春、夏季黄海南部和东海北部蟹类群落结构的空间分布特征
2018-04-11徐开达卢占晖薛利建周永东徐汉祥
孙 婉,徐开达,卢占晖,薛利建,周永东,徐汉祥
(1.浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,浙江省海洋渔业资源可持续利用技术研究重点实验室,农业部重点渔场渔业资源科学观测实验站,浙江舟山 316021;2.浙江海洋大学水产学院,浙江舟山 316022)
东海北部与黄海南部海域为陆架浅海区,受黄海沿岸流、台湾暖流、长江冲淡水、黄海冷水团以及东海沿岸流的混合作用,营养物质丰富,是包括蟹类在内的多种渔业生物种类的的产卵场与索饵场[1-2]。近年以来,对东黄海蟹类生态学方面的研究多集中于蟹类的种类组成和数量分布[3-10]、资源量评估[11-13]等方面[3-13],也有部分蟹类群落多样性与结构特征方面的研究[14-16],但对于研究海域蟹类群落结构的空间格局以及与环境因子之间的关系尚未见专门的报道。本研究基于“东海区主要渔场重要渔业资源调查与评估(2008-2009年)”项目所获得的桁杆拖虾网大面定点调查数据,采用典范对应分析(CCA)和多元统计分析等数据分析手段,对东海北部和黄海南部海域蟹类群落的空间分布特征以及环境因子之间的关系进行了分析与探讨,以期更为全面的了解研究海域的蟹类生态分布,保护其群落结构,修复海洋生态环境,为合理的利用蟹类资源提供基础资料与科学支撑。
1 材料与方法
1.1 数据来源
选取2008年春季(5月)和夏季(8月)开展的桁杆拖虾调查所获得的蟹类数据。2个航次的站位均设置为38个,其调查范围为 30°30′-33°00′N,127°00′E 以西海域,定点调查站位分布如图1所示。调查网具的囊网10支,桁杆长度为36 m,网囊网目54 mm。实际调查过程中,采用24 h连续不间断调查,每个站位拖网时间均为1 h,拖网调查的拖速控制在2 kn左右。所有采样流程与实验室测定过程均严格按照《海洋调查规范——海洋生物调查》[17]的相关标准开展。每个站位所捕获的渔获物均随机取样20 kg,如果不足20 kg,则将渔获物全部取样,带回实验室进行种类鉴定与称重。所取渔获样品中,包括蟹类在内的种类均尽可能鉴定到最小分类单元,并记录每种的尾数与重量。

图1 黄海南部和东海北部调查站位分布图(●)Fig.1 Location of sampling stations(●)in the southern Yellow Sea and the northern East China Sea
1.2 数据处理与分析
1.2.1 种类筛选
鉴于调查所捕获的某些蟹类种类,其生物量和出现频率均较低,对多元分析的结果影响较小。因此,对蟹类种类按照一定的筛选标准进行缩减[18]。本研究在构成“站位×种类生物量”矩阵的过程中,按照出现频率大于5%的蟹类作为分析对象进行筛选。
1.2.2 聚类分析和排序
为弱化各数据之间大小的悬殊,先对不同蟹类生物量进行平方根的转化。在此基础上,对各站位的Bray-Curtis相似性系数进行计算,此系数不但给予较丰富的种类更多的权重,而且不受矩阵为0的影响[19]。然后,采用非度量多维标度(NMDS)的排序方法以及未加权的组平均方法(UPGMA)的分类方法分析调查站位的蟹类组成特征[20]。基于以上2种分析方法的互补性,使之两种方法均为分析群落格局的有效工具,并可互为验证。
检验NMDS分析结果的优劣用胁强系数(Stress)来衡量,通常认为胁强系数(Stress)<0.05时,群落空间结构非常明显;stress<0.1时可认为群落空间结构较好;stress<0.2时表示可用NMDS的二维点图表示,其表征的群落空间具有一定的解析意义[21]。与此同时,采用非参数的相似性分析(ANOSIM)检验各分类和排序分组之间的差异显著性。用SIMPER方法计算各种类对样本组内相似性的平均贡献率。
1.2.3 群落与环境因子的关系
