Harnessing migraines for neural regeneration
2018-05-05JonathanBorkum
Jonathan M. Borkum
1 Department of Psychology, University of Maine, Orono, ME, USA
2 Health Psych Maine, Waterville, ME, USA
Introduction
Since the discovery of neurogenesis in the adult mammalian brain, efforts have been made to use this capacity for purposes of neural regeneration. Approaches have included transplantation of exogenous or autologous stem cells, and the delivery of growth factors and other pharmaceuticals to stimulate innate neurogenesis (Carvalho et al., 2015).
However, there may already be an endogenous, integrated mechanism for increasing neural protection and repair that is physiologic and prevalent: the migraine attack. In this paper we will first review how the elements and feedback loops of a migraine attack may implement a neurorestorative function. We will then speculate on how migraine processes might be elicited to support exogenously administered stem cells in neural restoration.
Migraines and Oxidative Stress
Migraine attacks, prototypically lasting from 4 to 72 hours,involve moderate to severe throbbing pain that is increased by routine physical activity and accompanied by nausea and/or painful sensitivity to light and sound (Headache Classification Committee of the International Headache Society (IHS), 2013).In about 30% of cases, the migraine is preceded by visual aura,comprised of such experiences as scintillating lines and blind spots. During an attack, migraineurs feel and look ill.
In fact, these symptoms and the underlying physiology may defend the brain against oxidative stress (Borkum, 2018).Transient receptor potential ankyrin-1 (TRPA1) ion channels,found on pain-sensitive nerve endings in the dura mater, are able to detect oxidative stress and transduce it into a neural signal (Kozai et al., 2014). In turn, this signal elicits neurogenic inflammation, the cardinal feature of migraines, in animal models (Benemei et al., 2014).
In theory, the oxidative stress may have various sources,differing among migraineurs, including (1) mitochondrial defects (Welch et al., 1989; Markley, 2012) which tend to increase the production of superoxide (Stuart and Griffiths,2012) and impair antioxidant defenses (Wu et al., 2014); (2)cortical hyperexcitability (Coppola et al., 2007), in which oxidative stress may result from the high metabolic rate of brain tissue; (3) excessive activity of vasoconstrictors such as angiotensin, which entails production of superoxide as a byproduct(Ripa et al., 2014); and (4) genetically less active antioxidant enzymes (Neri et al., 2015). Moreover, migraine triggers may be exposures that further raise levels of oxidants in the brain(Borkum, 2016).
Thus, oxidative stress may be a final common pathway,signaling any number of unfavorable conditions in the brain.Further, oxidative stress can itself be harmful, as the brain is uniquely exposed and susceptible to damage from oxidants.Such damage may be a key early step in neurodegenerative diseases (Cahill-Smith and Li, 2014).
Migraine attacks appear to be a naturalistic means for the brain to counteract this damage, by reducing the brain’s energy demands, strengthening antioxidant defenses, delivering a range of growth factors, boosting neurogenesis, preventing apoptosis, facilitating mitochondrial biogenesis, and supporting the survival, proliferation, development, and complex architecture of neurons. We will first review these aspects of migraine in detail, and then consider possible applications.
Elements of the Migraine Attack
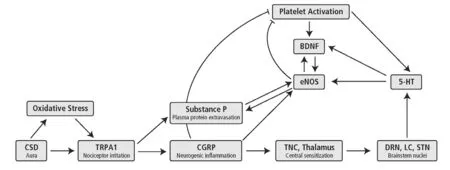
Figure 1 Schematic representation of a migraine attack.
The migraine attack unfolds as a complex, coordinated sequence of physiological processes (Goadsby, 2012), for which one starting point may be the stimulation of pain-sensitive nerve endings in the dura mater (Figure 1). This stimulation may arise from a buildup of oxidative stress, as we have seen. Alternatively, in migraine with visual aura, a wave of activation is thought to propagate over the cortex,followed by a marked decrease in spontaneous firing (cortical spreading depression; CSD), corresponding to the aura’s scintillations and scotomas, respectively. The irritation of nociceptive nerve endings may then arise from the potassium ions and glutamate released by this activity, in addition to the oxidants it generates.
Irritation of nociceptive fibers causes them to release calcitonin gene-related peptide (CGRP) and substance P from their distal ends, instigating neurogenic inflammation.CGRP causes mast cell degranulation, vasodilation, hyperemia and possibly further sensitization of nearby nociceptors. Substance P causes plasma protein extravasation into cerebrospinal fluid.
The activity of the pain fibers is transmitted proximally to the trigeminal nucleus caudalis and the thalamus, sensitizing central nociceptive pathways and contributing to the pain.The activation further spreads to such brainstem nuclei as the solitary tract nucleus (responsible for nausea and vomiting), the locus ceruleus and the hypothalamus (involved in the stress response), the basal ganglia (involved in pain reception), and likely the dorsal raphe nucleus (DRN). The simultaneous firing of these regions is characteristic of a migraine attack.
Meanwhile, platelet activating factor (PAF) is released early in a migraine, causing platelets to aggregate in the cerebral venules. Platelets and presumably the DRN release serotonin which,via5-HT2Breceptors in the blood vessels,activates endothelial nitric oxide synthase (eNOS) and raises production of nitric oxide. The platelets, in response to activation, and the endothelium, in response to nitric oxide,then release brain-derived neurotrophic factor (BDNF).
This stereotyped sequence of responses is further coordinated by feedback loops. Some are reinforcing and may sustain the migraine attack. Thus, serotonin, CGRP, substance P,PAF, and BDNF all raise the activity of eNOS. The resulting nitric oxide then facilitates the production of BDNF, and vice versa, in a positive feedback cycle (Monnier et al., 2017).Substance P and eNOS seem to be embedded in a similar cycle (Yonehara and Yoshimura, 1999, 2000).
