铝处理对巨尾桉9号幼苗光合作用及叶绿素荧光特性的影响
2018-04-08广西农业职业技术学院生物技术系广西南宁530007广西大学林学院广西南宁530005
刘 云,杨 梅(.广西农业职业技术学院 生物技术系,广西 南宁 530007; 2.广西大学 林学院,广西 南宁 530005)
铝是地壳中含量最高的金属元素,常以难溶性的硅酸盐、氧化铝等形式存在于含铝矿物中,水溶性较小,但在酸性条件下(pH值<5),结合态铝可转化为可溶性铝,对植物产生毒害作用[1]。铝容易富集于植物根尖,抑制其根系生长,从而影响根对水和养分的吸收,限制整个植株的生长[2]。铝对植物根系的生长具有抑制作用,且随胁迫时间的增加而加剧[3-4],铝对植物的毒害作用还表现为影响阳离子的吸收[5]。光合作用是植物生长中重要的生理特征,叶绿素荧光分析技术常用于检测植物光合机构对环境胁迫的响应[6]。近年来,关于铝胁迫下光合作用及叶绿素荧光的研究不断增多[7-10],铝对植物的影响与其浓度有关[11],对西瓜[12]、山茶属植物[13]、刺槐[14]、烤烟[15]、野生油茶[16]等植物的光合作用及叶绿素荧光常表现为“低促高抑”;铝对植物的影响还与其处理时间密切相关。
桉树是我国南方主要的速生用材树种,我国南方桉树人工林面积大,广西是桉树的主要种植区,而且我国南方土壤多呈富铝化酸性土壤,酸雨、作物连作及施用肥料等会不断加剧土壤酸化程度,产生活性铝积累,这将会对广西大面积桉树人工林产生不利影响,因此,对铝胁迫下的桉树进行研究具有重要意义。相关研究指出,养分供应是影响桉树人工林生产力的关键因子,持续施肥可加剧土壤酸化,使桉树林地土壤活性铝不断增多[17-18],而土壤酸度对桉树林地土壤的铝含量及桉树根系铝的积累也存在着一定影响[19]。杨梅等[20-22]对铝胁迫下4个速生桉优良无性系(巨尾桉9号Eucalyptusgrandis×E.urophyllaNo.9、巨尾桉12号E.grandis×E.urophyllaNo.12、尾叶桉4号E.urophyllaNo.4、韦赤桉3号E.wetarensis×E.CamaldulensisNo.3)的生长及生理进行了初步研究,指出巨尾桉9号具有较强的耐铝性,但未涉及光合及叶绿素荧光特性等方面。因此,以耐铝性较强的桉树无性系巨尾桉9号为试验材料,从叶绿素含量、光合作用参数以及叶绿素荧光参数入手,探讨铝处理对光合作用和叶绿素荧光特性的影响,以期为筛选、培育和推广耐铝性桉树无性系及桉树耐铝机制方面的研究提供参考依据,同时也为其他林木耐铝性研究提供借鉴,为解决酸性土壤上人工林地力衰退问题奠定基础。
1 材料和方法
1.1 材料
供试所用的巨尾桉9号(E.grandis×E.urophyllaNo.9)幼苗由广西林业科学研究院种苗中心提供,为2个月生组培苗。
SHIMADZUUV-2550 型紫外可见分光光度计(岛津国际贸易有限公司)、CI 340型光合分析仪(美国CID公司)和Handy Fluor Cam荧光成像仪(法国PSI公司)。
1.2 方法
