代谢能力的进化理论,自然选择在倾向于选择个体拥有较强的最大有氧代谢能力同时也会倾向于选择较高的维持代谢, 因为具备较强功能的器官(如发达的呼吸循环系统和运动系统)的机体相应具有更多收益(即具有更高的最大代谢率), 但这些器官也具有相应的代价(即相应较高的能量消耗)[28]。因此, 维持代谢消耗与最大代谢率在理论上将呈正相关。支持该假说的鱼类实验证据已见报道, 例如鲤[7]、青鱼[24]和鳟(Salmo truttaLinnaeus)[25]的RMR与MMR呈现正相关。
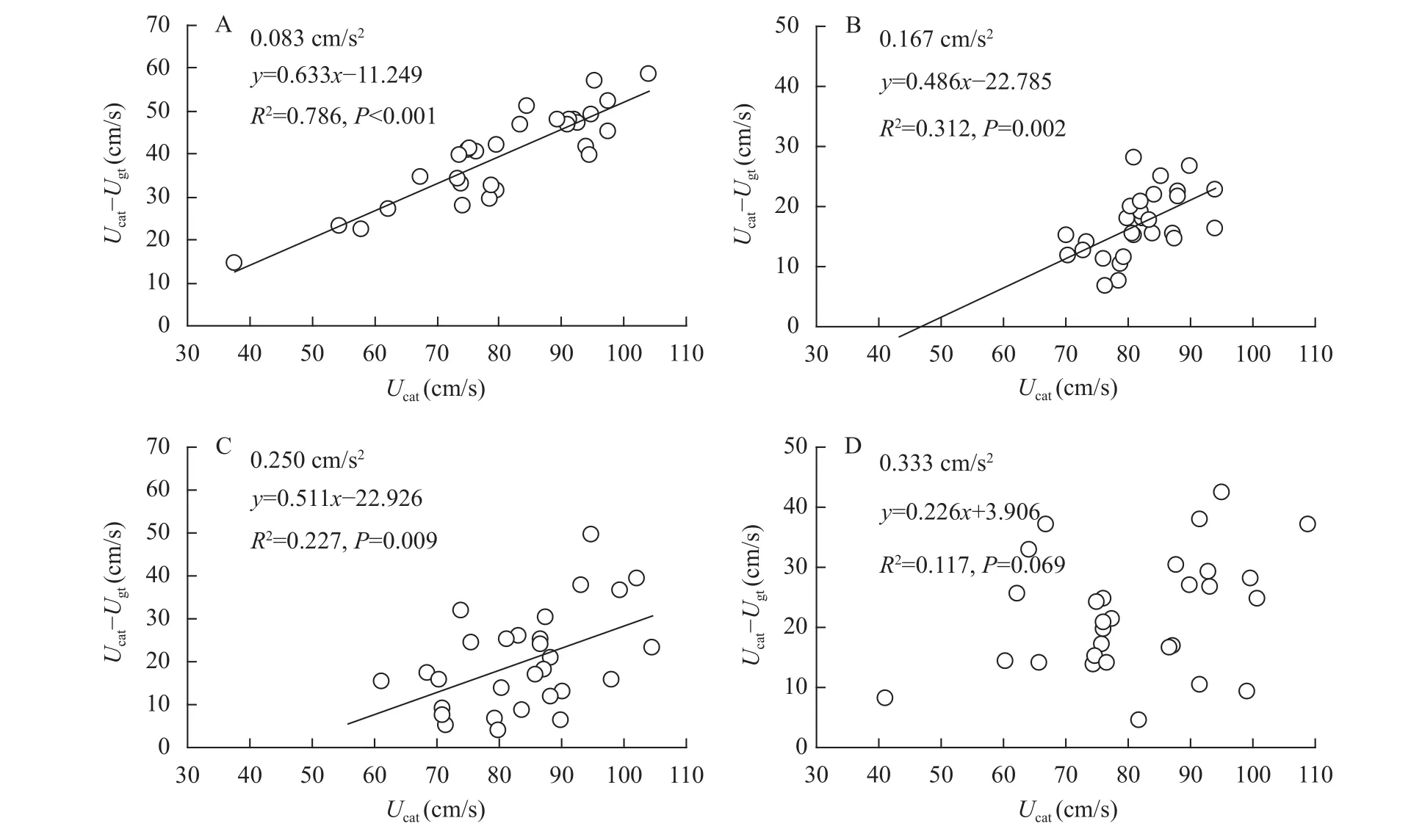
图 4 不同加速度条件下鲫幼鱼匀加速游泳能力与无氧代谢的关系Fig. 4 Relationships between acceleration swimming performance and anaerobic metabolism in juvenile crucian carp at different accelerated rates
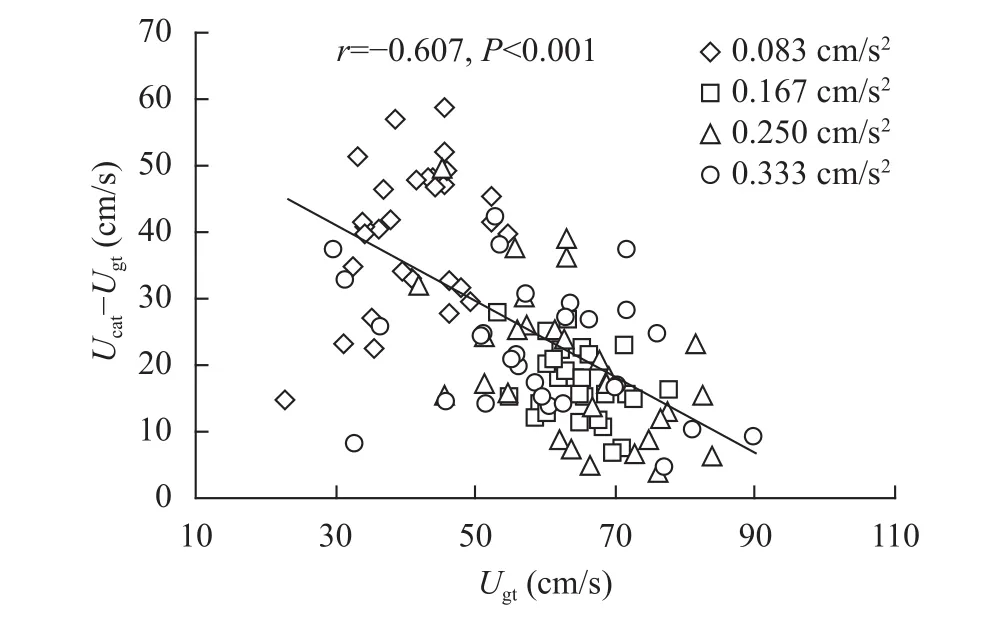
图 5 鲫幼鱼的有氧代谢能力(Ugt)与无氧代谢能力(Ucat–Ugt)的相关性Fig. 5 Relationship between aerobic metabolic capacity and anaerobic metabolic capacity in juvenile crucian carp
对于鱼类种内RMR与MMR呈正相关而言, 在排除体重差异和发育因素外迄今还未找到与之有关器官形态学的证据, 但在生化水平上发现证据:如鳟肝脏上两种有氧代谢关键酶的活性与RMR及MMR均呈正相关[25]。然而, 也有研究发现南方鲇(Silurus meridionalisChen)的RMR与MMR及AS不相关[14]; 欧鲈(Dicentrarchus labraxLinnaeus)[5]、鲫[15]和鳊(Parabramis pekinensisBasilewsky)[16]的RMR与MMR不相关, 与AS呈负相关; 大西洋鲑(Salmo salarLinnaeus)的RMR与FAS呈现负相关[29],暗示该种鱼的RMR与MMR也不相关; 上述研究表明不同鱼类的研究结果差异甚大, 但可总结为两种类型: 一是正相关, 二是不相关。本研究发现, 两种运动法中鲫幼鱼RMR与MMR均不相关, EPOC法中RMR与AS呈负相关, 符合第二种相关类型。在自然界中, 假定自然选择会使鱼类同时具有较强的有氧代谢能力和较高的代谢维持, 并且较强的有氧代谢能力有助于个体的生存、生长和繁衍[30], 理论上拥有该表型个体会获得较高的适合度。因此, 在长进化时间尺度上, 自然选择结果是该鱼类种群的有氧代谢能力表型峰值更高且表型维度变异较窄。然而, 这种理论推测并未得到野外和实验室观测数据的支持, 因为野外和实验室表型特征的个体变异仍然较大[3—5]。另一方面, 在实验室条件下可以获取鱼类的最大有氧代谢能力, 但无法确定鱼类在野外条件下最大有氧代谢能力的使用频率[31], 该功能表型受到自然选择的强度及其与生活史的关联的研究也十分困难, 这也可能是导致维持代谢与最大代谢不相关的原因之一。
3.3 鲫幼鱼的匀加速能力及加速度影响
鱼类匀加速游泳能力的评价研究最早见于2000年报道, 该研究采用的加速度为0.167 cm/s2[10 (cm/s)/min][3], 但该研究并未对加速度的采用原因进行解释, 即加速度的影响被忽视。本研究发现在4个加速度(0.083、0.167、0.250、0.333 cm/s2)下鲫幼鱼的匀加速能力Ucat不受加速度的影响。同样, 虹鳟(Oncorhynchus mykissWalbaum)[32]和鳊幼鱼的Ucat在较大加速度范围内也保持不变[33,34], 与本研究结果类似。前期研究发现在较小加度速下的Ucat介于Ucrit与较大加速度下Ucat之间[33,34], 即加速度越小,Ucat值越接近于Ucrit, 本研究鲫幼鱼Ugt也具有相似变化规律, 该现象可能是因为在较小加速度条件下实验鱼在大部分时间进行有氧运动,Ucrit很大程度上反映了鱼类有氧代谢能力并且可被认为是最小匀加速的特例, 鱼类的低匀加速游泳是一种非常接近有氧运动的游泳类型, 因而低加速度下Ucat值接近Ucrit值[33]。然而, 从另一方面考虑, 如果以过大加速度考察鱼类的匀加速游泳能力, 加速度越快, 实验鱼的运动力竭时间相应越短[33], 那么过高的加速度将影响鱼类匀加速游泳能力的准确评估, 因为过快频率的流速变化可能导致实验鱼提前停止游泳而造成假的运动力竭现象。值得注意的是, Farrell研究的加速度递增时间为每分钟[32], 本研究中及实验室前期研究[33,34]的加速度递增时间为每秒钟。以每秒钟递增的加速度的设定方式对鱼类匀游泳运动的引导作用更为积极连贯, 可更好反映鱼类的匀加速游泳能力, 同时降低了因流速的突然陡增对鱼产生的生理胁迫影响。
