植被退化对尕海湿地枯落物分解的影响
2018-03-29马维伟赵锦梅陈国鹏朱海栋
马维伟, 李 广, 王 立, 赵锦梅, 陈国鹏, 朱海栋
(1.甘肃农业大学 林学院, 甘肃 兰州 730070; 2.尕海则岔自然保护区管理局, 甘肃 碌曲 747200)
枯落物分解是湿地生态系统养分和碳循环的关键环节之一,是土壤有机质主要来源,其分解过程对维持湿地生态系统正常生物地球化学循环和养分平衡等方面起着重作用[1-2]。枯落物分解过程主要受生物因素(土壤动物、微生物、植物群落等)、湿地所处的物理和化学条件(如土壤水分、温度)及枯落物本身性质(如枯落物物种和化学组成)[3-6]等因素影响。植被状况是湿地生态系统功能变化的主要体现者,其退化过程会显著改变所处土壤环境条件、植被生产力及其有机质的积累和分解速率,进而影响到生态系统碳氮循环[7],因此,研究植被退化过程中湿地枯落物分解特征对全面理解高寒湿地碳氮循环变化趋势具有重要意义。
尕海湿地位于全球气候变化最为敏感的地区——青藏高原东部,是中国独有青藏高原湿地重要组成部分,为国家重点保护湿地。在气候变暖和人为因素叠加影响下,尕海湿地植物普遍退化,原有植被盖度显著降低,一些湿生植物逐渐消失,中、旱生毒杂草逐渐增加,同时,植物生产力和土壤水分条件显著降低[8-10],这种变化不仅影响着湿地植物的生态过程,而且对地枯落物产生深刻影响。研究表明草地植被盖度下降,枯落物分解速率降低[3];也有研究认为,草地植被盖度下降,土壤温度上升,枯落物分解速率加快[6],但在较为干旱地区,温度上升也使得土壤水分散失加剧,从而对枯落物分解产生抑制作用。尽管这些关于草地盖度下降对枯落物分解影响的研究,一定程度上为高寒湿地植被退化过程中枯落物分解分析提供了启示,但对高寒湿地植被退化过程中枯落物分解特征尚不清楚,这限制了对高寒湿地枯落物分解机制的深入认识。因此,本文拟以尕海高寒湿地作为研究对象,采用时空替代法,研究泥炭沼泽和沼泽化草甸枯落物早期分解过程对植被退化的响应,并探讨气候对枯落物分解的影响。以期为深入理解环境变化条件下高寒湿地枯落物分解过程的响应机制提供数据支持,进一步认识和估计该区域内湿地物质循环与能量流动。
1 研究区域与研究方法
1.1 研究区概况
尕海湿地位于青藏高原东北边缘甘肃省碌曲县尕海—则岔国家级自然保护区内(33°58′12″—34°32′16″N,102°05′00″—102°47′39″E),海拔3 430~4 300 m。根据全国第二次湿地普查显示,保护区内湿地面积57 846 hm2,其中永久性河流1 675 hm2,季节性河流279 hm2,永久性淡水湖4 732 hm2,沼泽化草甸51 160 hm2[8]。该地区属于青藏高原气候带、高寒湿润气候区,年均气温1.2 ℃,年平均日较差13.7 ℃,气温以7月最高,平均为10.5 ℃;1月最低,平均为-9.1 ℃,无绝对无霜期,年均降水量781.8 mm,降雨通常集中在7—9月;年蒸发量1 150.5 mm。土壤类型主要包括暗色草甸土、沼泽土和泥炭土等。