蟹类群落与环境因子之间的关系,采用典范对应分析(CCA,canonical correspondence analysis)的方法进行。为使种类和环境数据的分布更为接近正态分布,在数据分析之前,首先将其进行log(x+1)转换[22]。
采样站位图采用Surfer 8.0软件绘制。数据分析中,群落的聚类分析、非度量多维标度(NMDS)、非参数的相似性分析(ANOSIM)以及各种类在不同样本组内贡献率(SIMPER)的计算均使用PRIMER5.0软件进行处理;CCA分析在CANOCO4.0软件环境下进行运算,排序结果用物种-环境因子关系的双序图表示[23-24]。
2 结果
2.1 种类筛选
本次调查中共出现蟹类45种,隶属于12科,29属。其中,以梭子蟹科的种类最多,有9种,占全部蟹类种类总数的20.0%;其次为关公蟹科(8种),占17.8%。春季出现种类38种,夏季出现种类29种。其中,出现频率大于5%,生物量高于0.1%蟹类种类分别为36种(春季)和24种(夏季)(表1)。
2.2 蟹类群聚特征
根据各站位的蟹类组成进行NMDS排序和聚类分析,研究海域在春夏两季的蟹类群落均大致可分为3个群落(群落Ⅰ、群落Ⅱ和群落Ⅲ);胁强系数春季为0.10,夏季为0.14。这表明NMDS排序的结果可用二维点图表示;而且这两组群落具有较为明显的空间分布(图2)。
春季群落Ⅰ包括18个站位,群落Ⅱ包括10个站位,群落Ⅲ包括9个站位。相似性分析(ANOSIM)表明:群落Ⅰ和群落Ⅱ (R=0.908,P<0.01)、群落Ⅰ和群落Ⅲ(R=0.977,P<0.01)和群落Ⅱ和群落Ⅲ(R=0.752,P<0.01)之间均呈极显著差异,因此这样的群落划分是可行的。
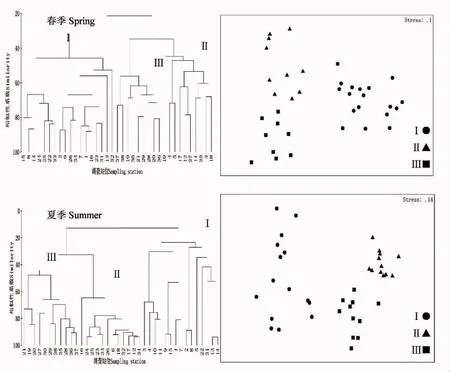
图2 春、夏季调查站位的聚类树状图(左)和MDS排序图(右)Fig.2 Charts of cluster analysis(left)and MDS ordination analysis(right)of the sampling stations in spring and summer
夏季群落Ⅰ包括15个站位,群落Ⅱ包括12个站位,群落Ⅲ包括11个站位。相似性分析(ANOSIM)表明:群落Ⅰ和群落Ⅱ(R=0.951,P<0.01)、群落Ⅰ和群落Ⅲ(R=0.688,P<0.01)和群落Ⅱ和群落Ⅲ(R=0.920,P<0.01)之间均呈极显著差异,因此这样的群落划分是合理且可行的。
根据春季和夏季蟹类群落类型的空间分布格局(图3),分别将这两个季节中蟹类的不同群落类型定义为近岸群落(群落Ⅰ)、混合群落(群落Ⅱ)和外海群落(群落Ⅲ)。其中,近岸群落(群落Ⅰ)为3个群落中最大的群落,主要分布于近岸向东延伸至125°00′E附近,南北方向则覆盖整个调查海域,春季近岸群落(群落Ⅰ)
有西北向东南延伸,而夏季则是由西南向东北延伸;外海群落(群落Ⅲ)在春夏两季的分布区域均相对较为稳定,主要分布在调查海域的外缘部分;混合群落(群落Ⅱ)则分布于近岸群落(群落Ⅰ)和外海群落(群落Ⅲ)之间狭长的海域,但夏季的分布范围略大于春季。

表1 黄海南部和东海北部海域春、夏季主要蟹类组成Tab.1 Main crab species component in the southern Yellow Sea and the northern East China Sea in spring and summer

图3 春、夏季黄海南部和东海北部蟹类群落格局的空间分布Fig.3 Charts showing spatial patterns of crab communities on the southern Yellow Sea and the northern East China Sea in spring and summer
2.3 各群落类型的主要特征种类
2.3.1 近岸群落(群落Ⅰ)