1.2.1供试幼苗培养试验于2015年在广西南宁市广西大学林学院实验室内进行。于3月10日将上述桉树幼苗进行预培养,采用Hoagland[23]营养液培养20 d(培养液浓度由1/5 Hoagland以1/5升幅逐级调至全Hoagland[23],pH值由5.5以0.1降幅逐级调至4.0),植株材料每周用质量分数为0.1%多菌灵消毒1次,试验处理前用质量分数为0.1%多菌灵消毒根系20 min,并转入pH值4.0、0.5 mmol/L CaCl2中清洗1 d,待进行试验处理。选取长势一致、根系发达、生长健康的巨尾桉9号幼苗移栽到嵌有小孔的圆形泡沫板(直径14 cm、高1 cm)上进行水培,每块泡沫板中间留一孔(直径1 cm)通气,泡沫板上距边缘2 cm处分布均匀地留4个孔(直径1 cm)用于固定幼苗,每孔栽植5株幼苗,每板20株。用岩棉将幼苗固定在泡沫板上,植株根系浸入装有营养液的容积2 L的烧杯(直径14.5 cm、高18.5 cm)中,烧杯周围用黑纸包裹以模拟土壤黑暗环境,每个烧杯放置1块泡沫板,栽培期间使用通气泵进行24 h连续通气,通气量为50 L/h。课题组前期研究表明,广西凭祥地区5年生桉树纯林林地不同土层土壤可溶性Al浓度平均为4.44 mmol/L[22],本试验采用完全随机设计,以0.5 mmol/L CaCl2为背景溶液,铝离子以AlCl3·6H2O形式加入其中,设置0、4.44 mmol/L 2个浓度,分别设置3.5、4.0两个pH值,每个试验分别处理1、6、12、24 h,以上各试验每个处理重复3次,每个重复20株幼苗。分别于处理的1、6、12、24 h测定各处理植株叶片的光合和叶绿素荧光参数,并随机取样,测定其叶绿素含量。
1.2.2叶绿素含量的测定叶绿素含量的测定采用熊庆娥[24]的方法进行,每个处理下随机摘取幼苗从上数第5片完全展开叶,将叶片洗净擦干后去除中脉,称取0.2 g,剪碎放入50 mL的具塞试管中,加入乙醇-丙酮混合液(等体积混合)20 mL,使叶片完全浸在液体中,盖上试管塞,放入暗处24 h,期间进行多次摇动,当叶片完全变成白色时进行比色,以提取液作对照,测其A665、A649,每个处理重复3次。相关计算公式如下:
叶绿素a质量浓度(mg/L):
Ca=13.95A665-6.88A649;
叶绿素b质量浓度(mg/L):
Cb=24.96A649-7.32A665;
叶绿素总质量浓度(mg/L):Ca+b=Ca+Cb;
叶绿素含量(mg/g)=
1.2.3光合及叶绿素荧光参数的测定每个处理分别随机选取5株测定光合参数及叶绿素荧光参数, 测定叶片选取不同方位从顶部向下数的第5~8片功能叶, 每个处理重复15片叶。将光照强度设置为500 μmol/(m2·s),CO2体积浓度设置为400 μL/L,测定净光合速率[μmol/(m2·s)]和蒸腾速率[mmol/(m2·s)];叶片经暗适应20 min 后测定初始荧光(Fo),接着,给1个饱和脉冲光[设置光量子通量密度为500 μmol/(m2·s)],1个脉冲后关闭,测定最大荧光(Fm),并参照Demming-Adams[25]的方法计算可变荧光(Fv)、PSⅡ潜在活性(Fv/Fo)。
1.3 数据处理和分析
