上海动物园卷羽鹈鹕小种群遗传多样性的初步分析
2018-04-17黄康宁陈晓兰傅建义徐正强
周 颖, 马 珺, 黄康宁, 陈晓兰, 傅建义, 徐正强
(上海动物园, 上海 200335 )
由于种群数量的大幅减少,卷羽鹈鹕(Pelecanuscrispus) 已被世界自然保护联盟(IUCN)列为渐危种(Vulnerable)和中国的二级重要保护动物。20世纪50年代末,上海动物园从内蒙古乌粮苏湖自然保护区引进了十几只鹈鹕,以卷羽鹈鹕为主。20世纪70年代报道卷羽鹈鹕首次成功繁殖[1]。目前,种群已发展至40余羽(不包括出让至其他动物园的个体)。该种群是一个隔离种群,缺乏与外界个体的交流,再加上近年来人工繁殖个体很难引入种群内[2],种群出现了繁殖力下降,后代存活率降低等[3]问题。另一方面由于长期以来缺乏谱系标记和资料记录,种群内个体的遗传背景并不清楚,很难对其进行有效的种群管理。
随着对濒危动物保护遗传学研究的日益重视和加强,微卫星分子标记技术已被用来揭示濒危物种的进化历史、分析种群遗传结构、辅助种群调查、鉴定亲缘关系[4-5]和近缘物种及杂交个体[6]等。目前,尽管对卷羽鹈鹕的遗传学研究报道较少,但Machado等[7]对非洲南部白鹈鹕(P.onocrotalus)种群的微卫星进行筛选和分析,得到10个微卫星座位,并在褐鹈鹕(P.occidentalis)、美洲鹈鹕(P.erythrorhynchos)和粉红背鹈鹕(P.rufescens)中能够较好地扩增出相应的条带;Hickman等[8]筛选了美洲鹈鹕(P.erythrorhynchos)的9个微卫星座位,分析了北美洲东西两个种群的遗传多态性和种群结构,并用于制定种群保护计划。其中在现存8种鹈鹕的进化关系中,卷羽鹈鹕与粉红背鹈鹕亲缘关系很近,同为旧世界进化一支[9]。鉴于微卫星引物具有通用性的特点,本文从已报道的19个微卫星座位中筛选出12个微卫星多态性的座位用于分析上海动物园卷羽鹈鹕小种群的遗传多态性和种群结构,以期为该种群的有效管理提供遗传学依据,并为野生种群遗传多样性保护提供参考。
1 材料与方法
1.1 样品的采集
本次研究共收集了2010—2012年卷羽鹈鹕亚成体的血液样品,共计26份。用75%乙醇按4∶1处理血样,采样后快速置于-20℃冰箱保存。
1.2 基因组DNA的提取与扩增
用柱式小量组织/细胞基因组DNA提取试剂盒(华舜W6501)直接抽提。本实验从19个微卫星座位中筛选出12个多态性较丰富的座位进行分析:PeErH09-11、PeErH09-13、PeErH10-01、PeErH10-03、PeErH10-04、PeErH10-05[8]和PeOnH10-08、PeOnH10-09、PeOnH10-12、PeOnH10-13、PeOnH10-14和PeOnH10-15[7]。其引物序列详见表1。