There are also negative feedback loops that seem designed to prevent the neurogenic inflammation from triggering excessive classical inflammation. Thus, CGRP (Matsumoto et al., 1996) and nitric oxide (Golebiewska and Poole, 2015)limit platelet activation. Moreover, at least in the mouse peritoneal cavity, CGRP attenuates the movement of neutrophils and monocytes through the endothelium (Gomes et al., 2005) and thus CGRP may help to maintain the immune privilege of the CNS. Serotonin, directly through 5-HT2Creceptors (Hwang et al., 2008) and indirectly through the release of fractalkine (Cardona et al., 2006), limits the extent of microglial activation.
Migraines as a Healing Environment
Let us look more closely at these processes from the standpoint of neurorestoration.
Platelet activation
Platelets are classically thought to contribute to migraines by releasing serotonin, which may sensitize nociceptive nerve endings; proinflammatory cytokines, which may contribute to inflammation; and nitric oxide, adding to vasodilation(Danese, et al., 2014).
However, activated platelets are also first responders to a site of injury, releasing a range of growth factors to support healing, including BDNF, platelet-derived growth factor(PDGF), basic fibroblast growth factor (bFGF), vascular endothelial growth factor (VEGF), nerve growth factor (NGF),and transforming growth factor-β1 (TGF-β1) (Mancuso and Santagostino, 2017).
Not surprisingly, activated plateletsin vitrofacilitate angiogenesis, induce the survival and proliferation of neural stem cells, and support their differentiation into neurons and glia (Hayon et al., 2012a). In anin vivomodel of cerebral ischemia, activated platelets help limit damage and facilitate repair of brain tissue (Hayon et al., 2012b).
Now, excessive platelet activation could cause a harmful procoagulant state. Moreover, platelets, by producing interleukin-1β and certain chemokines, create an inflammatory milieu, attracting monocytes and encouraging their differentiation into macrophages (Mancuso and Santagostino,2017), potentially decreasing the viability of transplanted stem cells (Hermann et al., 2014). We have seen, however,that during a migraine attack, CGRP and nitric oxide serve to feed back and limit platelet activation.
Release of CGRP
CGRP is at the crux of migraines, causing mast cell degranulation, neurogenic inflammation, and pain sensitization in the periphery, and contributing to central sensitization in the trigeminal nucleus caudalis.
In addition, CGRP is protective, as it is a strong vasodilator, suppresses apoptosis of neurons under oxidizing(Schaeffer et al., 2003) and ischemic conditions (Abushik et al., 2017), reduces the expression of oxidant-generating NADPH oxidase (Zhou et al., 2010), and upregulates antiox-idant enzymes (She et al., 2003).
CGRP also increases the expression of a number of neurotrophic factors, including glial cell line-derived neurotrophic factor (GDNF), insulin-like growth factor-1 (IGF-1), NGF,VEGF, bFGF, and TGF-β (Russell et al., 2014; Ringer et al.,2017). Not surprisingly, then, CGRP increases neurogenesis in a mouse model of psychosocial stress (Hashikawa-Hobara et al., 2015).
In the brain, CGRP expression can be upregulated by injury and infection, and in turn can activate astrocytes and microglia (Reddington et al., 1995). This latter effect could work against the survival of transplanted stem cells. However, as noted, in migraine, serotonin, directly andviafractalkine, limits microglial activation.
CGRP has been used experimentally in stem cell grafting.In vitro, CGRP enhances survival and promotes the differentiation of adipose-derived stem cells into neurons (Yang et al., 2014). In anin vivomodel of spinal cord injury, CGRP had a chemotactic effect, attracting intrathecally injected human umbilical cord stem cells to the site of injury, at least in transection lesions (Zhang et al., 2016). Clinically, CGRP has been explored for preventing vasospasm following surgery for subarachnoid hemorrhage (European CGRP in Subarachnoid Haemorrhage Study Group, 1992).
BDNF
Production of BDNF by neurons and the endothelium is increased in migraine, and it released as well by activated platelets. It is thought to promote central pain sensitization in migraine attacks.
In addition, through uncoupling protein 2, BDNF decreases the production of oxidants by the mitochondria, and BDNF upregulates a number of antioxidant enzymes and proteins(Wu et al., 2016). In addition, of course, BDNF is a growth factor, and seems to underlie endogenous neuroprotection(Larsson et al., 1999), angiogenesis (Bowling et al., 2016) and the increased neurogenesis from exercise and antidepressant treatment (Vilar and Mira, 2016). BDNF has been used in mouse models of traumatic brain injury (TBI) (Kim et al.,2016) and spinal cord injury (Robinson and Lu, 2017) to support the survival of neural stem cells, their differentiation into neurons, and possible integration into neural circuits.
Serotonin
Substance P
Substance P mediates the extravasation of plasma proteins,chiefly albumin, into cerebrospinal fluid. Albumin is protective as it is an antioxidant (Taverna et al., 2013), carries unsaturated fatty acids that may aid in the repair of neuronal membranes (Rodriguez de Turco et al., 2002), and stimulates astrocytes to produce oleic acid, which facilitates the differentiation of neurons (Bento-Abreu et al., 2008).
In mice, IV injection of substance P markedly increases neural stem cell proliferation and differentiation into neurons, and leads to anatomical recovery after spinal cord injury. Substance P also promotes functional recovery, suggesting successful integration of the newly formed cells into neural networks (Yang et al., 2017). In ischemia, protein leakage through the blood-brain barrier seems to induce neurogenesis in latent niches throughout the midline circumventricular system (Lin et al., 2015).
Activation of eNOS
Nitric oxide, produced by eNOS that has been activated by serotonin, contributes to the vasodilation of migraines and to the release of substance P. Nitric oxide also protects the brain under ischemic conditions through a number of mechanisms including vasodilation, antagonism of oxidant production by the renin-angiotensin system (González et al., 2014), induction of antioxidant enzymes (Astort et al., 2014), and stimulation of mitochondrial biogenesis (Nisoli et al., 2005). Moreover,activated eNOS causes the endothelium to release VEGF and BDNF (Zhang et al., 2003; Monnier et al., 2017).