采用SPSS 21.0统计软件对试验数据进行差异显著性分析。采用Excel数据处理软件对所得数据进行整理汇总,应用SPSS 21.0软件对数据进行单因素方差分析及多重比较(Duncan’s新复极差法)(P<0.05表示差异显著),包括同一处理不同时间间的差异显著性分析、同一时间不同处理间的差异显著性分析及处理、时间间的双因素方差分析。
2 结果与分析
2.1 不同铝处理及处理时间对巨尾桉9号叶绿素含量的影响
2.1.1铝处理、处理时间及其交互作用对巨尾桉9号叶绿素含量影响的双因素方差分析双因素方差分析结果表明(表1),不同铝处理、处理时间及其交互作用对叶绿素总含量、叶绿素a、叶绿素b含量及叶绿素a/b均具有极显著影响。
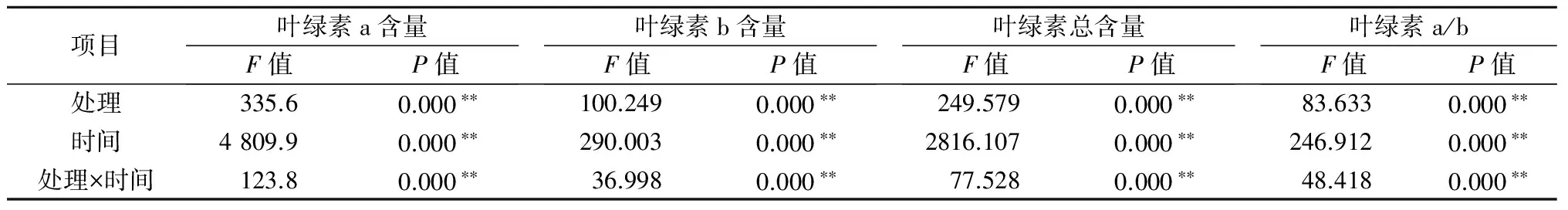
表1 铝处理、处理时间及其互作对叶绿素的影响
注:** 表示影响极显著(P<0.01),下同。
2.1.2铝处理、处理时间对巨尾桉9号叶绿素a含量的影响如表2所示,同一铝处理下,随着处理时间的增加,巨尾桉9号叶片叶绿素a含量变化较大,总体上呈先下降后上升趋势,且均在处理时间为12 h时达到最低。同一处理时间下,不同铝处理间叶绿素a含量相差较大,1、6、12、24 h 4个处理时间下的叶绿素a含量均在T1处理下达到最低,显著小于其他处理,且在Al 4.44 mmol/L、pH值3.5处理12 h时巨尾桉9号幼苗叶片叶绿素a含量最小,为0.44 mg/g。
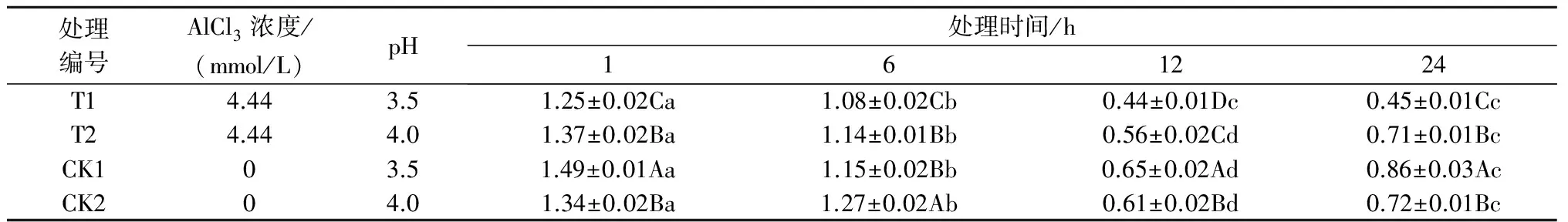
表2 铝处理、处理时间对叶绿素a含量的影响 mg/g
注:T表示处理,CK表示对照;数据为平均值±标准差,n=3;同列中不同的大写字母表示同一时间不同处理间差异显著(P<0.05);同行中不同的小写字母表示同一指标在不同处理时间差异显著(P<0.05)。下同。
2.1.3铝处理、处理时间对巨尾桉9号叶绿素b含量的影响由表3可知,同一铝处理下,不同处理时间之间巨尾桉9号叶片叶绿素b含量间差异明显,随着处理时间的增加,总体上呈先下降后上升趋势(除T1处理外),在处理时间为12 h时叶绿素b含量最低。同一处理时间下,不同铝处理间叶绿素b含量相差较大,1、6、12 h 3个处理时间下的叶绿素b含量均在T2处理下最低,而24 h处理下的叶绿素b含量在T1处理最低,且在Al 4.44 mmol/L、pH值3.5处理24 h时巨尾桉9号幼苗叶片叶绿素b含量达到最小,为0.23 mg/g。
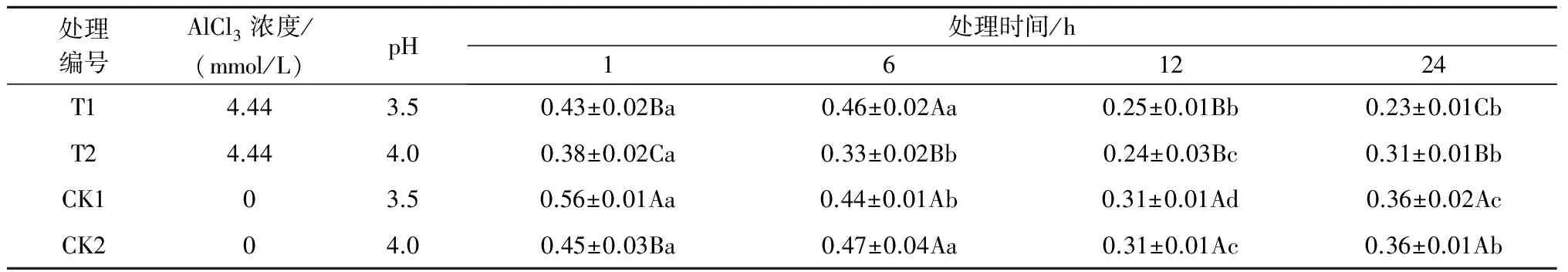
表3 铝处理、处理时间对叶绿素b含量的影响 mg/g
2.1.4铝处理、处理时间对巨尾桉9号叶绿素a/b的影响由表4可知,同一铝处理下,不同处理时间巨尾桉9号叶片叶绿素a/b间相差较大,随着处理时间的增加,叶绿素a/b先下降后上升(除T2处理外),处理时间为12 h时的叶绿素a/b最低。同一处理时间下,不同铝处理间叶绿素a/b也存在着一定的差异,处理时间为1 h 时T1、CK1的叶绿素a/b较低,而在6、12、24 h处理下的叶绿素a/b在T1处理最低,且在Al 4.44 mmol/L、pH值3.5处理12 h时巨尾桉9号幼苗叶片叶绿素a/b达到最小,为1.76。