所有PCR扩增都在T3-Thermocycler DNA扩增仪上进行。反应总体积为20 μL:1×PCR Buffer (10 mmol/L Tris-HCl, 50 mmol/L KCl,pH 8.3),25.0 μg/mL BSA,1 μmol/L引物,200 μmol/L dNTPs,0.5 UTaq酶,约100 ng/μL DNA,其中MgCl2浓度参见表1。
所有扩增反应程序:94℃预变性10 min;94℃变性30 s,退火温度复性30 s,72℃延伸60 s,循环35次;循环结束后72℃再延伸10 min;扩增产物4℃保存。各引物的退火温度见表1。
扩增产物用8%变性聚丙烯酰胺凝胶电泳,电泳结果通过银染法检测[10]。等位基因的分子量大小通过100 bp Marker (TaKaRa)用ImageTool (IT)3.0软件确定。
1.3 数据分析
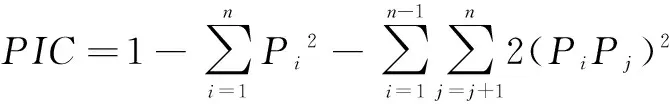
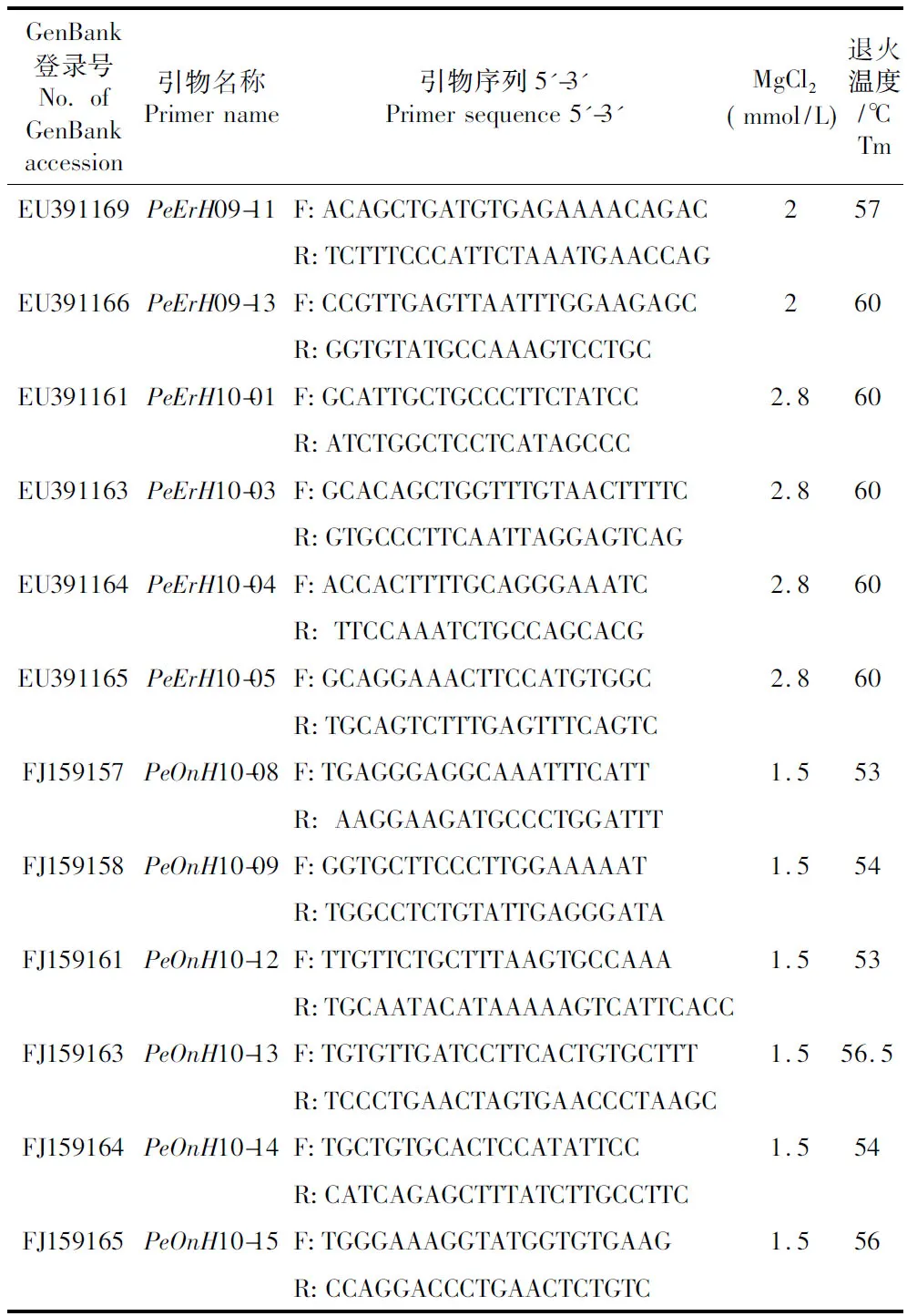
表1 12对微卫星座位的引物序列和PCR扩增条件
2 结果与分析
2.1 遗传多样性
在26个样品中成功扩增出了上述12个微卫星座位,多态性100%(表2)。每个位点的等位基因数不等,变幅为3~9个,平均5.33个;有效等位基因数平均为3.17个,占58.33%;观察杂合度的变化范围为0~1,平均为0.36;期望杂合度最大为0.58,最小为0.20,平均为0.39;Nei’s(1973)基因多样性指数H为0.32~0.8,平均为0.64;PIC为0.29~0.78,平均0.60(表2)。
2.2 聚类分析
利用NTsys 2.10e软件进行聚类分析,构建出个体间的亲缘关系树状图。结果(如图1)显示,在相似系数为0.77处,树形图分出11个聚类,其中16个个体构成一个大的聚类,编号2和4的个体聚成一类,其余8个个体后代分别聚类。
3 讨论与结论
3.1 遗传多样性分析
有效等位基因数,基因杂合度和多态信息含量是反映群体遗传变异的常用测度。在保护遗传学研究中,有时更强调等位基因数目对种群的影响,但有效基因数目容易受到样本量的影响[16]。与等位基因数相比,其杂合度的估计不依赖于样本的数量,因此可以较好地反应种群的遗传多样性水平[17]。基因杂合度是度量群体遗传变异的一个最适函数,Ott[18]将多态位点定义为杂合度至少为0.10的位点,而Takezaki和Nei[19]认为用于测定遗传差异的标记在群体中的平均杂合度在0.3~0.8才有实际意义。表2中各微卫星座位的杂合度均大于0.2,但有近一半的微卫星座位的基因杂合度小于0.3,平均值为0.39。以上说明该种群的基因杂合度值偏小,不具有较高的群体杂合度和丰富的遗传多样性,这与其实际的遗传背景是极其吻合的。上海动物园的卷羽鹈鹕种群作为一个封闭性的种群,由最初的建群个体发展到现在,种群内基因的纯合子增加,遗传差异明显减少。