研究认为, 鱼类Ucat包括有氧代谢组分(Ugt)和无氧代谢组分(Ucat–Ugt), 并且2个组分间可能存在权衡, 因为前者是由红肌有氧代谢供能而后者是白肌无氧代谢供能[4,5]。本研究发现鲫幼鱼的Ugt与Ucat–Ugt呈现负相关, 并且Ucat与Ucat–Ugt呈正关, 表明该种鱼在匀加速游泳能力过程中有氧代谢和无氧代谢之间存在权衡。大西洋鳕(Gadus morhuaLinnaeus)的研究数据支持上述权衡观点[3], 但欧鲈的实验结果并不支持[4,5]。上述权衡观点在人类运动员研究中找到了肌肉形态结构的支持证据, 即短跑运动员的腿部快肌(无氧代谢)拥有比例高于慢肌(有氧代谢), 而马拉松运动员的腿部慢肌(有氧代谢)拥有比例高于快肌(无氧代谢)[35]。同样, 大部分鱼类的肌肉组成也包括常见的红肌和白肌两种类型, 二者的解剖学结构差异明显, 是上述功能权衡的形态学基础[36]。随着游泳速度的逐渐增加, 鱼类的游泳行为特征发生相应变化, 例如: 在较高游泳速度下稳定游泳的控制程度下降, 摆尾幅度和频率增加[20]及“爆发–滑行”行为的发生频率上升[10]; 在生化角度上, 无氧代谢大量参与的证据在于鱼类体内重要器官(如肌肉、肝脏和血液)的乳酸水平明显上升, 但是在静止状态下也保持一定的乳酸水平[18,33,34], 代谢底物和产物含量的个体差异可能有助于解释上述权衡现象。然而, 验证上述权衡的不同研究结果出现分歧, 仍有必要结合形态学与生化水平进一步探索权衡现象背后的内在机制。
综上所述, 本研究采用2种运动方法诱导鲫幼鱼有氧代谢能力的观测值不论在组间水平还是个体水平均无方法学的差异, 并且能量代谢参数之间存在表型关联; 加速度对鲫幼鱼的Ucat测定无影响,建议选择较高的加速度, 加速度对Ugt测定有影响,在不同加速度条件下鲫幼鱼的匀加速游泳能力保持较高的重复性, 并且有氧代谢与无氧代谢存在权衡。
参考文献:
[1] Plaut I. Critical swimming speed: its ecological relevance[J].Comparative Biochemistry and Physiology A, 2001,131(1): 41—50
[2] Peake S J. Gait transition speed as an alternate measure of maximum aerobic capacity in fishes [J].Journal of Fish Biology, 2008, 72(3): 645—655
[3] Reidy S P, Kerr S R, Nelson J A. Aerobic and anaerobic swimming performance of individual Atlantic cod [J].Journal of Experimental Biology, 2000, 203(2): 347—357
[4] Marras S, Claireaux G, McKenzie D J,et al. Individual variation and repeatability in aerobic and anaerobic swimming performance of European sea bass,Dicentrarchus labrax[J].Journal of Experimental Biology, 2010,213(1): 26—32
[5] Marras S, Killen S S, Domenici P,et al. Relationships among traits of aerobic and anaerobic swimming performance in individual European sea bassDicentrarchus labrax[J].PLoS One, 2013, 8(9): e72815
[6] Pang X, Fu S J, Li X M,et al. The effects of starvation and re-feeding on growth and swimming performance of juvenile black carp (Mylopharyngodon piceus) [J].Fish Physiology and Biochemistry, 2016, 42(4): 1203—1212
[7] Pang X, Fu S J, Zhang Y G. Acclimation temperature alters the relationship between growth and swimming performance among juvenile common carp (Cyprinus carpio) [J].Comparative Biochemistry and Physiology A,2016, 199: 111—119
