桂南木莲繁育系统及其传粉适应性1)
2018-03-27罗峰金晓玲李瑞雪张哲张方静胡希军
罗峰 金晓玲 李瑞雪 张哲 张方静 胡希军
(中南林业科技大学,长沙,410004) (湖南省环境资源植物开发与利用工程技术研究中心)
植物繁育系统一般指直接影响后代遗传组成的所有有性特征,包括花部综合特征、花各性器官寿命、开放式样、自交亲和程度和交配系统等,这些特征结合传粉者和传粉行为是影响生殖后代遗传组成和适合度的主要因素[1],对调节种群结构有重大影响[2]。植物对传粉者的传粉适应是被子植物花部特征进化的根本动力[3],如花瓣颜色[4]、花大小[5]、花形态[6]、花结构对称性[7-8]等花部特征的塑造都受其传粉昆虫的影响。此外,植物花部特征的形成也会受一些非传粉者因子的影响[9]。因此,研究植物繁育系统及传粉适应,是探讨物种多样性发生历史、维持机理和保护策略的重要基础[1]。
桂南木莲(Manglietiachingi)是木兰科(Magnoliaceae)木莲属(Manglietia)常绿乔木,花朵白色清香,具树形优美和材质优良等特点,是重要的观赏树种和林业用材[10]。但桂南木莲主要分布在海拔700~1 000 m的山林中,自然种群分布零散,自然条件下结实率低[11],严重制约了种群的自然繁衍和开发利用。目前,对桂南木莲的研究主要有引种栽培[10]和种子繁育[12],对其繁育系统的研究尚未开展。为探讨桂南木莲结实率低下的原因,文中对其繁育系统进行研究,包括开花生物学观察、花粉活力与柱头可授性的测定、花粉/胚珠比值的估算以及传粉昆虫传粉行为的观察。阐明桂南木莲开花特征的传粉适应机制,为桂南木莲的资源保护和开发利用提供科学的依据。
1 材料与方法
1.1 材料
试验材料是中南林业科技大学植物园中的桂南木莲人工种群,植物园位于111°54′~114°15′E,27°51′~28°40′N,海拔100~120 m,属于亚热带湿润气候,地带性植物为常绿阔叶林,年均降水量1 361.6 mm。园内植物种类丰富,有桂南木莲常见的野外伴生树种,如壳斗科、樟科、安息香科、金缕梅科等植物。
1.2 方法
开花生物学观察:在2016—2017年每年4—6月份,参照Dafni[13]201-211方法选择10株10~15年生植株进行观察。盛花期(全株50%以上花朵开放)时随机标记20朵花朵,观察花瓣、雌蕊、雄蕊的变化过程,单花开花动态,计算单花花期。
柱头可授性测定:采用联苯胺-过氧化氢法测定不同开花阶段柱头的可授性。
花粉活力测定:用TTC染色法检测不同开花阶段花粉活力;参考付玉嫔等[14]试验结果,采用培养基萌发法测定闭合阶段花粉活力。分别以不同质量浓度的蔗糖和硼酸混合溶液为培养基,并设清水为对照,共10组处理(表1),在培养箱中25 ℃暗培养花粉10 h,用光学显微镜拍照后统计花粉萌发率。

表1 花粉萌发不同培养基组合
访花昆虫的观察:选择晴天对盛花期花朵的访花昆虫进行全天候观察。
杂交指数(OC,I)的测定:盛花期时随机选择闭合期的30朵花,按照Dafni[13]201-211标准测量花部特征。
花粉/胚珠比的估算:随机选取10朵未散粉花朵,将雄蕊取下分别装于洁净的小瓶中,待花粉全部散出后加入10 mL蒸馏水,磁力搅拌器下振荡30 min,取1 μL滴载玻片上,在光学显微镜下统计全部花粉数。同时将子房壁用解剖刀切开,统计单花胚珠数值。最后计算出P/O的值,参照Cruden[15]标准判定其繁育系统。
人工控制性授粉试验:设置5个处理,即对照,为不套袋,不去雄,自由传粉,检测自然条件下结实情况;去雄,为套袋,检测是否存在无融合生殖;不去雄,为套袋,检测是否能自花授粉;人工同株异花授粉,为检测自交是否亲和;异株异花授粉,为检测异交是否亲和。果实成熟后统计座果率和结籽率。座果率=(结实总数/处理个数)×100%;结籽率=(果实中种子数/胚珠总数)×100%。
1.3 数据处理
数据采用Spss软件进行方差分析和Duncan多重比较,用Excel作图。
2 结果与分析
2.1 开花物候及开花动态
2.1.1 开花物候
桂南木莲于4月上旬芽鳞开裂(图1A)新叶生长,10~12 d后形成花蕾,随后外苞片开裂脱落(图1B),18~22 d花朵开放。其群体花期从4月底持续到6月初,单株花期(31±5)d,若遇连续睛天植株迅速进行入盛花期。
2.1.2 开花动态
桂南木莲单朵花期3~4 d,可分为5个阶段,即松蕊阶段,时间08:00—19:00,外轮花被片开裂,露出白色中轮花瓣(图1C);初次开放阶段,时间19:00—21:00,柱头分泌较多黏液(图1D);闭合阶段,21:00—翌日19:00,外轮花被片舒展但内3轮合拢,花药开裂但雄蕊紧贴雌蕊基部,雄蕊并未对外散粉(图1E);再次开放阶段,时间翌日19:00—21:00,内3轮花被片再次打开,花药开始对外散粉(图1F);衰败阶段,前期内3轮花被片合拢但相互之间留有空隙,大量花粉散落在花被片底部(图1G),后期花药和花被片脱落,柱头变褐色失去活性(图1H)。
2.2 柱头可授性测定及雄蕊散粉情况
桂南木莲柱头可授性及雄蕊群散粉情况表明(表2),5个开花时阶段柱头都有可授性。其中闭合阶段柱头可授性最强(图2C),持续时间较长,约16 h,但花朵闭合不利于昆虫授粉;初次开放阶段柱头可授性强(图2B),但持续时间短暂,约3 h,且在传粉昆虫活动少的夜晚;再次开放阶段柱头可授性较强(图2D),雄蕊对外散粉,柱头位于雄蕊群下方,能授上自花花粉。