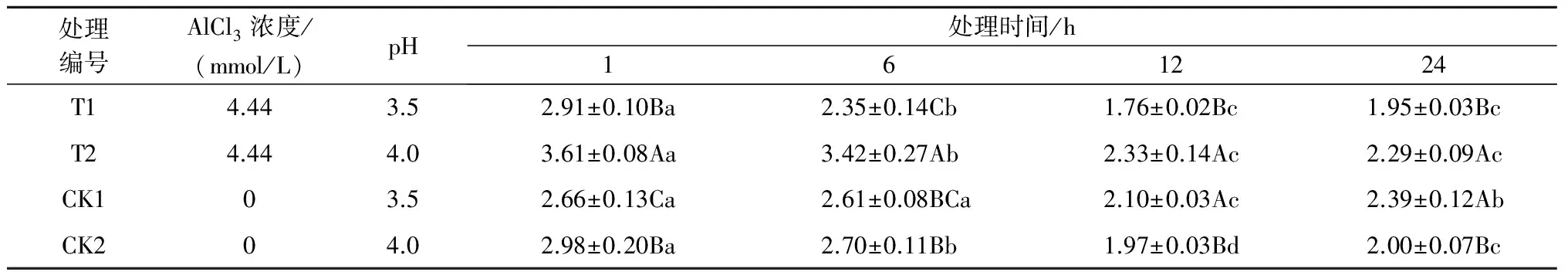
表4 铝处理、处理时间对叶绿素a/b的影响
2.1.5铝处理、处理时间对巨尾桉9号叶绿素总含量的影响表5所示,同一铝处理下,不同处理时间巨尾桉9号叶片叶绿素总含量间相差较大,随着处理时间的增加,叶绿素总含量总体上呈先下降后上升趋势(除T1处理外),且在处理时间为12 h时达到最低。同一处理时间下,不同铝处理间叶绿素总含量差异较大,在1、12、24 h时,T1处理下的叶绿素总含量最低,且在Al 4.44 mmol/L、pH值3.5处理24 h时巨尾桉9号幼苗叶片叶绿素总含量达到最小,为0.68 mg/g。
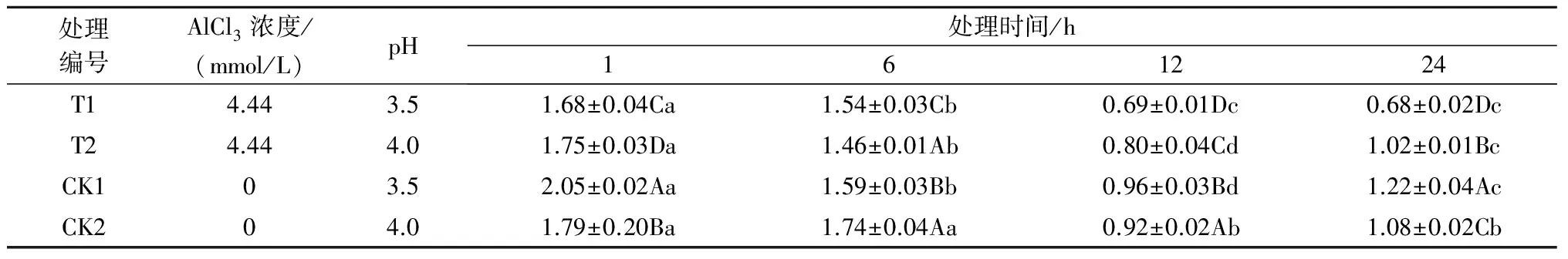
表5 铝处理、处理时间对叶绿素总含量的影响 mg/g
2.2 不同铝处理及处理时间对巨尾桉9号光合作用的影响
2.2.1铝处理、处理时间及其交互作用对巨尾桉9号光合作用影响的双因素方差分析双因素方差分析结果表明(表6),不同铝处理、处理时间及其交互作用对巨尾桉9号幼苗叶片净光合速率的影响不显著,而不同铝处理对蒸腾速率具有显著影响,不同处理时间、铝处理与处理时间的交互作用对蒸腾速率的影响均达到极显著水平。
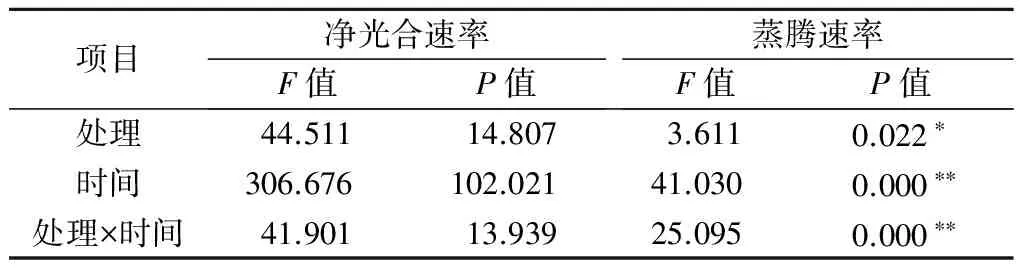
表6 铝处理、处理时间及其互作对光合作用的影响
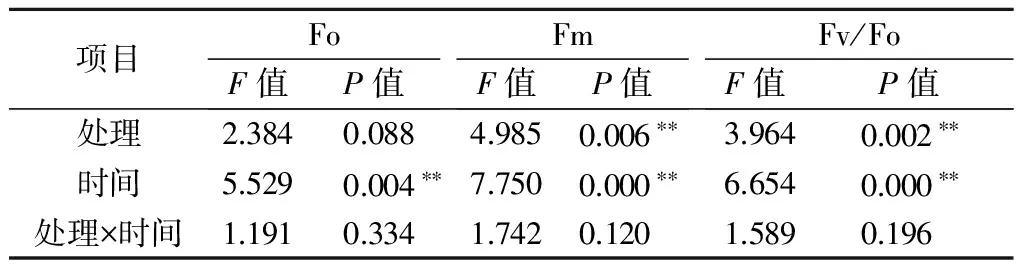
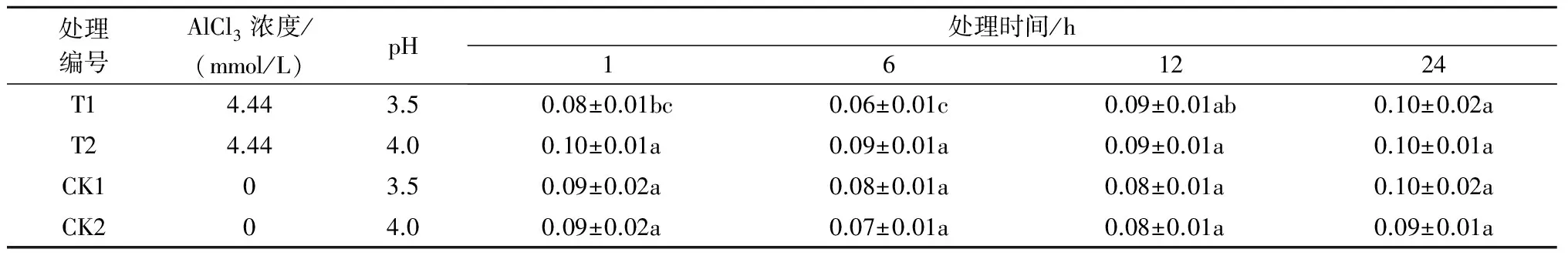
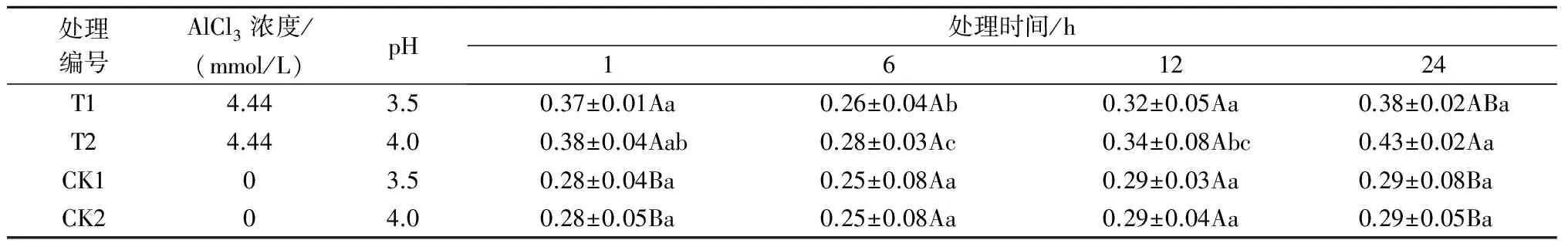
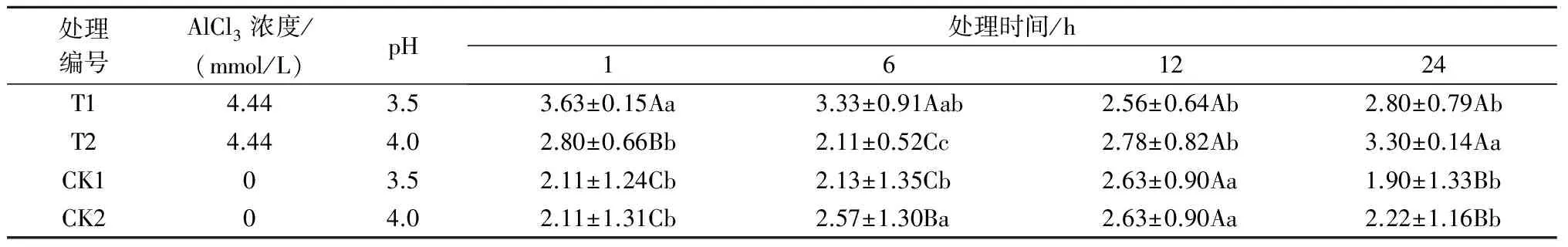
注:*表示影响显著(0.01 2.2.2铝处理、处理时间对巨尾桉9号净光合速率的影响如表7所示,同一铝处理下,不同处理时间巨尾桉9号叶片净光合速率间相差较大,随着处理时间的增加,巨尾桉9号叶片净光合速率总体上呈先上升后下降趋势,处理时间为6 h时,达到最高。同一处理时间下,不同铝处理间净光合速率存在一定差距,1、6、12、24 h 4个处理时间下的净光合速率均在T1处理下达到最低。 表7 铝处理、处理时间对净光合速率的影响 μmol/(m2·s) 2.2.3铝处理、处理时间对巨尾桉9号蒸腾速率的影响如表8所示,同一铝处理下,不同处理时间巨尾桉9号叶片蒸腾速率间差异较明显,随着处理时间的增加,巨尾桉9号叶片蒸腾速率总体上先上升后下降,在处理时间为6 h时,达到最高。同一处理时间下,不同铝处理间的蒸腾速率相差较大,1、12 h 处理时间下的蒸腾速率在T1处理下达到最低;而6、24 h 处理时间下的蒸腾速率在T2处理下达到最低。 表8 铝处理、处理时间对蒸腾速率的影响 mmol/(m2·s) 2.3.1铝处理、处理时间及其交互作用对巨尾桉9号叶绿素荧光参数影响的双因素方差分析双因素方差分析结果表明(表9),不同铝处理及其交互作用对巨尾桉9号幼苗叶片叶绿素荧光参数Fo的影响不显著,而不同处理时间对Fo具有极显著影响;不同铝处理、处理时间对Fm、Fv/Fo的影响均达到极显著水平,而不同铝处理、处理时间的交互作用对Fm、Fv/Fo的影响均不显著。 