[8] Penghan L Y, Pang X, Fu S J. The effects of starvation on fast-start escape and constant acceleration swimming performance in rose bitterling (Rhodeus ocellatus) at two acclimation temperatures [J].Fish Physiology and Biochemistry, 2016, 42(3): 909—918
[9] Burton T, Killen S S, Armstrong J D,et al. What causes intraspecific variation in resting metabolic rate and what are its ecological consequences [J]?Proceedings of the Royal Society B, 2011, 278(1724): 3465—3473
[10] Norin T, Malte H. Repeatability of standard metabolic rate, active metabolic rate and aerobic scope in young brown trout during a period of moderate food availability[J].Journal of Experimental Biology, 2011, 214(10):1668—1675
[11] Pang X, Yuan X Z, Cao Z D,et al. Analysis of power curve model on swimming locomotion in qingbo (Spinibarbus sinensis) [J].Journal of Chongqing Normal University(Natural Sciences), 2012, 29(6): 26—29 [庞旭, 袁兴中, 曹振东, 等. 中华倒刺鲃幼鱼的游泳运动功率曲线模型分析. 重庆师范大学学报(自然科学版), 2012,29(6): 26—29]
[12] Yang Y, Cao Z D, Fu S J. Effects of water temperature on the critical swimming speed and metabolic scope of juvenileParabramis pekinensis[J].Chinese Journal of Ecology, 2013, 32(5): 1260—1264 [杨阳, 曹振东, 付世建. 温度对鳊幼鱼临界游泳速度和代谢范围的影响. 生态学杂志, 2013, 32(5): 1260—1264]
[13] Yan G J, He X K, Cao Z D,et al. An interspecific comparison between morphology and swimming performance in cyprinids [J].Journal of Evolutionary Biology,2013, 26(8): 1802—1815
[14] Zeng L Q, Penghan L Y, Wang J W,et al. Repeatability and variation in locomotion metabolism of the juvenile southern catfish during starvation [J].Journal of Fisheries of China, 2015, 39(1): 52—64 [曾令清, 彭韩柳依,王健伟, 等. 南方鲇幼鱼运动代谢的个体变异和重复性及其饥饿响应. 水产学报, 2015, 39(1): 52—64]
[15] Zeng L Q, Fu S J, Cao Z D. Correlations between standard metabolic rate and metabolic characteristics after exhaustive exercise, and behaviour in the juvenile crucian carp (Carassius auratus) [J].Acta Ecologica Sinca, 2016,36(6): 1747—1755 [曾令清, 付世建, 曹振东. 鲫幼鱼(Carassius auratus)标准代谢的个体差异与力竭后代谢特征及行为的关联. 生态学报, 2016, 36(6): 1747—1755]