The endothelium is an important component of stem cells’neurovascular niche. Thus, eNOS can induce the proliferation,differentiation, and migration of neural progenitor cells (Chen et al., 2005) and spur angiogenesis (Asada et al., 2009). A nitric oxide donor increased neurogenesis and functional recovery in a rat model of ischemic stroke (Zhang et al., 2001).
Serotonin is released by platelets at the start of a migraine(Ferrari et al., 1989). Moreover, there is increased firing in the region of the dorsal raphe nucleus (Weiller et al., 1995),which may distribute serotonin throughout the cortex(Azmitia, 2007). These seem to reverse a serotonin deficit that intensifies in the days leading up to a migraine (Hamel,2007). Serotonin is thought to participate in an attack by promoting inflammation and sensitizing pain receptors.
In addition, serotonin has antioxidant propertiesin vitro(Kalogiannis et al., 2016).In vivoit causes astrocytes to release metallothioneins, a class of antioxidant enzyme, and cysteine, which is taken up by neurons and used to produce the antioxidant glutathione (Miyazaki and Asanuma, 2016).Serotonin is also a growth factor and induces the release of such other trophic factors as BDNF, VEGF (Greene et al.,2009), IGF-1 (Aberg et al., 2003), and S100B (Miyazaki and Asanuma, 2016). Through BDNF, serotonin increases the proliferation and migration of neural stem cells in the subventricular zone (Chiaramello et al., 2007).
Note that if release of serotonin were the only component of migraine, it could work against tissue healing by promoting vasoconstriction. However, this property of serotonin is antagonized in migraine by nitric oxide and CGRP.
Serotonin agonists have been used in animal models of TBI (Cheng, et al., 2016) and a clinical trial following ischemic stroke (Chollet, et al., 2011) to promote recovery.
Cortical spreading depression (CSD)
CSD is thought to underlie migraine aura (Lauritzen, 1994).In CSD, a wave of activation and reactive hyperemia spreads geographically over the cortex, leaving in its wake a region of diminished cortical firing. These are believed to correspond,respectively, to the scintillations and scotomas of aura.
Acutely, CSD raises oxidant production (Shatillo et al.,2013). This is followed by a number of processes that protect against subsequent ischemia, including activation of AMP kinase and downregulation of energy-demanding pathways(Viggiano et al., 2014), increased transcription of antioxidant enzymes and proteins (Choudhuri et al., 2002), and induction of uncoupling protein 5, which reduces production of superoxide by the mitochondria (Viggiano et al., 2016).
Further, the intense neuronal activation and consequent increased blood flow in CSD markedly upregulates the production of BDNF and transcription of the gene for its receptor,TrkB (Urbach et al., 2006). In this sense, CSD resembles electrical stimulation protocols to improve functional recovery after stroke or brain injury (Henrich-Noack et al., 2017).
Thus, migraine attacks appear to be an integrated set of homeostatic processes that defend the brain against oxidative stress. Moreover, each component of migraine seems to facilitate the survival, proliferation, differentiation, and/or migration of stem cells. These effects are summarized in the Table 1.
Prospects for Harnessing Migraines
For both grafted and endogenous stem cells, efficacy depends on a facilitative environment in the injured tissue (Hermann et al., 2014). Moreover, it is unlikely that any single molecule will be sufficient for neural regeneration; rather, combination strategies are needed (Hermann and Chopp, 2012; Morales et al., 2016). As we have seen, migraine attacks seem to function in a multifaceted way to provide a healing milieu.
For successful neural repair, stem cells must proliferate,differentiate, migrate, mature, survive, and become integrated into existing neural circuits (Jones and Connor, 2017).These processes, in turn, are affected by chemicals secreted in the microenvironments of the neurogenic niches and the target injured brain region, particularly neurotrophic factors(Carvalho et al., 2015).
However, the administration of these growth factors is hampered by their short half-lives in the body, rapid degradation, and poor ability to cross the blood-brain barrier(Carvalho et al., 2015). Moreover, an oversupply of neurotrophins is associated with epilepsy, autism and bipolar disorder, suggesting that their concentration in the brain must be tightly regulated (Carvalho et al., 2015). Stem cells themselves carry the theoretical risk of creating benign (Amariglio et al., 2009) or malignant tumors (Hermann et al., 2014).
Technologies are being developed to circumvent these problems, such as encapsulating neurotrophins in nanoparticles structured to protect the drug, provide sustained release, and target specific brain regions through receptor binding (Angelova et al., 2013; Angelov and Angelova, 2017;Angelova and Angelov, 2017; Guerzoni et al., 2017). However, the body may have its own techniques for enhancing the availability of growth factors. In particular, a migraine attack, by creating a physiologic environment conducive to neural repair, might be a naturalistic way of upregulating neurotrophic signaling, and provide a fertile ground for the successful utilization of stem cells.
This raises the question – admittedly speculative – of purposely drawing on migraine physiology in stem cell treatments. At this point, little is known to guide such a project.Logically, however, three broad approaches could be taken:(1) eliciting a full migraine attack combined with pain suppression; (2) eliciting a subset of migraine components; and(3) eliciting a subthreshold analogue of migraines.
Full migraine
An obvious way of eliciting a migraine, of course, would be to stimulate TRPA1 ion channels chemically. A range of naturally occurring molecules including thymol, ethyl vanillin, allyl isothiocyanate, cinnamaldehyde, and hydrogen sulfide open TRPA1 ion channels (Wu et al., 2017a; Weinhold et al., 2017).At least in skeletal muscle, TRPA1 agonists seem to activate stem cells (satellite cells) and facilitate their migration and early differentiation into myoblasts (Osterloh et al., 2016).