表9 铝处理、处理时间及其互作对叶绿素荧光参数的影响 2.3.2铝处理、处理时间对巨尾桉9号Fo的影响由表10可知,不同铝处理及处理时间下,巨尾桉9号叶片Fo的变化均较小,在铝处理下,巨尾桉9号叶片Fo随着时间的增加呈先减少后增加趋势,在6 h时达到最低。 表10 铝处理、处理时间对巨尾桉9号幼苗Fo的影响 2.3.3铝处理、处理时间对巨尾桉9号Fm的影响由表11可知,同一铝处理下,随着处理时间的增加,巨尾桉9号叶片Fm总体上先下降后上升,在处理时间为6 h时达到最低,而在24 h时达到最高。同一处理时间下,不同铝处理间的Fm相差较大,1、6、12、24 h 处理时间下的Fm均在T2处理下达到最高。 表11 铝处理、处理时间对巨尾桉9号幼苗Fm的影响 2.3.4铝处理、处理时间对巨尾桉9号Fv/Fo的影响由表12所示,在T1处理下,随着处理时间的增加,巨尾桉9号叶片Fv/Fo逐渐下降,在T2处理下,随着处理时间的增加,巨尾桉9号叶片Fv/Fo先下降后增加;在CK1、CK2处理下随着处理时间的增加,巨尾桉9号叶片Fv/Fo呈先增加后下降趋势。 表12 铝处理、处理时间对巨尾桉9号幼苗Fv/Fo的影响 叶绿体是植物光合作用的场所,也是对胁迫环境最敏感的细胞器[26]。叶绿素是植物体内的主要光合色素,在光合作用中具有关键性的作用,参与光能的吸收、传递和转化,其含量的变化可反映植物叶片光合作用的强弱[27]。叶绿素a和叶绿素b均是光合膜的组成部分,而比值可用来表示植物捕获、转化光能的能力[28]。叶绿素荧光常被用于评价环境胁迫对植物光合结构的影响,原初反应作为光合作用的起点,包括光能的吸收、传递和转换,是由叶绿体内片层膜上的蛋白质复合体和与它们相结合的色素分子共同来完成的一系列过程,研究认为,Fo与叶绿素a/b有关。本研究中,不同铝处理下,随着处理时间的增加,巨尾桉9号叶片叶绿素含量总体呈先降低后升高趋势,以Al 4.44 mmol/L、pH值3.5处理12 h巨尾桉9号幼苗叶片叶绿素a含量及叶绿素a/b均达到最小,而处理24 h时叶绿素总含量、叶绿素b含量、净光合速率均达到最小,铝处理、处理时间及其交互作用对叶绿素a、叶绿素b、叶绿素总含量及叶绿素a/b均有极显著影响,说明铝处理可以降低巨尾桉9号叶片的叶绿素总含量、叶绿素a、叶绿素b含量及叶绿素a/b,且随着铝处理时间的增加,巨尾桉9号叶片的叶绿体被膜被破坏,叶绿素的合成能力下降,导致叶绿素含量降低,从而对植株的光合作用造成影响;植物吸收的铝离子还可与合成叶绿素相关酶的肽链中富含巯基的部分结合,从而改变酶的结构[29];随着处理时间的增加,巨尾桉9号对铝处理的危害逐渐适应,其叶绿素含量又有所升高。 