表2 卷羽鹈鹕微卫星数据统计
NA:等位基因数 Number of alles;NE:有效等位基因数 Number of effective alleles;H:基因多样性指数 Genetic diversity index;HO:观察杂合度 Observed heterozygosity;HE:预期杂合度Expected heterozygosity;PIC:多态信息含量 Polymorphism information content
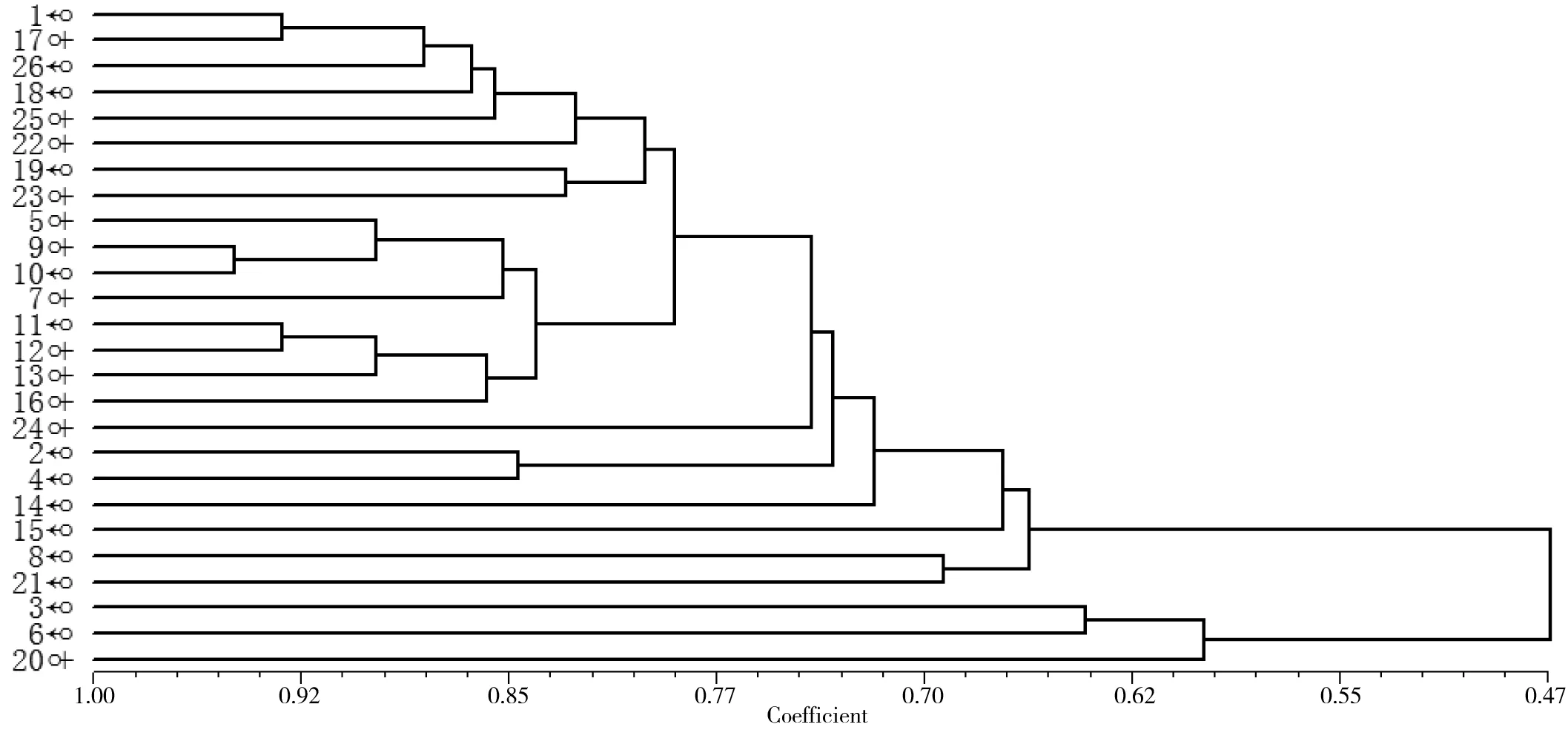
图 1 鹈鹕26个个体之间相互关系的分支聚类图
多态信息含量(PIC) 是等位基因频率和数目变化的函数,反映着基因变异程度的高低。当PIC>0.5 时,该座位为高度多态性座位;0.25 26个样本的聚类分析结果如图1所示,在相似系数为0.77处,树形图分出11个聚类。其中16个个体构成一个大的聚类,全为2对卷羽鹈鹕的后代,个体相似系数为0.79~0.95,这与观察结果相符合。除编号为2和4的个体亲缘系数较近(个体相似系数为0.84),可能来自同一对父母外,其余8个体分别聚类。这10羽后代个体可能在不同程度上混有白鹈鹕(P.onocrotalus)的基因。 卷羽鹈鹕种群(16个样本)的遗传多样性较低。这2对卷羽鹈鹕的后代如编号9和10,编号1和17,编号11和12的个体相似系数高达0.948、0.9310和0.9310。编号9和10的遗传距离最短,只有0.052,这表明该群体的遗传多样性很低。 上海动物园的卷羽鹈鹕存在近亲繁殖和杂交并存的现象。通过对卷羽鹈鹕遗传多样性分析,从遗传的角度掌握该种群的基本信息,便于对其进行合理有效的遗传管理。建议如下:1)根据聚类分析结果,重点在于建立2对纯卷羽鹈鹕种鸟后代谱系,对其后代个体添加脚标或芯片,进行长期监测;2)对于种群的近亲交配问题要予以高度重视,可采取人为干预措施,以避免过高的近交程度,而利于群体内杂合子的保留;3)可考虑引进亲缘关系较远的卷羽鹈鹕,以改善种群的遗传结构,增加种群的遗传多样性;4)继续监测和分析该卷羽鹈鹕种群的遗传多样性。鉴于聚丙烯酰胺凝胶电泳方法存在的不足,应采用更先进、有效的方法如测序,对种群内个体进行持续有效的遗传管理。 致谢:上海交通大学医学院动物实验科学中心提供了分子研究实验条件,特此致谢。 [1]何宝庆, 张 福, 方雪飞. 鹈鹕繁殖生态的观察[J]. 动物学杂志, 1984(4): 19-21. [2]马 珺, 傅建义, 虞坚颐, 等. 斑嘴鹈鹕引入天鹅湖的试验研究[J]. 生物学杂志, 2011, 28(1): 31-33. [3]徐正强, 沈莉萍, 桂剑锋, 等. 