植物种类以乌拉苔草(Carexmeyeriana)、唐松草(Thalictrumaquilegifolium)、线叶蒿(Arternisiasubulata)、问荆(Equisetumarvense)、珠芽蓼(Polygonumviviparum)、蕨麻(Potentillaanserina)、散穗早熟禾(Poasubfastigiata)、青藏苔草(Carexmoorcroftii)、棘豆(Oxytropisochrocephala)等。
1.2 研究方法
1.2.1 样地设置 植被退化是指由于植物群落受到干扰而发生的植被逆向演替过程,主要指在人为活动或不利自然因素所引起的植被质量衰退,生产力、经济潜力及服务功能降低,环境变劣以及生物多样性或复杂程度降低,恢复功能减弱或失去恢复功能等[11-12]。植被退化造成植被盖度和生物量减少是高寒草甸湿地退化的明显特征。随着高寒草甸湿地退化,其湿生植物优势种群组成逐渐减少,植被的正常生长发育受到抑制,稳定的物质平衡受到破坏,植被变得低矮稀疏,组成结构改变,盖度减少,产草量下降。尕海湿地植被退化由原生湿地为中心向外逐渐扩展。本研究通过对尕海湿地生态环境和植被特征实地调查及相关资料分析基础上,选择刘惠斌等[13]确定的退化泥炭沼泽区贡巴点和退化沼泽化草甸区尕海湖周边。采用空间序列代替时间序列方法,以现在生长季可见的原生湿地为中心向外延伸的办法,选择地势相对平缓坡向一致的地段,根据尕海湿地植物种类组成、地上生物量、群落高度、群落盖度等指标,将泥炭沼泽(阳坡)和沼泽化草甸(较平坦地段)划分为不同的植被退化梯度,具体将泥炭沼泽划分为未退化(PⅠ)、退化(PⅡ)类型(按照调查区泥炭沼泽实际布设为未退化和退化两种类型),沼泽化草甸划分为未退化(SⅠ)、轻度退化(SⅡ)、中度退化(SⅢ)及重度退化(SⅣ)4种退化类型,样地植物基本特征(见表1)[8]。

表1 样地基本情况描述
1.2.2 土壤采样与样品分析
(1) 枯落物分解试验。试验分别在2013,2014,2016年生长季(5—9月)进行。采用分解袋法,分解袋由孔径为1 mm孔径的尼龙网制成,孔袋3面用尼龙线缝合,一面开口,有效面积为11 cm×11 cm。枯落物收集分别于每年4月中旬进行,为了降低压覆对枯落物破碎作用的影响,本试验采用现有植物立枯体。在泥炭沼泽和沼泽化草甸各退化类型样地内收集其优势种的立枯体,带回实验室剪成10 cm左右的小段,装进分解袋,每袋装15 g(烘干重),每种退化梯度枯落物分装12袋。分别于每年的5月中旬将制好枯落物分解袋(每年各60代)依次投放到上述不同植被退化梯度的2类湿地中。为了保持试验尽量与自然环境保持一致,分解袋置于土壤表层5—10 cm处。在随后各年的6,7,8,9月中下旬分别回收分解袋,每种退化类型分解袋收回3个,每月共计收回15个(沼泽化草甸重度退化阶段由于植被稀少,枯落物输入较少,未布设分解袋)。将分解袋带回实验室拣出植物根系、苔鲜及泥土颗粒,70 ℃烘干至恒重。采用赛多利斯电子天平BAS2 245(d=0.1 mg)称量,气温和降雨采用尕海湿地保护站气象观测站数据。