根据SIMPER分析的结果显示,近岸群落(群落Ⅰ)在春季的平均相似性为56.60%,夏季为32.65%;春季对近岸群落(群落Ⅰ)的相似性贡献较大的种类依次为:细点圆趾蟹Ovalipes punctatus、三疣梭子蟹Portunus trituberculatus和双斑蟳Charybdis bimaculata,夏季对近岸群落(群落Ⅰ)的相似性贡献较大的种类依次为:双斑蟳、日本蟳Charybdis japonica、三疣梭子蟹、泥脚隆背蟹Carcinoplax vestita、隆线强蟹Eucrate crenata和细点圆趾蟹(表2)。
2.3.2 混合群落(群落Ⅱ)
根据SIMPER分析的结果显示,混合群落(群落Ⅱ)在春季的平均相似性为52.36%,夏季为79.52%;春季对混合群落(群落Ⅱ)的相似性贡献较大的种类依次为:双斑蟳、细点圆趾蟹和泥脚隆背蟹,夏季对混合群落(群落Ⅱ)的相似性贡献较大的种类为:细点圆趾蟹(表2)。
2.3.3 外海群落(群落Ⅲ)
根据SIMPER分析的结果显示,外海群落(群落Ⅲ)在春季的平均相似性为62.53%,夏季为61.53%;春季对外海群落(群落Ⅲ)的相似性贡献较大的种类依次为长手隆背蟹Carcinoplax longimana、细点圆趾蟹、双斑蟳和中华隆背蟹Carcinoplax sinica,夏季对外海群落(群落Ⅲ)的相似性贡献较大的种类为:细点圆趾蟹、双斑蟳和长手隆背蟹(表2)。

表2 根据SIMPER分析得到的各蟹类群落类型中对平均相似性的贡献率>5%的主要特征种类Tab.2 The indicator species contributions>5%within communities from SIMPER analysis
2.4 群落与环境因子关系的CCA分析
春、夏季调查海域蟹类群落与表、底层温度(ST与BT)、表、底层盐度(SS与BS)以及水深(D)等5个环境因子的CCA排序结果如图4。根据分析结果:春夏两个季节前两个排序轴的特征值分别为0.497和0.194、0.420和0.230,而后两个排序轴的特征值均小于0.1,故使用前两个排序轴来分析春、夏季蟹类群落与环境因子之间的相关性。春、夏季前两个环境因子与种类排序轴之间的相关系数为0.939和0.740、0.847和0.752,前两个环境排序轴的相关系数均为0,前2个种类排序轴近似垂直(相关系数0.048 0和0.184 1))。以上CCA排序的特征值能够说明排序轴与环境因子间线性结合的程度较好地反映了调查站位与环境之间的关系,排序的结果是可靠的[23]。
春季,CCA第一排序轴与5个环境因子的相关性中,与水深(D)呈最大正相关,其相关系数为0.908 8,与底层温度(BT)呈较强负相关,相关系数为-0.566 3;第二排序轴与5个环境因子的相关性中,与底层温度(BT)呈最大负相关,其相关系数为-0.469 9(表3)。根据环境因子和排序轴的相关系数,春季影响调查海域蟹类群落的主要环境因子为水深(D),其次为底层温度(BT),而与其它3个环境因子的相关性较小(图4a)。
夏季,CCA第一排序轴与5个环境因子的相关性中,与水深(D)呈最大负相关,其相关系数为-0.691 1,其次为底层盐度(BS),相关系数为-0.646 6;第二排序轴与5个环境因子的相关性均不大,仅与底层温度(BT)呈一定负相关,相关系数为-0.379 6,与底层盐度(BS)和水深(D)的相关系数分别为-0.335 9和0.322 5,根据环境因子和排序轴的相关系数,夏季影响调查海域蟹类群落的主要环境因子为水深(D),其次为底层盐度(BS)和底层温度(BT)(图 4b)。

图4 春季(a)、夏季(b)调查站位与环境因子的CCA排序Fig.4 CCA ordination biplot between sampling stations and environmental factors in spring(a)and summer(b)

表3 环境因子与CCA排序轴之间的相关系数Tab.3 Correlation coefficients between CCA ordination axes and environmental factors
3 讨论