卷羽鹈鹕幼鸟感染大肠杆菌诊治报告[J]. 中国兽医杂志, 2014, 50(3): 34-35. [4]张于光, 李迪强, 饶立群, 等. 东北虎微卫星DNA遗传标记的筛选及在亲子鉴定中的应用[J]. 动物学报, 2003, 49(1): 118-123. [5]孙 强, 谈智华, 吴 锋, 等. 微卫星技术分析猎豹遗传资源及亲缘关系[J]. 江苏农业学报, 2010, 26(3): 541-545. [6]FITZSIMMONS N N, BUCHAN J C, LAM P V, et al. Identification of purebredCrocodylussiamensisfor reintroduction on Vietnam[J]. Journal of Experimental Zoology, 2002, 294(4): 373-381. [7]DE PONTE MACHADO M, FELDHEIM K A, SELLAS A B, et al. Development and characterization of microsatellite loci from the Great White Pelican (Pelecanusonocrotalus) and widespread application to other members of the Pelecanidae[J]. Conserv Genet, 2009, 10(4): 1033-1036. [8]HICKMAN C R, PETERS M B, CRAWFORD N G, et al. Development and characterization of microsatellite loci in the American white pelican (Pelecanuserythrorhynchos) [J]. Molecular Ecology Resources, 2008, 8(6): 1439-1441. [9]KENNEDY M, TAYLOR S A, NDVORNK P, et al. The phylogenetic relationships of the extant pelicans inferred from DNA sequence data[J]. Molecular Phylogenetics and Evolution, 2013, 66(1): 215-222. [10]高 东, 杜 飞, 朱有勇. 低背景、高分辨率PAGE简易银染法[J]. 