(2) 数据处理与分析。枯落物干物质残留率(RW)用公式:
RW=Wt/W0·100%
(1)
式中:RW——枯落物干物质残留率(%);Wt——t时刻枯落物干重(g);W0——枯落物的初始干重(g)。
枯落物干物质残留率变化用Olson单项指数模型:
y=ae-kt,y=Wt/W0
(2)
式中:a——拟合参数;k——分解系数;t——时间。
采用Excel 2007进行统计分析与作图,运用Spss 17.0进行差异显著性检验,文中泥炭沼泽枯落物残留率季节变化、分解速率为2013,2014年2 a数据的均值,沼泽化草甸为2013,2014,2016年3 a数据均值。
2 结果与分析
2.1 气候因子变化特征
2013,2014,2016年研究期间5—9月平均温度分别是9.7,9.3,10.3 ℃,显著高于5—9月的多年平均温度(8.3 ℃)。观测期间温度的时间变化与历史同期变化趋势一致(图1)。
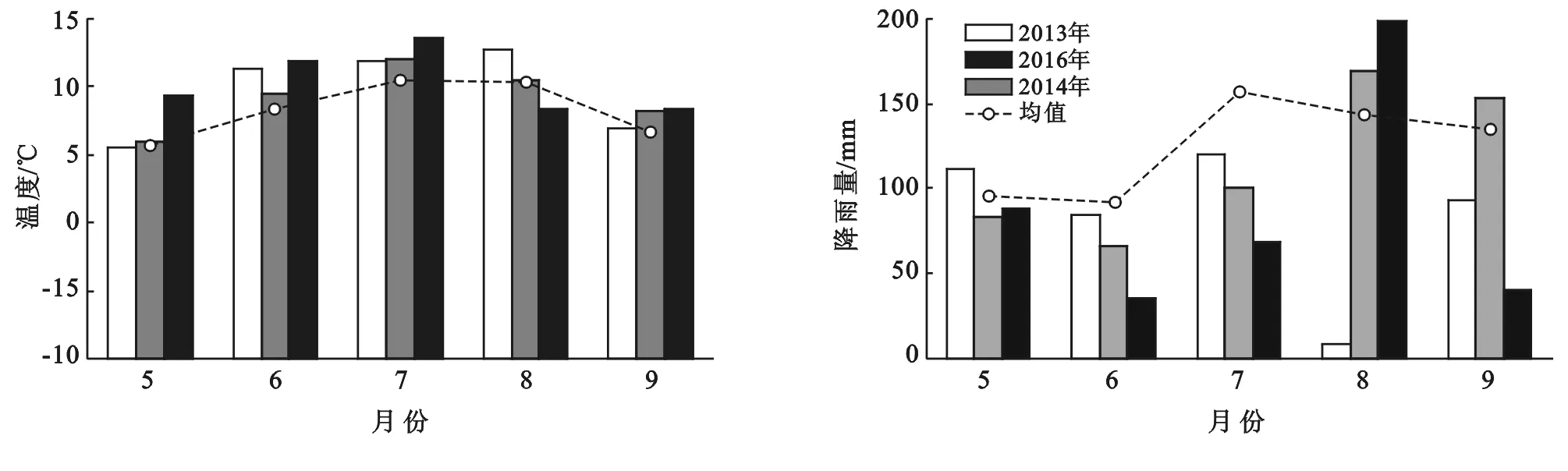
图1 尕海湿地2013,2014,2016年5-9月的气候特征(均值为5-9月的多年平均值)
然而,降雨的年内和年际变化非常明显,降雨和温度这种变化一定程度上影响了尕海湿地枯落物分解过程。2013,2104年,2016年5—9月降雨量分别为419.0,572.0和432.3 mm,而5—9月多年平均降雨量为621.9 mm,2013,2016年降雨量显著小于多年平均降雨量,分别是多年的0.67,0.70倍,而2014年降雨量是多年0.92倍。这表明2013,2016年是相对干旱年份,而2016年接近历史同期降雨。降雨的时间变化在在观测期不同年份也不相同,2013年最大降雨出现在7月,2014,2016年最大降雨在8月,但总体上与多年降雨量分布基本相同,主要集中在7—9月。
一般来说,降雨如果充足,枯落物分解较快,而2013,2016年降雨相对较少,这可能对植被退化湿地枯落物分解产生显著影响。
2.2 枯落物残留率季节变化
植被退化对尕海两类湿地的枯落物分解过程均有显著影响,但随植被退化演替,各湿地枯落物分解过程呈现出不同变化趋势(图2)。在沼泽泥炭地,植被未退化枯落物残留率均小于植被退化地,且随着时间延长,差异逐渐增大,到121 d时,植被未退化、退化枯落物残留率分别为68.01%和80.74%,未退化比退化减少了12.73%,说明植被退化抑制了沼泽泥炭地枯落分解过程,使得沼泽泥炭枯落物残留量显著增加。
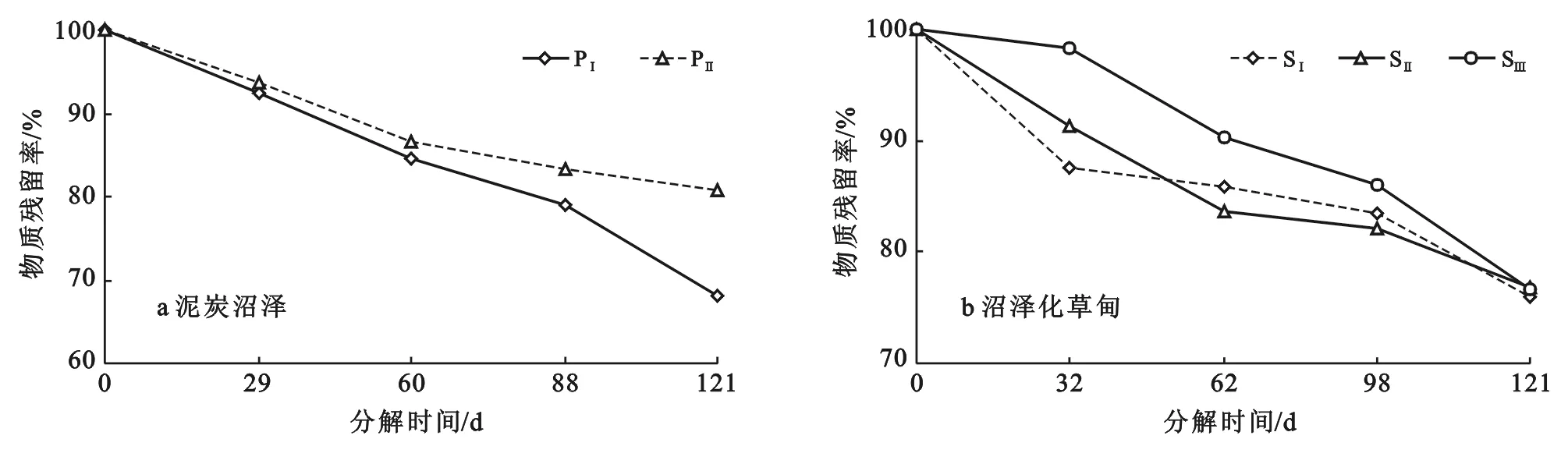
注:PⅠ为泥炭沼泽未退化; PⅡ为泥炭沼泽退化; SⅠ为沼泽化草甸未退化; SⅡ为沼泽化草甸轻度退化; SⅢ为沼泽化草甸中度退化。下同。
图2不同退化梯度枯落物残留率季节变化