However, the effect of such an intervention may depend on the specific disorder. Thus, in a transgenic mouse model of Alzheimer’s disease, TRPA1 ion channels are more numerous on cortical neurons and hippocampal astrocytes than in wild type mice. Moreover, stimulation of this channel led to increased production of proinflammatory cytokines,increased deposition of amyloid beta, and exacerbation of behavioral deficits (Lee et al., 2016), suggesting that at least in genetic Alzheimer’s disease there may be a specific vulnerability to TRPA1 agonists. TRPA1 also contributed to cell death in a retinal model of ischemia (Araújo et al., 2017)but protected cardiac myocytes during reperfusion (Lu et al.,2016). Thus, the safety and efficacy of TRPA1 agonists would need to be assessed carefully in a context-dependent manner.
Subsets of migraine components
Several techniques that we can now understand to be eliciting parts of a migraine attack have already been studied.One such approach draws on Raskin et al.’s (1987) observation that deep brain stimulation (DBS) of the DRN can elicit a migraine-like headache. The DRN is one of the brainstem nuclei whose coordinated firing characterizes a migraine attack. DBS of the raphe nuclei improved working memory in an animal model of TBI (Carballosa Gonzalez et al., 2013).
Of course, DBS carries the risks of an invasive procedure.Moreover, Raskin et al. (1987) describe individuals for whom the migraine-like headaches were continuous and intractable. However, recent techniques may allow more moderate and noninvasive stimulation. Thus, oscillating high frequency electric fields can be positioned to interfere at a frequency that will influence neural firing at the desired depth in the brain (Grossman et al., 2017).
There may be other ways of eliciting aspects of the migraine attack by physical means. Thus, transcranial focused ultrasound with microbubbles has been used to transiently open the blood-brain barrier for delivery of drugs and to promote neurogenesis (Scarcelli, et al., 2014). This leads to sterile inflammation that shares certain features with migraine, including extravasation of albumin and other plasma proteins, and upregulation of VEGF and BDNF (Kovacs et al., 2017). However, the sterile inflammation has also included activation of microglia, increased proinflammatory cytokines, and migration of macrophages to the affected area, suggesting classical inflammation, which can result from protein extravasation (Ralay Ranaivo and Wainwright,2010). This may reflect overly intense treatment parameters(McMahon and Hynynen, 2017), with research needed to ascertain whether with gentler stimulation the effects can be restricted to neurogenic inflammation.
A milder and more physiologic approach would be to simulate several features of a migraine attack simultaneously. The cholinesterase inhibitor donepezil improves spatial learning in mice by causing hippocampal astrocytes to release CGRP (Narimatsu et al., 2009) and presumably downstream growth factors. We have seen that serotonin agonists,BDNF, CGRP, substance P, and a nitric oxide donor havebeen studied individually for promoting stem cell grafting and/or functional recovery. Considering migraine as an integrated system, conjoint activation of these processes might be more effective (Table 1).

Table 1 Migraine components
Subthreshold analogue of migraine
An alternative to activating parts of a migraine attack would be to elicit a subsyndromal version of an attack. Whether this is feasible is not yet known. Interestingly, however, there is evidence that stimulation of theZusanli(ST36) acupuncture point, just below the anterolateral knee, increases serotonin content of the DRN (Wu et al., 2017b), raises serum levels of BDNF (Tao et al., 2014), stimulates hippocampal neurogenesis after ischemia (Tao et al., 2014), suppresses apoptosis (Chavez et al., 2017), and may raise levels of activated eNOS (Leung et al., 2016), CGRP (Lee et al., 2012) and brain antioxidant defenses (Chavez et al., 2017). Thus, stimulation of this acupuncture point appears to replicate many of the neuroprotective features of migraine without the pain of a full attack.
Pain suppression
Of course were a full migraine attack to be elicited, it would need to be dissociated from pain in order to be of clinical value. Presumably this can be facilitated with analgesic medications that do not limit neurogenic inflammation.Alternatively, it might theoretically be possible to create a“pain-free migraine” by simultaneous use of TRESK potassium channel agonists. The TRESK channel, expressed primarily in neurons of the trigeminal, dorsal root, and autonomic ganglia, appears to decrease neuronal excitability in response to exposure to histamine, and thus may reduce pain under inflammatory conditions (Lafrenière and Rouleau, 2011). A very small number of individuals, all of them having migraine with typical visual aura, have been found to have a frameshift mutation in which TRESK channels are non-functional (Lafrenière and Rouleau, 2011). In theory,then, a TRESK agonist might make it possible to bring about a migraine attack without inducing pain.
At this point, it is unknown whether such a “migraine” is possible or would have neurorestorative properties. However, we have seen that individual migraine components indeed assist in neural repair, suggesting that pain is not an obligatory feature for migraines to be of use.
Conclusions
Thus, the idea that migraine attacks are an endogenous, integrated mechanism for neural repair, suggests their possible use in creating a healing environment for application in stem cell technology. Nonetheless, this possibility is still in its infancy. Research to find effective chemical or mechanical techniques for eliciting components of a migraine, dissociating these components from pain, and studies to determine safety and dosing in various pathophysiological contexts will be key. Stimulation of theZusanli(ST36) acupuncture point seems a particularly promising candidate.
Author contributions:JMB is responsible for all aspects of this paper.
Conflicts of interest:The authors declare that there is no conflict of interest.
Financial support:None.
Plagiarism check:Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewers:Joanna Czarnecka, Nicholaus Copernicus University,Poland; Angelina Angelova, Centre National de la Recherche Scientifique,France.
Aberg MA, Aberg ND, Palmer TD, Alborn AM, Carlsson-Skwirut C, Bang P, Rosengren LE, Olsson T, Gage FH, Eriksson PS (2003) IGF-I has a direct proliferative effect in adult hippocampal progenitor cells. Mol Cell Neurosci 24:23-40.
Abushik PA, Bart G, Korhonen P, Leinonen H, Giniatullina R, Sibarov DA, Levonen AL, Malm T, Antonov SM, Giniatullin R (2017) Pro-nociceptive migraine mediator CGRP provides neuroprotection of sensory,cortical and cerebellar neurons via multi-kinase signaling. Cephalalgia 37:1373-1383.