随着处理时间的增加,巨尾桉9号净光合速率与蒸腾速率在6 h时有所升高,而后下降,且铝处理、处理时间及其交互作用对蒸腾速率有极显著影响,较短时间的铝处理下巨尾桉9号光合作用还未受到抑制,而随着处理时间的增加,净光合速率与蒸腾速率逐渐降低,说明较短时间内,虽然叶绿素含量呈下降趋势,植物为维持基本生理活动而发生光合补偿效应[15],其捕获、转化光能的能力仍能维持其光合作用,且铝处理刺激了叶绿体,使其对铝处理的危害作出响应,提高光合作用以产生更多能量供植物在逆境下更好生长[14];而随着铝处理时间的增加,叶绿素含量不断下降,叶绿体内的光合Hill反应和光合磷酸化系统受到抑制,从而抑制了光合作用的光反应过程,净光合速率与蒸腾速率下降[13]。 叶绿素含量与荧光参数有一定的相关性[10],随着处理时间的增加,巨尾桉9号叶绿素荧光参数Fo、Fm及Fv/Fo变化趋势与其叶绿素含量相同,呈先降低后增高趋势,不同铝处理、铝处理时间对Fm、Fv/Fo的影响均达到极显著水平,铝处理降低了叶绿素含量,迫使PSⅡ反应中心和潜在活性中心受损,抑制光合作用的原初反应,光合电子传递过程受到影响,其叶片潜在光合活力下降[30]。 在1~24 h内,随着铝处理时间增长,总体上,巨尾桉9号幼苗叶绿素含量及其叶绿素荧光参数Fo、Fm及Fv/Fo逐渐降低,而后又有所升高,而光合作用则表现为先增强后减弱,在短时间(1~12 h)内巨尾桉9号幼苗受到一定伤害,到24 h时该伤害可以通过适应及植物自身抗逆境机制的调节而得到一定缓解,说明巨尾桉9号对酸铝处理具有一定的适应能力。 参考文献: [1]Kochian L V.Cellular mechanisms of aluminum toxicity and resistance in plants[J].Annual Review of Plant biology,1995,46(1):237-260. [2]Rangel A F,Rao I M,Horst W J.Intracellular distribution and binding state of aluminum in root apices of two common bean(Phaseolusvulgaris) genotypes in relation to Al toxicity[J].Physiologia Plantarum,2009,135(2):162-173. [3]王水良,王平,王趁义.铝处理下马尾松幼苗有机酸分泌和根际pH值的变化[J].生态与农村环境学报,2010,26(1):87-91. [4]Tahara K,Norisada M,Yamanoshita T,etal.Role of aluminum-binding ligands in aluminum resistance ofEucalyptuscamaldulensisandMelaleucacajuputi[J].Plant and Soil,2008,302(1/2):175-187. [5]Huang J H,Bachelard E P.Effects of aluminium on growth and cation uptake in seedlings ofEucalyptusmanniferaandPinusradiata[J].Plant and Soil,1993,149(1):121-127. [6]Kitao