在沼泽化草甸地,未退化枯落物样品在分解前期32 d时显著高于中度退化(p<0.05),但与轻度退化差异不显著(p>0.05),而随着分解时间延长,未退化、轻度退化和中度退化枯落物残留率趋于一致,到121 d时,未退化、轻度退化和中度退化枯落物残留率分别是75.94%,76.89%和76.69%。以上研究结果表明,植被退化对沼泽化草甸枯落物分解表现出明显抑制作用,但抑制作用随时间增加逐渐减小。
2.3 枯落物分解速率变化
在120 d的分解期间,植被退化对尕海2类湿地枯落物分解系数影响较小(表2),但延长了枯落物分解时间。在沼泽泥炭地,植被未退化的枯落物分解95%时间为2.75 a,而植被退化为4.07 a,退化比未退化分解时间延长了1.32 a。但在沼泽化草甸,植被退化对枯落物分解时间延长较小,枯落物分解95%的时间,中度退化、轻度退化分别比未退化延长0.01,0.07 a。

表2 枯落物物质残留率(y=Wt/W0)与分解天数(t)间的指数方程及相应参数
对尕海两类湿地的0~120 d枯落物分解速率进行计算,发现2类湿地枯落物分解速率受环境及枯落物本身性质变化,在不同分解时段内,枯落物分解速率存在明显差异(见表3)。
由表3可以看出,生长季内沼泽泥炭植被未退化枯落物分解速率明显高于退化,但差异不显著(p<0.05);分解速率前90 d内,随时间变化趋势大致相同,而在90~120 d,未退化显著增大,退化显著减小。生长季内沼泽化草甸0~120 d内平均分解速率顺序为:未退化(0.028 9 g/d)>中度退化(0.028 7 g/d)>轻度退化(0.028 0 g/d),差异不显著(p>0.05);未退化和轻度退化分解速率随时间变化趋势大致相同,其中0~30 d内两种退化的分解速率显著高于其他时段,中度退化分解速率随时间变化呈波动性变化,而30~60和90~120 d内分解速率高于其他时间。
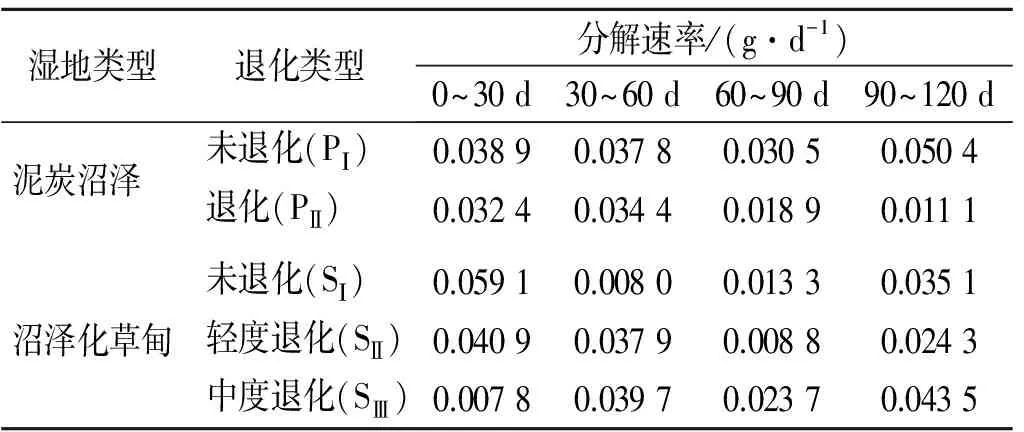
表3 尕海湿地不同植被退化条件下枯落物分解速率变化
2.4 枯落物残留率年际变化
在试验期间,各植被退化阶段的湿地枯落物分解过程具有明显年际变化特征(图3)。在泥炭沼泽地,2013,2014年各植被退化阶段湿地的分解过程变化趋势相同,但2014年枯落物残留率明显低于2013年,如2013年未退化在121 d时残留率为70.72%,而在2014年为65.31%;从枯落物残留率的年际分异来看(表4),未退化变异系数在1.63%~5.62%之间,而退化变异系数在0.27%~3.65%,未退化年际分异较大。而在沼泽花草甸地,2013,2014年,2016年分解过程差异较小,但总体表现为2014年分解较快,其他两年相对较慢;未退化、轻度和中度退化变异系数变化范围分别为2.01%~4.40%,1.18%~10.18%和0.35%~1.75%,轻度退化年际变化最为显著。
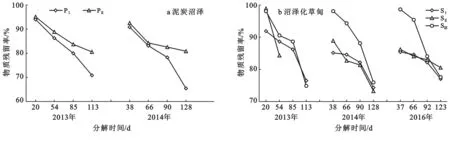
图3 尕海湿地不同退化梯度枯落物残留率年际变化