Amariglio N, Hirshberg A, Scheithauer BW, Cohen Y, Loewenthal R, Trakhtenbrot L, Paz N, Koren-Michowitz M, Waldman D, Leider-Trejo L,Toren A, Constantini S, Rechavi G (2009) Donor-derived brain tumor following neural stem cell transplantation in an ataxia telangiectasia patient. PLoS Med 6:e1000029.
Angelov B, Angelova A (2017) Nanoscale clustering of the neurotrophin receptor TrkB revealed by super-resolution STED microscopy. Nanoscale 9:9797-9804.
Angelova A, Angelov B (2017) Dual and multi-drug delivery nanoparticles towards neuronal survival and synaptic repair. Neural Regen Res 12:886-889.
Angelova A, Angelov B, Drechsler M, Lesieur S (2013) Neurotrophin delivery using nanotechnology. Drug Discov Today 18:1263-1271.
Araújo DSM, Miya-Coreixas VS, Pandolfo P, Calaza KC (2017) Cannabinoid receptors and TRPA1 on neuroprotection in a model of retinal ischemia. Exp Eye Res 154:116-125.
Asada M, Ebihara S, Yamanda S, Niu K, Okazaki T, Sora I, Arai H (2009)Depletion of serotonin and selective inhibition of 2B receptor suppressed tumor angiogenesis by inhibiting endothelial nitric oxide synthase and extracellular signal-regulated kinase 1/2 phosphorylation. Neoplasia 11:408-417.
Astort F, Mercau M, Giordanino E, Degese MS, Caldareri L, Coso O,Cymeryng CB (2014) Nitric oxide sets off an antioxidant response in adrenal cells: involvement of sGC and Nrf2 in HO-1 induction. Nitric Oxide 37:1-10.
Azmitia EC (2007) Serotonin and brain: evolution, neuroplasticity, and homeostasis. Int Rev Neurobiol 77:31-56.
Benemei S, Fusi C, Trevisan G, Geppetti P (2014) The TRPA1 channel in migraine mechanism and treatment. Br J Pharmacol 171:2552-2567.
Bento-Abreu A, Velasco A, Polo-Hernandez E, Perez-Reyes PL, Tabernero A, Medina JM (2008) Megalin is a receptor for albumin in astrocytes and is required for the synthesis of the neurotrophic factor oleic acid. J Neurochem 106:1149-1159.
Borkum JM (2016) Migraine triggers and oxidative stress: a narrative review and synthesis. Headache 56:12-35.
Borkum JM (2018) The migraine attack as a homeostatic, neuroprotective response to brain oxidative stress: preliminary evidence for a theory.Headache 58:118-135.
Bowling H, Bhattacharya A, Klann E, Chao MV (2016) Deconstructing brain-derived neurotrophic factor actions in adult brain circuits to bridge an existing informational gap in neuro-cell biology. Neural Regen Res 11:363-367.
Cahill-Smith S, Li JM (2014) Oxidative stress, redox signalling and endothelial dysfunction in ageing-related neurodegenerative diseases: a role of NADPH oxidase 2. Br J Clin Pharmacol 78:441-453.
Carballosa Gonzalez MM, Blaya MO, Alonso OF, Bramlett HM, Hentall ID(2013) Midbrain raphe stimulation improves behavioral and anatomical recovery from fl uid-percussion brain injury. J Neurotrauma 30:119-130.
Cardona AE, Pioro EP, Sasse ME, Kostenko V, Cardona SM, Dijkstra IM,Huang D, Kidd G, Dombrowski S, Dutta R, Lee JC, Cook DN, Jung S,Lira SA, Littman DR, Ransohoff RM (2006) Control of microglial neurotoxicity by the fractalkine receptor. Nat Neurosci 9:917-924.
Carvalho IM, Coelho PB, Costa PC, Marques CS, Oliveira RS, Ferreira DC(2015) Current neurogenic and neuroprotective strategies to prevent and treat neurodegenerative and neuropsychiatric disorders. Neuromolecular Med 17:404-422.
Chavez LM, Huang SS, MacDonald I, Lin JG, Lee YC, Chen YH (2017)Mechanisms of acupuncture therapy in ischemic stroke rehabilitation: a literature review of basic studies. Int J Mol Sci 18:E2270.
Chen J, Zacharek A, Zhang C, Jiang H, Li Y, Roberts C, Lu M, Kapke A,Chopp M (2005) Endothelial nitric oxide synthase regulates brain-derived neurotrophic factor expression and neurogenesis after stroke in mice. J Neurosci 25:2366-2375.
Cheng JP, Leary JB, Sembhi A, Edwards CM, Bondi CO, Kline AE (2016)5-hydroxytryptamine1A (5-HT1A) receptor agonists: A decade of empirical evidence supports their use as an efficacious therapeutic strategy for brain trauma. Brain Res 1640:5-14.
Chiaramello S, Dalmasso G, Bezin L, Marcel D, Jourdan F, Peretto P, Fasolo A, De Marchis S (2007) BDNF/ TrkB interaction regulates migration of SVZ precursor cells via PI3-K and MAP-K signalling pathways. Eur J Neurosci 26:1780-1790.
Chollet F, Tardy J, Albucher JF, Thalamas C, Berard E, Lamy C, Bejot Y,Deltour S, Jaillard A, Niclot P, Guillon B, Moulin T, Marque P, Pariente J,Arnaud C, Loubinoux I (2011) Fluoxetine for motor recovery after acute ischaemic stroke (FLAME): a randomised placebo-controlled trial. Lancet Neurol 10:123-130.
Choudhuri R, Cui L, Yong C, Bowyer S, Klein RM, Welch KM, Berman NE(2002) Cortical spreading depression and gene regulation: relevance to migraine. Ann Neurol 51:499-506.
Coppola G, Pierelli F, Schoenen J (2007) Is the cerebral cortex hyperexcitable or hyperresponsive in migraine? Cephalalgia 27:1427-1439.