M,Utsugi H,Kuramoto S.Light dependen photosythesis characteristics indicated by chlorophyll fluorescence in five mangrove species native to pohnpei island,micronesia[J].Physiol Plant,2003,117(3):376-382. [7]高培培,章艺,吴玉环,等.外源水杨酸对铝胁迫下栝楼光合特性及耐铝性的影响[J].水土保持学报,2012,26(6):268-273. [8]刘强,尹丽,龙婉婉,等.铝胁迫对蓼科植物生长和光合、蒸腾特性的影响[J].广西植物,2011,31(2):227-232,284. [9]李俊钰,胥晓,杨鹏.铝胁迫对青杨雌雄幼苗生理生态特征的影响[J].应用生态学报,2012,23(1):45-50. [10]陈香,胡雪华,肖宜安.铝胁迫下北美车前和车前生长及叶绿素荧光特性的比较研究[J].植物研究,2011,31(6):680-685. [11]Chen L S,Tang N,Jiang H X,etal.Changes in organic acid metabolism differ between roots and leaves ofCitrusgrandisin response to phosphorus and aluminum interactions[J].Journal of Plant Physiology,2009,166(18):2023-2034. [12]郑阳霞,贾松涛,赵英鹏,等.铝胁迫对西瓜幼苗光合及叶绿素荧光特性的影响[J].华北农学报,2015,30(4):150-156. [13]顾艳红,刘鹏,徐根娣,等.铝胁迫对山茶属植物光合特性的影响[J].浙江林业科技,2006,26(4):15-18. [14]赵燕,李莎莎,柳晨意,等.铝胁迫对刺槐幼苗光合色素和矿质元素含量的影响[J].江苏农业科学,2015,43(4):200-202. [15]李淮源,刘柏林,邓世媛,等.铝胁迫对烤烟生长和光合特性的影响[J].烟草科技,2015(9):9-13,26. [16]贺根和,刘强,彭水娥.铝胁迫对野生油茶光合特性的影响[J].湖北农业科学,2010,49(7):1593-1595,1598. [17]Rhoades C,Binkley D.Factors influencing decline in soil pH in Hawaiian eucalyptus and albizia plantations[J].Forest Ecology and Management,1996,80:47-56. [18]Rifai S W,Markewitz D,Borders B.Twenty years of intensive fertilization and competing vegetation suppression in loblolly pine plantations:Impacts on soil