湿地类型退化类型变异系数/%5—6月6—7月7—8月8—9月泥炭沼泽 未退化2.362.621.635.62退化1.933.650.790.27未退化4.402.682.762.01沼泽化草甸轻度退化7.301.182.1010.11中度退化0.352.722.931.75
2.5 气候因子对枯落物分解的影响分析
从表5可以看出,各植被退化阶段枯落物分解与气候因子相关分析可以看出,泥炭沼泽地植被未退化地枯落物分解速率与降雨、温度呈明显正相关,而植被退化枯落物分解速率与温度正相关,与降雨呈负相关,但相关性均未达到显著水平。由此看出,水分条件是影响植被退化泥炭沼泽枯落物分解的主要因子。而在沼泽化草甸,除未退化和中度退化枯落物分解速率与降雨呈负相关,其他均呈正相关,尤其在未退化地,温度与分解速率呈极显著正相关,说明植被退化会导致影响枯落物分解的主要气候因子发生了改变。

表5 尕海湿地枯落物分解速率与环境因子相关系数
3 讨 论
近年来,受气候变化、过度放牧等作用,尕海高寒湿地植被出现不同程度退化[8,14],湿地植物群落的物种组成及枯落物的积累发生改变,使得群落中与凋落物分解相关的植物功能特征组合(如高纤维物种比例等)发生变化,进而导致凋落物质量和分解特征改变[15]。同时,植被退化引起湿地原有环境恶化(如土壤紧实度增加,土壤养分、水分减少等),导致枯落物分解特征发生变化。
在众多气候因素中,以气温和降水对分解过程的影响较为深刻。一般而言,温度与分解速率呈正相关,主要由于温度能够直接影响微生物活性和土壤动物的新陈代谢活动,致使分解速率随气温的升高而增加[16],本研究也进一步证实分解速率与温度呈正相关,但随着植被退化梯度增加,相关系数逐渐减小,可能由于植被退化导致湿地土壤水分和养分含量降低,消弱了温度对湿地枯落物分解的促进作用,这与张艳博等[3]对青藏高原高寒草地凋落物分解研究不尽一致。
降水是影响枯落物分解的另一重要气候因素。降水主要是通过影响淋溶过程和分解者活性而影响枯落物分解速率。本研究发现在泥炭沼泽地中,枯落物分解与降水量呈正相关,与Raija等[17]研究结果一致。虽然降水能够一定程度引起地面温度降低,这可能降低分解速率。但在不同湿地间的环境梯度,决定了影响分解的环境因子不同。本研究选取泥炭地在尕海泥炭沼泽的阳坡地区,气温回升快,全年平均气温相对较高,且为常年不淹水泥炭沼泽地,故水分条件成为分解过程的主要影响因素。沼泽化草甸地的植被未退化和中度退化枯落物分解速率与降水成负相关,这可能是在植被未退化沼泽化草甸中,水分含量相对饱和,一些研究也表明,饱和的水分形成的厌气条件反而会使生态系统的枯落物分解变慢[18],但在中度退化沼泽化草甸,枯落物分解与土壤水分成反比,可能由于枯落物本身基质和土壤环境条件的作用显著改变了降水对枯落物的分解作用,其原因需要进一步研究。总之,枯落物分解状况主要取决于气温和降水(湿度)的对比关系(湿热比),当温度相对于湿度很高时,分解迅速;反之,分解缓慢。