Danese E, Montagnana M, Lippi G (2014) Platelets and migraine. Thromb Res 134:17-22.
European CGRP in Subarachnoid Haemorrhage Study Group (1992) Effect of calcitonin-gene-related peptide in patients with delayed postoperative cerebral ischaemia after aneurysmal subarachnoid haemorrhage. Lancet 339:831-834.
Ferrari MD, Odink J, Tapparelli C, Van Kempen GM, Pennings EJ, Bruyn GW (1989) Serotonin metabolism in migraine. Neurology 39:1239-1242.
Goadsby PJ (2012) Pathophysiology of migraine. Ann Indian Acad Neurol 15:S15-22.
Golebiewska EM, Poole AW (2015) Platelet secretion: From haemostasis to wound healing and beyond. Blood Rev 29:153-162.
Gomes RN, Castro-Faria-Neto HC, Bozza PT, Soares MB, Shoemaker CB,David JR, Bozza MT (2005) Calcitonin gene-related peptide inhibits local acute inflammation and protects mice against lethal endotoxemia. Shock 24:590-594.
González J, Valls N, Brito R, Rodrigo R (2014) Essential hypertension and oxidative stress: New insights. World J Cardiol 6:353-366.
Greene J, Banasr M, Lee B, Warner-Schmidt J, Duman RS (2009) Vascular endothelial growth factor signaling is required for the behavioral actions of antidepressant treatment: pharmacological and cellular characterization. Neuropsychopharmacology 34:2459-2468.
Grossman N, Bono D, Dedic N, Kodandaramaiah SB, Rudenko A, Suk HJ,Cassara AM, Neufeld E, Kuster N, Tsai LH, Pascual-Leone A, Boyden ES (2017) Noninvasive deep brain stimulation via temporally interfering electric fields. Cell 169:1029-1041.e16.
Guerzoni LP, Nicolas V, Angelova A (2017) In vitro modulation of TrkB receptor signaling upon sequential delivery of curcumin-DHA loaded carriers towards promoting neuronal survival. Pharm Res 34:492-505.
Hamel E (2007) Serotonin and migraine: biology and clinical implications.Cephalalgia 27:1293-1300.
Hashikawa-Hobara N, Ogawa T, Sakamoto Y, Matsuo Y, Ogawa M, Zamami Y, Hashikawa N (2015) Calcitonin gene-related peptide pre-administration acts as a novel antidepressant in stressed mice. Sci Rep 5:12559.
Hayon Y, Dashevsky O, Shai E, Varon D, Leker RR (2012a) Platelet microparticles promote neural stem cell proliferation, survival and differentiation. J Mol Neurosci 47:659-665.
Hayon Y, Dashevsky O, Shai E, Brill A, Varon D, Leker RR (2012b) Platelet microparticles induce angiogenesis and neurogenesis after cerebral ischemia. Curr Neurovasc Res 9:185-192.
Headache Classification Committee of the International Headache Society(IHS) (2013) The International Classification of Headache Disorders, 3rd edition (beta version). Cephalalgia 33:629-808.
Henrich-Noack P, Sergeeva EG, Sabel BA (2017) Non-invasive electrical brain stimulation: from acute to late-stage treatment of central nervous system damage. Neural Regen Res 12:1590-1594.
Hermann DM, Chopp M (2012) Promoting brain remodelling and plasticity for stroke recovery: therapeutic promise and potential pitfalls of clinical translation. Lancet Neurol 11:369-380.
Hermann DM, Peruzzotti-Jametti L, Schlechter J, Bernstock JD, Doeppner TR, Pluchino S (2014) Neural precursor cells in the ischemic brain - integration, cellular crosstalk, and consequences for stroke recovery. Front Cell Neurosci 8:291.
Hwang J, Zheng LT, Ock J, Lee MG, Suk K (2008) Anti-inflammatory effects of m-chlorophenylpiperazine in brain glia cells. Int Immunopharmacol 8:1686-1694.
Jones KS, Connor B (2017) Endogenous Brain Repair: Overriding intrinsic lineage determinates through injury-induced micro-environmental signals. Neurogenesis (Austin, Tex) 4:1-5.
Kalogiannis M, Delikatny EJ, Jeitner TM (2016) Serotonin as a putative scavenger of hypohalous acid in the brain. Biochim Biophys Acta 1862:651-661.
Kim JY, Choi K, Shaker MR, Lee JH, Lee B, Lee E, Park JY, Lim MS, Park CH, Shin KS, Kim H, Geum D, Sun W (2016) Promotion of cortical neurogenesis from the neural stem cells in the adult mouse subcallosal zone.Stem Cells 34:888-901.
Kovacs ZI, Kim S, Jikaria N, Qureshi F, Milo B, Lewis BK, Bresler M, Burks SR, Frank JA (2017) Disrupting the blood-brain barrier by focused ultrasound induces sterile inflammation. Proc Natl Acad Sci U S A 114:E75-E84.
Kozai D, Ogawa N, Mori Y (2014) Redox regulation of transient receptor potential channels. Antioxid Redox Signal 21:971-986.
Lafrenière RG, Rouleau GA (2011) Migraine: Role of the TRESK two-pore potassium channel. Int J Biochem Cell Biol 43:1533-1536.
Larsson E, Nanobashvili A, Kokaia Z, Lindvall O (1999) Evidence for neuroprotective effects of endogenous brain-derived neurotrophic factor after global forebrain ischemia in rats. J Cereb Blood Flow Metab 19:1220-1228.
Lauritzen M (1994) Pathophysiology of the migraine aura. The spreading depression theory. Brain 117:199-210.
Lee CH, Kim DK, Yook TH, Sasaki M, Kitamura N (2012) Effectiveness of electroacupuncture at Zusanli (ST36) on the immunohistochemical density of enteroendocrine cells related to gastrointestinal function. J Acupunct Meridian Stud 5:63-71.
Lee KI, Lee HT, Lin HC, Tsay HJ, Tsai FC, Shyue SK, Lee TS (2016) Role of transient receptor potential ankyrin 1 channels in Alzheimer’s disease. J Neuroinflammation 13:92.