C,N,and microbial biomass[J].Soil Biology & Biochemistry,2010,42(5):713-723. [19]Chen R F,Shen R F,Gu P,etal.Investigation of aluminum tolerant species in acid soils of south China[J].Communications in Soil Science and Plant Analysis,2008,39(9/10):1493-1506. [20]杨梅,黄寿先,方升佐,等.酸铝处理下4个速生桉优良无性系的生长反应[J].植物营养与肥料学报,2011,17(1):195-201. [21]杨梅,黄晓露,谭玲,等.4个桉树优良无性系的耐铝性评价指标分析[J].中南林业科技大学学报,2011,31(9):28-32. [22]Yang M,Tan L,Xu Y Y,etal.Effect of low pH and aluminum toxicity on the photosynthetic characteristics of different fast-growing 3 eucalyptus vegetatively propagated clones[J].PLoS ONE,2015,10(6):E0130963.DOI:10.1371/ journal.pone.0130963. [23]陈帅飞.桉树水培采穗圃营养液配比及营养不平衡对桉树苗生长影响研究[D].北京:中国林业科学研究院,2007. [24]熊庆娥.植物生理学实验教程[M].成都:四川科学技术出版社,2003. [25]Demmig-Adams B,Adams W W,Baker D H,etal.Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation[J].Physiologia Plantarum,1996,98(2):253-264. [26]王瑜,王思荣,张玲玲.铝胁迫对木荷幼苗光合特性的影响及添加盐基阳离子和磷的调节作用[J].热带亚热带植物学报,2014,22(1):61-67. [27]吕三三,杜国栋,刘志琨,等.覆草对苹果叶片显微结构及光系统功能的影响[J].中国农业科学,2015,48(1):130-139. [28]应小芳,刘鹏.铝胁迫对大豆叶片光合特性的影响[J].应用生态学报,2005,16(1):166-170. [29]曹林,吴玉环,章艺,等.外源水杨酸对铝胁迫下菊芋光合特性及耐铝性的影响[J].水土保持学报,2015,29(4):260-266. [30]俞慧娜,刘鹏,徐根娣.大豆生长及叶绿素荧光特性对铝胁迫的反应[J].中国油料作物学报,2007,29(3):257-265.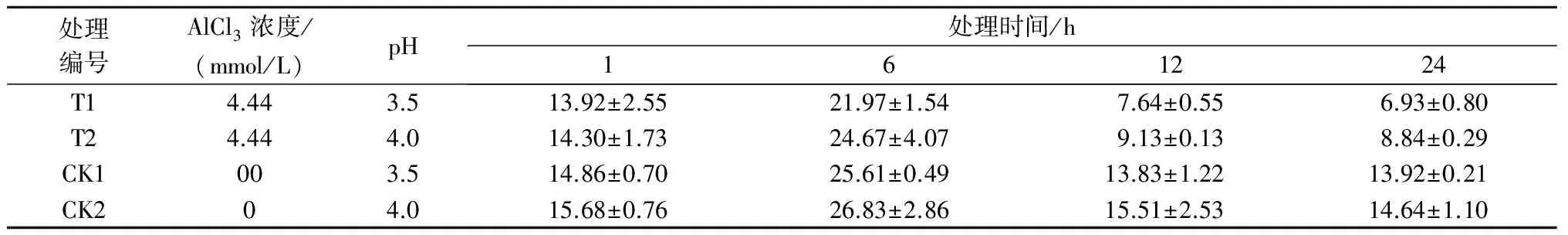
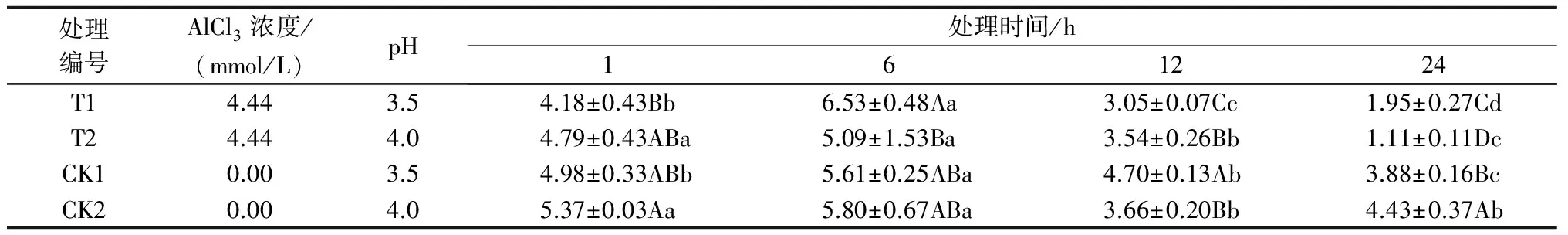
2.3 不同铝处理及处理时间对巨尾桉9号叶绿素荧光参数的影响
3 结论与讨论