3.1 研究海域蟹类群落的划分
春、夏季调查海域蟹类群落的空间分布特征明显。根据桁杆拖虾网调查数据的等级聚类分析和多维度(NMDS)排序结果,研究海域蟹类群落可分为3种群落类型:近岸群落(群落Ⅰ)、混合群落(群落Ⅱ)和外海群落(群落Ⅲ)。据图3可得,3种群聚类型在春、夏季具有较为稳定的空间格局,随时间的变化相对较小。
近岸群落(群落Ⅰ)为3个群落中最大的群落,主要分布于近岸向东延伸至125°00′E附近海域,其水深在20~50 m之间,底层盐度变化范围在25~33.5之间,底层水温的变化范围在10~24℃之间,呈广温、广盐属性;外海群落(群落Ⅲ)在春、夏季的分布区域相对稳定,主要分布于调查海域的外缘部分,其水深在60~100 m之间,底层盐度稳定在34以上,底层水温的变化幅度为15~25℃,呈高温、高盐属性;混合群落(群落Ⅱ)则分布于以上2个群落之间狭长的海域,水深在30~60 m之间,底层盐度和底层温度的变化值不大,水文环境相对稳定。
3.2 各群落类型的主要特征种类
细点圆趾蟹、双斑蟳和三疣梭子蟹对春、夏季近岸群落(群落Ⅰ)内相似性贡献较大,为近岸群落(群落Ⅰ)的主要特征种类,日本蟳在仅在夏季对近岸群落(群落Ⅰ)内相似性贡献较大,为近岸群落(群落Ⅰ)的次要特征种类;细点圆趾蟹对春、夏季混合群落(群落Ⅱ)内相似性贡献最大,为混合群落(群落Ⅱ)的主要特征种类,而双斑蟳和泥脚隆背蟹仅在春季对混合群落(群落Ⅱ)内相似性贡献较大,为混合群落(群落Ⅱ)的次要特征种类;细点圆趾蟹、双斑蟳和长手隆背蟹对春、夏季外海群落(群落Ⅲ)内相似性贡献较大,为外海群落(群落Ⅲ)的主要特征种类,中华隆背蟹在春季对外海群落(群落Ⅲ)内相似性贡献较大,为外海群落(群落Ⅲ)的次要特征种类。
本研究中发现研究海域只在1种群落中出现的蟹类种类相对较少,多数蟹类种类同时出现在2个或3个群落类型中。其中,细点圆趾蟹对3个群落类型的群落内相似性贡献最大,为研究海域蟹类群落中的绝对优势种,也与以前的研究结果相一致[6,13-14,25]。除细点圆趾蟹以外,其它蟹类种类交错分布,但是其分布相对集中的区域明显不同,在不同群落类型中的生物量和出现频率有很大差异。影响研究海域蟹类群落划分的主要特征种类中,绝大分布种类为广温广盐种类,这些种类可以根据繁殖或摄食的需求而在调查海域范围内洄游以寻找最适的海洋环境,因此对3个群落类型的定义是相对的,很难定义一个永久的群聚地理区域或者与其它群聚相互隔离的群聚,这一点在东海其它游泳生物群落的划分中同样得以印证[26-29]。
3.3 与环境因子之间的关系
海洋生物群落在时空上呈现一定的异质性,这与复杂的理化因子和海底地貌关系十分密切[30]。本研究对调查海域的蟹类与环境因子进行典范对应(CCA)分析,结果表明:水深(D)是影响蟹类分布最为重要的环境因子,近岸群落(群落Ⅰ)主要分布于水深50 m以浅的海域,外海群落(群落Ⅲ)主要分布于水深60 m以深的海域,而混合群落(群落Ⅱ)中绝大部分站位水深在40~50 m之间,这也恰好说明混合群落(群落Ⅱ)为近岸群落(群落Ⅰ)与外海群落(群落Ⅲ)的过渡带。此外,影响调查海域蟹类群落的次要环境因子为底层温度(BT),而底层盐度(BS)、表层温度(ST)和表层盐度(SS)与蟹类的分布关系则不是十分明显。在相关东海蟹类分布与环境因子的关系研究中,大部分研究认为水深是影响蟹类分布的主要因素[4,7,14],也有学者认为温度和盐度是影响蟹类群落的主要环境因子[15]。其它一些环境因子(如水团特征、地形地貌、温跃层特征、底质类型、营养盐、溶解氧以及pH等)与水深的变化有着密切的关系,它们可以以不同的组合在某种程度上影响研究海域蟹类群落结构以及空间分布特征[16]。
研究海域空间尺度同样是影响蟹类群落类型因素之一,可能会导致对群落结构的划分不尽相同[31]。此外,本次调查使用的网具为桁杆拖虾网,根据蟹类的游泳能力、栖息水层等生态习性,本次调查的网具能够更好的捕获包括蟹类在内的底栖种类,从而使研究结果能够更加真实的反映研究海域蟹类群落的空间特征及其与环境因子之间的关系。