Leung SB, Zhang H, Lau CW, Lin ZX (2016) Attenuation of blood pressure in spontaneously hypertensive rats by acupuncture was associated with reduction oxidative stress and improvement from endothelial dysfunction. Chin Med 11:38.
Lin R, Cai J, Nathan C, Wei X, Schleidt S, Rosenwasser R, Iacovitti L (2015)Neurogenesis is enhanced by stroke in multiple new stem cell niches along the ventricular system at sites of high BBB permeability. Neurobiol Dis 74:229-239.
Lu Y, Piplani H, McAllister SL, Hurt CM, Gross ER (2016) Transient receptor potential ankyrin 1 activation within the cardiac myocyte limits ischemia-reperfusion injury in rodents. Anesthesiology 125:1171-1180.
Mancuso ME, Santagostino E (2017) Platelets: much more than bricks in a breached wall. Br J Haematol 178:209-219.
Markley HG (2012) CoEnzyme Q10 and riboflavin: the mitochondrial connection. Headache 52 Suppl 2:81-87.
Matsumoto Y, Ueda S, Matsushita S, Ozawa T, Yamaguchi H (1996) Calcitonin gene-related peptide inhibits human platelet aggregation. Jpn Circ J 60:797-804.
McMahon D, Hynynen K (2017) Acute inflammatory response following increased blood-brain barrier permeability induced by focused ultrasound is dependent on microbubble dose. Theranostics 7:3989-4000.
Miyazaki I, Asanuma M (2016) Serotonin 1A receptors on astrocytes as a potential target for the treatment of Parkinson’s disease. Curr Med Chem 23:686-700.
Monnier A, Prigent-Tessier A, Quirie A, Bertrand N, Savary S, Gondcaille C, Garnier P, Demougeot C, Marie C (2017) Brain-derived neurotrophic factor of the cerebral microvasculature: a forgotten and nitric oxide-dependent contributor of brain-derived neurotrophic factor in the brain.Acta Physiol (Oxf) 219:790-802.
Morales, II, Toscano-Tejeida D, Ibarra A (2016) Non pharmacological strategies to promote spinal cord regeneration: a view on some individual or combined approaches. Curr Pharm Des 22:720-727.
Narimatsu N, Harada N, Kurihara H, Nakagata N, Sobue K, Okajima K(2009) Donepezil improves cognitive function in mice by increasing the production of insulin-like growth factor-I in the hippocampus. J Pharmacol Exp Ther 330:2-12.
Neri M, Frustaci A, Milic M, Valdiglesias V, Fini M, Bonassi S, Barbanti P(2015) A meta-analysis of biomarkers related to oxidative stress and nitric oxide pathway in migraine. Cephalalgia 35:931-937.
Nisoli E, Tonello C, Cardile A, Cozzi V, Bracale R, Tedesco L, Falcone S,Valerio A, Cantoni O, Clementi E, Moncada S, Carruba MO (2005)Calorie restriction promotes mitochondrial biogenesis by inducing the expression of eNOS. Science 310:314-317.
Osterloh M, Bohm M, Kalbe B, Osterloh S, Hatt H (2016) Identification and functional characterization of TRPA1 in human myoblasts. Pflugers Arch 468:321-333.
Ralay Ranaivo H, Wainwright MS (2010) Albumin activates astrocytes and microglia through mitogen-activated protein kinase pathways. Brain Res 1313:222-231.
Raskin NH, Hosobuchi Y, Lamb S (1987) Headache may arise from perturbation of brain. Headache 27:416-420.
Reddington M, Priller J, Treichel J, Haas C, Kreutzberg GW (1995) Astrocytes and microglia as potential targets for calcitonin gene related peptide in the central nervous system. Can J Physiol Pharmacol 73:1047-1049.
Ringer C, Tune S, Bertoune MA, Schwarzbach H, Tsujikawa K, Weihe E,Schütz B (2017) Disruption of calcitonin gene-related peptide signaling accelerates muscle denervation and dampens cytotoxic neuroinflammation in SOD1 mutant mice. Cell Mol Life Sci 74:339-358.
Ripa P, Ornello R, Pistoia F, Carolei A, Sacco S (2014) The renin-angiotensin system: a possible contributor to migraine pathogenesis and prophylaxis. Expert Rev Neurother 14:1043-1055.
Robinson J, Lu P (2017) Optimization of trophic support for neural stem cell grafts in sites of spinal cord injury. Exp Neurol 291:87-97.
Rodriguez de Turco EB, Belayev L, Liu Y, Busto R, Parkins N, Bazan NG,Ginsberg MD (2002) Systemic fatty acid responses to transient focal cerebral ischemia: influence of neuroprotectant therapy with human albumin. J Neurochem 83:515-524.
Russell FA, King R, Smillie SJ, Kodji X, Brain SD (2014) Calcitonin gene-related peptide: physiology and pathophysiology. Physiol Rev 94:1099-1142.
Scarcelli T, Jordão JF, O’Reilly MA, Ellens N, Hynynen K, Aubert I (2014)Stimulation of hippocampal neurogenesis by transcranial focused ultrasound and microbubbles in adult mice. Brain Stimul 7:304-307.
Schaeffer C, Vandroux D, Thomassin L, Athias P, Rochette L, Connat JL(2003) Calcitonin gene-related peptide partly protects cultured smooth muscle cells from apoptosis induced by an oxidative stress via activation of ERK1/2 MAPK. Biochim Biophys Acta 1643:65-73.
Shatillo A, Koroleva K, Giniatullina R, Naumenko N, Slastnikova AA, Aliev RR, Bart G, Atalay M, Gu C, Khazipov R, Davletov B, Grohn O, Giniatullin R (2013) Cortical spreading depression induces oxidative stress in the trigeminal nociceptive system. Neuroscience 253:341-349.