[16] Zeng L Q, Fu C, Xi L,et al. Phenotypic correlations and individual variation of energy metabolism and personality in juvenile Chinese bream (Parabramis pekinensis)[J].Acta Ecologica Sinca, 2017, 37(14): 4807—4816 [曾令清, 付成, 奚露, 等. 鳊幼鱼能量代谢和个性行为的个体变异及表型关联. 生态学报, 2017, 37(14): 4807—4816]
[17] Milligan C L, Wood C M. Tissue intracellular acid-base status and the fate of lactate after exhaustive exercise in the rainbow trout [J].Journal of Experimental Biology,1986, 123(2): 123—144
[18] Li L, Cao Z D, Fu S J. Lactate, glycogen and glucose levels of juvenile catfish (Silurus asotusLinnaeus) in response to exhaustive exercise [J].Acta Hydrobiologica Sinica, 2007, 31(6): 127—132 [李黎, 曹振东, 付世建. 力竭性运动后鲇鱼幼鱼乳酸、糖原和葡萄糖水平的变动.水生生物学报, 2007, 31(6): 127—132]
[19] Lee C G, Farrell A P, Lotto A,et al. Excess post-exercise oxygen consumption in adult sockeye (Oncorhynchus nerka) and coho (O. kisutch) salmon following critical speed swimming [J].Journal of Experimental Biology,2003, 206(18): 3253—3260
[20] Killen S S, Marras S, Steffensen J F,et al. Aerobic capacity influences the spatial position of individuals within fish schools [J].Proceedings of The Royal Society B,2012, 279(1727): 357—364
[21] Kieffer J. Limits to exhaustive exercise in fish [J].Comparative Biochemistry and Physiology A, 2000, 126(2):161—179
[22] Steffensen J F. Some errors in respirometry of aquatic breathers: how to avoid and correct fro them [J].Fish Physiology and Biochemistry, 1989, 6(1): 49—59
[23] Fu S J, Cao Z D, Zeng L Q,et al. Fish Locomotionstrategy and Adaptive Evolution [M]. Beijing: Science Press. 2014, 73—150 [付世建, 曹振东, 曾令清, 等. 鱼类游泳运动-策略与适应性进化. 北京: 科学出版社. 2014,73—150]
[24] Pang X, Fu S J, Zhang Y G. Individual variation in metabolism and swimming performance in juvenile black carp(Mylopharyngodon piceus) and the effects of hypoxia [J].Marine and Freshwater Behaviour and Physiology, 2015,48(6): 431—443
[25] Norin T, Malte H. Intraspecific variation in aerobic metabolic rate of fish: relations with organ size and enzyme activity in brown trout [J].Physiological and Biochemical Zoology, 2012, 85(6): 645—656