遗传, 2009, 31(6): 668-673. [11]YEH F C, YANG R C, BOYLE T B J, et al. POPGENE, the user-friendly shareware for population genetic analysis[M]. Molecular Biology and Biotechnology Centre, University of Alberta, Canada, 1997. http://sites.ualberta.ca/~fyeh/faqs.html [12]KIMURA M,CROW J F. The number of alleles that can be maintained in a finite population[J]. Genetics, 1974, 49(4): 725-738. [13]NEI M. Analysis of gene diversity in subdivided population[J]. Proceedings of the National Academy of Science of the USA, 1973, 70(12): 3321-3323. [14]庾太林, 韩增超, 管清新, 等. 濒危黑颈长尾雉圈养种群RAPD遗传多样性分析[J]. 基因组学与应用生物学, 2012, 31(4): 369-373. [15]姚 慧, 钟金城, 姬秋梅, 等. 西藏牦牛遗传多样性的ISSR分析[J]. 生态学杂志, 2015, 34(11): 3278-3282. [16]乔利英, 袁亚男. 微卫星标记遗传多样性的度量指标及影响因素[J]. 中国畜牧兽医, 2010, 37(1): 107- 111. [17]蔡 垚. 应用微卫星标记进行江豚种群遗传结构和亲缘关系判别的研究[D].南京: 南京师范大学, 2004. [18]OTT J. Analysis of Human Genetic Linkage(Revised edition)[M]. Baltimore: Johns Hopkins University Press, 1991: 112-115. [19]TAKEZAKI N,NEI M. Genetic distance and reconstruction of polygenetic trees from microstatellite DNA[J]. Genetics, 1996, 144(1): 389-399. [20]BOTSTEIN D, WHITE R L, SKOLNICK M, et al. Construction of a genetic linkage map in man using restriction fragment polymorphisms[J]. The American Journal of Human Genetics, 1980, 32(3): 314-331.3.2 种群遗传管理