She F, Sun W, Mao JM, Wang X (2003) Calcitonin gene-related peptide gene therapy suppresses reactive oxygen species in the pancreas and prevents mice from autoimmune diabetes. Sheng Li Xue Bao 55:625-632.
Stuart S, Griffiths LR (2012) A possible role for mitochondrial dysfunction in migraine. Mol Genet Genomics 287:837-844.
Tao J, Chen B, Gao Y, Yang S, Huang J, Jiang X, Wu Y, Peng J, Hong Z,Chen L (2014) Electroacupuncture enhances hippocampal NSCs proliferation in cerebral ischemia-reperfusion injured rats via activation of notch signaling pathway. Int J Neurosci 124:204-212.
Taverna M, Marie AL, Mira JP, Guidet B (2013) Specific antioxidant properties of human serum albumin. Ann Intensive Care 3:4.
Urbach A, Bruehl C, Witte OW (2006) Microarray-based long-term detection of genes differentially expressed after cortical spreading depression.Eur J Neurosci 24:841-856.
Viggiano E, Viggiano D, Viggiano A, De Luca B, Monda M (2014) Cortical spreading depression increases the phosphorylation of AMP-activated protein kinase in the cerebral cortex. Neurochem Res 39:2431-2439.
Viggiano E, Monda V, Messina A, Moscatelli F, Valenzano A, Tafuri D,Cibelli G, De Luca B, Messina G, Monda M (2016) Cortical spreading depression produces a neuroprotective effect activating mitochondrial uncoupling protein-5. Neuropsychiatr Dis Treat 12:1705-1710.
Vilar M, Mira H (2016) Regulation of Neurogenesis by neurotrophins during adulthood: expected and unexpected roles. Front Neurosci 10:26.Weiller C, May A, Limmroth V, Juptner M, Kaube H, Schayck RV, Coenen HH, Diener HC (1995) Brain stem activation in spontaneous human migraine attacks. Nat Med 1:658-660.
Weinhold P, Hennenberg M, Strittmatter F, Stief CG, Gratzke C, Hedlund P(2018) Transient receptor potential a1 (TRPA1) agonists inhibit contractions of the isolated human ureter. Neurourol Urodyn 37:600-608.
Welch KM, Levine SR, D’Andrea G, Schultz LR, Helpern JA (1989) Preliminary observations on brain energy metabolism in migraine studied by in vivo phosphorus 31 NMR spectroscopy. Neurology 39:538-541.
Wu CL, Chen SD, Yin JH, Hwang CS, Yang DI (2016) Nuclear factor-kappaB-dependent sestrin2 induction mediates the antioxidant effects of BDNF against mitochondrial inhibition in rat cortical neurons. Mol Neurobiol 53:4126-4142.
Wu SW, Fowler DK, Shaffer FJ, Lindberg JEM, Peters JH (2017a) Ethyl vanillin activates TRPA1. J Pharmacol Exp Ther 362:368-377.
Wu YT, Wu SB, Wei YH (2014) Roles of sirtuins in the regulation of antioxidant defense and bioenergetic function of mitochondria under oxidative stress. Free Radic Res 48:1070-1084.
Wu YY, Jiang YL, He XF, Zhao XY, Shao XM, Sun J, Shen Z, Shou SY, Wei JJ, Ye JY, Yan SS, Fang JQ (2017b) 5-HT in the dorsal raphe nucleus is involved in the effects of 100-Hz electro-acupuncture on the pain-depression dyad in rats. Exp Ther Med 14:107-114.
Yang L, Li G, Ye J, Lu D, Chen Z, Xiang AP, Jiang MH (2017) Substance P enhances endogenous neurogenesis to improve functional recovery after spinal cord injury. Int J Biochem Cell Biol 89:110-119.
Yang Q, Du X, Fang Z, Xiong W, Li G, Liao H, Xiao J, Wang G, Li F (2014)Effect of calcitonin gene-related peptide on the neurogenesis of rat adipose-derived stem cells in vitro. PLoS One 9:e86334.
Yonehara N, Yoshimura M (1999) Effect of nitric oxide on substance P release from the peripheral endings of primary afferent neurons. Neurosci Lett 271:199-201.
Yonehara N, Yoshimura M (2000) Interaction between nitric oxide and substance P on heat-induced inflammation in rat paw. Neurosci Res 36:35-43.
Zhang R, Zhang L, Zhang Z, Wang Y, Lu M, Lapointe M, Chopp M (2001)A nitric oxide donor induces neurogenesis and reduces functional deficits after stroke in rats. Ann Neurol 50:602-611.
Zhang R, Wang L, Zhang L, Chen J, Zhu Z, Zhang Z, Chopp M (2003) Nitric oxide enhances angiogenesis via the synthesis of vascular endothelial growth factor and cGMP after stroke in the rat. Circ Res 92:308-313.
Zhang Y, Yang J, Zhang P, Liu T, Xu J, Fan Z, Shen Y, Li W, Zhang H (2016)Calcitonin gene-related peptide is a key factor in the homing of transplanted human MSCs to sites of spinal cord injury. Sci Rep 6:27724.
Zhou Z, Hu CP, Wang CJ, Li TT, Peng J, Li YJ (2010) Calcitonin gene-related peptide inhibits angiotensin II-induced endothelial progenitor cells senescence through up-regulation of klotho expression. Atherosclerosis 213:92-101.
杂志排行

中国神经再生研究(英文版)的其它文章
- Acupuncture and neuroregeneration in ischemic stroke
- The adjustment of γ-aminobutyric acidA tonic subunits in Huntington’s disease: from transcription to translation to synaptic levels into the neostriatum
- Bridging the gap: axonal fusion drives rapid functional recovery of the nervous system
- Collagen for brain repair: therapeutic perspectives
- Stimulating effect of thyroid hormones in peripheral nerve regeneration: research history and future direction toward clinical therapy
- Synaptic dysfunction in Alzheimer’s disease: the effects of amyloid beta on synaptic vesicle dynamics as a novel target for therapeutic intervention