在气候变化背景下,过度放牧导致的植被退化对湿地枯落物分解有显著影响。本研究发现植被退化抑制了沼泽泥炭地枯落分解过程,增大了沼泽泥炭枯落物残留量,但对沼泽化草甸枯落物分解的抑制作用随时间增加而逐渐减小,这与先前对草地研究结论一致[3],虽然植被盖度降低有利于增加到达地表面辐射会加速枯落物分解,但在放牧导致湿地植被退化过程中,牲畜选择采食一些适口性高(养分含量高、粗纤维低)、易分解的植物,导致这些易分解植物物种逐步退出群落,而适口性低、难分解的物种在群落中的优势度将逐步增加,凋落物分解速度降低,由此构成负反馈调节。这也进一步表明尕海沼泽草甸湿地植被退化后,凋落物质量发生改变,分解速度开始加快,系统养分循环速度加快,可能引起湿地养分的输入逐渐减弱甚至出现净消耗,进而导致湿地退化加速,这与若尔盖湿地研究结果一致[17],也可能是近几十年来尕海高寒湿地呈加速退化的机制之一。
此外,植被退化导致地上生物量减少、植被覆盖度降低的同时,也使土壤紧实度增加、减少了土壤水分的蒸散。在高寒湿地,随着温度的上升,微生物的呼吸作用和土壤动物的活动性显著提高[3],一定程度上加速枯落物分解。而本研究发现却呈相反变化特征,为植被退化样地枯落物分解速率小于未退化,这说明枯落物分解除受自身性质影响外,还可能受到其他因素影响,如植被退化增加了土壤紧实度,限制O2交换速率,降低酚氧化酶的活性,而对湿地凋落物分解产生抑制,同时植被退化湿地水分、养分等立地条件变差,降低枯落物分解速率[16]。
植被退化对不同湿地类型枯落物分解的影响程度也有所不同。本研究利用Olson单项指数模型发现沼泽泥炭地植被退化比未退化分解时间延长了1.32 a,沼泽化草甸植被中度退化、轻度退化分别比未退化延长0.01,0.07 a,这一结果说明泥炭沼泽湿地枯落物分解对植被退化响应更为敏感和强烈。虽然难以直接比较植被退化对泥炭沼泽和沼泽化草甸地中物理和生物环境改变的大小,但植被退化对沼泽草甸凋落物分解的影响较小这一结果,可部分归因于牲畜啃食牧草使牧草本身的生态生理特征发生改变,从而影响到凋落物的质量和分解。
4 结 论
(1) 植被退化对尕海湿地枯落物分解表现出抑制作用,但抑制作用有所不同。Olson单项指数模型显示沼泽泥炭地植被退化比未退化分解时间延长了1.32 a,沼泽化草甸中度退化、轻度退化分别比未退化延长0.01,0.07 a。
(2) 各植被退化阶段湿地枯落物分解明显不同,沼泽泥炭未退化分解速率明显高于退化,而沼泽化草甸分解速率排序为:未退化(0.028 9 g/d)>中度退化(0.028 7 g/d)>轻度退化(0.028 0 g/d)。各退化阶段湿地枯落物分解年际变化明显,在泥炭沼泽地,2014年枯落物残留率明显低于2013年,而在沼泽花草甸,2013,2014,2016年枯落物残留率差异较小,但总体表现为2014年相对较小,其他两年较高。
(3) 温度和降雨对枯落物相对分解速率具有促进作用,但在不同植被退化阶段表现的相关性不同,甚至表现为负相关。
[1] 侯翠翠,宋长春,李英臣,等.不同水分条件下毛果苔草枯落物分解及营养动态[J].生态学报,2012,32(2):650-658.