[26] Hammond K A, Diamond J. Maximal sustained energy budgets in humans and animals [J].Nature, 1997,386(6624): 457—462
[27] Hayes J P, Garland Jr. The evolution of endothermy: testing the aerobic capacity model [J].Evolution, 1995,49(5): 836—847
[28] Bennett A F, Ruben J A. Endothermy and activity in vertebrates [J].Science, 1979, 206(4419): 649—654
[29] Cutts C J, Metcalfe N B, Taylor A C. Juvenile Atlantic salmon (Salmo salar) with relatively high standard metabolic rates have small metabolic scopes [J].Functional Ecology, 2002, 16(1): 73—78
[30] Claireaux G, Handelsman C, Standen E,et al. Thermal and temporal stability of swimming performance in the European sea bass [J].Physiological and Biochemical Zoology, 2007, 80(2): 186—196
[31] Peake S J, Farrell A P. Locomotory behaviour and postexercise physiology and oxygen consumption in relation to swimming speed and gait transition in free-swimming smallmouth bass (Micropterus dolomieu) [J].Journal ofExperimental Biology, 2004, 207(9): 1563—1575
[32] Farrell A P. Comparisons of swimming performance in rainbow trout using constant acceleration and critical swimming speed tests [J].Journal of Fish Biology, 2008,72(3): 693—710
[33] Wang J W, Cao Z D, Fu S J. The constant acceleration swimming performance in juvenile Chinese bream (Parabramis pekinensis) [J].Journal of Chongqing Normal University(Natural Science), 2015, 32(2): 13—16 [王健伟, 曹振东, 付世建. 鳊鱼匀加速游泳能力的研究. 重庆师范大学学报(自然科学版), 2015, 32(2): 13—16]
[34] Wang J W, Cao Z D, Fu S J. A comparison of constant acceleration swimming speeds when acceleration rates are different with critical swimming speeds in Chinese bream under two oxygen tensions [J].Fish Physiology and Biochemistry, 2016, 42(5): 1453—1461
[35] Komi P V. Physiological and biochemical correlates of muscle function: effects of muscle structure and stretchshortening cycle on force and speed [J].Exercise and Sport Sciences Reviews, 1984, 12(1): 81—121
[36] Seebacher F, Walter I. Differences in locomotor performance between individuals: importance of parvalbumin,calcium handling and metabolism [J].Journal of Experimental Biology, 2012, 215(4): 663—670