[2] 张雪梅,王永东,徐新文,等.塔里木沙漠公路防护林地表凋落物分解对施肥的响应[J].生态学报,2017,37(5):1506-1514.
[3] 张艳博,罗鹏,孙庚,等.放牧对青藏高原东部两种典型高寒草地类型凋落物分解的影响[J].生态学报,2012,32(15):4605-4617.
[4] Tiegs S D, Entrekin S A, Reeves G H, et al. Litter Decomposition, and Associated Invertebrate Communities, in Wetland Ponds of the Copper River Delta, Alaska(USA)[J]. Wetlands, 2013,33(1):1151-1163.
[5]Britson A, Wardrop D, Drohan P. Plant community composition as a driver of decomposition dynamics in riparian wetlands[J]. Wetlands ecology and management, 2016,24(3):335-346.
[6] Luo C Y, Xu G P, Chao Z G, et al. Effect of warming and grazing on litter mass loss and temperature sensitiveity of litter and dung mass loss on the Tibetan plateau[J]. Global Change Biology, 2010,16(5):1606-1617.
[7] 刘育红,李希来,李长慧,等.三江源区高寒草甸湿地植被退化与土壤有机碳损失[J].农业环境科报,2009,28(12):2559-2567.
[8] 马维伟,王辉,李广,等.甘南尕海湿地不同植被退化阶段土壤有机碳含量及动态[J].水土保持学报,2015,29(5):254-259.
[9] 马维伟,李广,石万里,等.甘肃尕海湿地退化过程中植物生物量及物种多样性变化动态[J].草地学报,2016,24(5):960-966.
[10] 马维伟,王辉,李广,等.甘南尕海湿地退化过程中植被生物量变化及其季节动态[J].生态学报,2017,37(15):5091-5101.
[11] 李新平,赵娟,安雁.植被退化评价研究展望[J].世界林业研究,2012,25(2):30-33.
[12] 王亚军,魏兴琉,杨萍.超载放牧对那曲地区高山篙草草甸植被退化的影响[J].兰州大学学报:自然科学版,2005,41(1):32-38.
[13] 刘惠斌,李俊臻.甘肃尕海湿地退化泥炭地恢复技术评价[J].湿地科学与管理,2010,6(2):26-29.
[14] 刘帆,刘晖,徐凌翔,等.青藏高原东部尕海湿地种子库沿水位梯度的分布格局[J].科学通报,2013,58(14):1332-1339.
[15] Garibaldi L A, Semmartin M, Chaneton E J. Grazing-induced changes in plant composition affect litter quality and nutrient cycling in flooding Pampa grasslands[J]. Oecologia, 2007,151(4):650-662.
[16] 武海涛,吕宪国,杨青,等.三江平原典型湿地枯落物早期分解过程及影响因素[J].生态学报,2007,27(10):4027-4035.
[17] Raija L, Jukka L, CarlC T, et al. Scots pine litter decomposition along drainage succession and soil nutrient gradients in peatland forests, and the effects of inter-annual weather variation[J]. Soil Biology and Biochemistry, 2004,36(7):1095-1109.
[18] 孙志高,刘景双,湿地枯落物分解及其对全球变化的响应[J].生态学报,2007,27(4):1606-1618.