表2 桂南木莲柱头可授性及雄蕊群散粉情况
注:“++++”表示柱头可授性最强;“+++”表示柱头可授性强;“++”表示柱头可授性较强;“+”表示柱头可授;“-”表示柱头不可授。

A.芽鳞开裂;B.花蕾外苞片脱落;C.松蕊阶段;D.初次开放阶段;E.闭合阶段;F.再次开放阶段;G.衰败前期;H衰败后期。
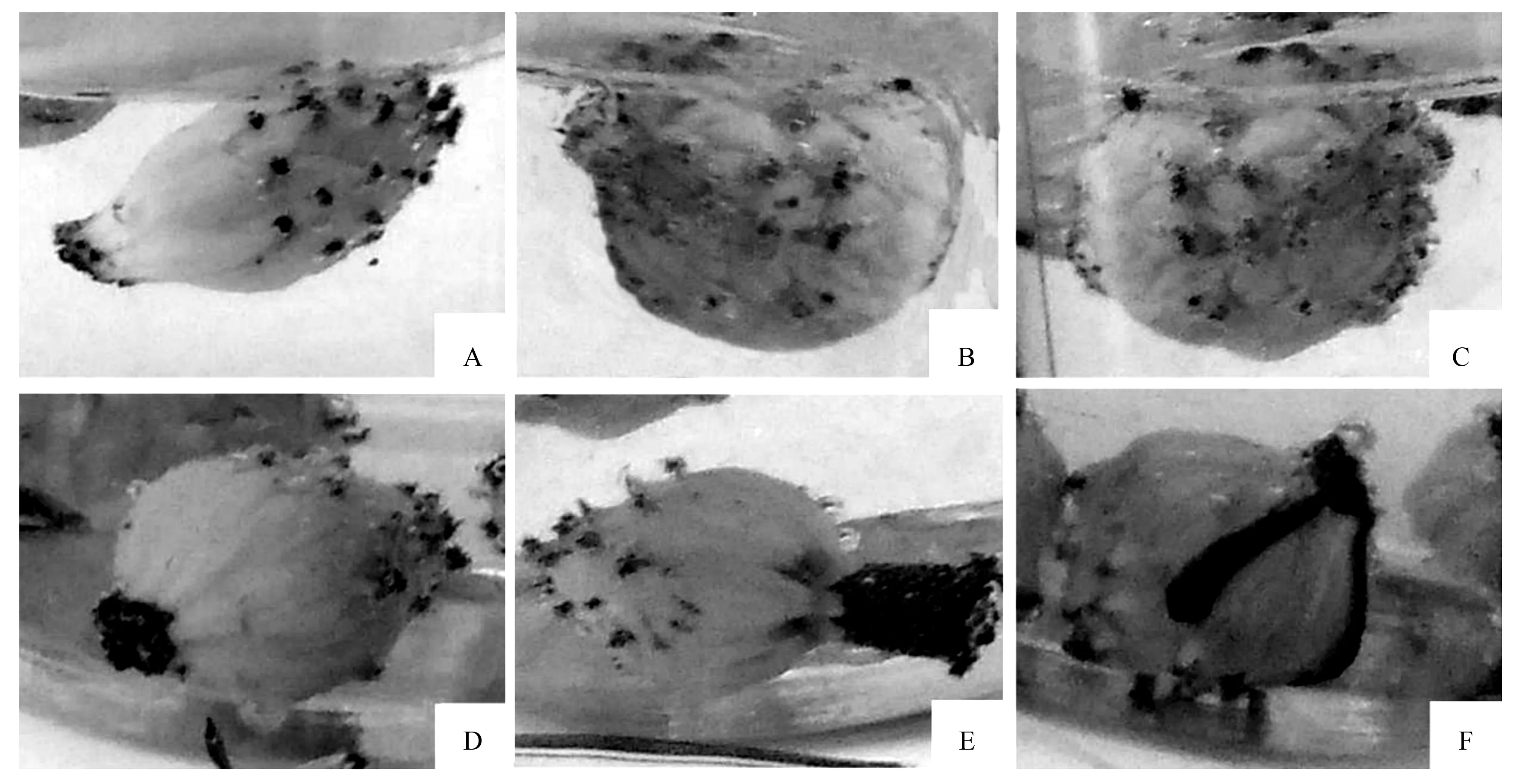
A.松蕊期;B.初次开放阶段;C.闭合阶段;D.再次开放阶段;E.衰败前期;F.衰败后期。
2.3 花粉活力测定
由TTC染色法测定桂南木莲花粉活力可知(表3),桂南木莲花粉活力随着开花进程逐渐增强,在闭合阶段花药开裂时花粉活力最高为31.34%,随后花粉活力下降;在再次开放阶段雄蕊群对外散粉时花粉活力较低,为8.85%;衰败前期花粉活力仅为1.93%,花粉活力散失快。此外,用花粉萌发法测定闭合期花粉最高萌发率也仅为26.23%,与其他木兰属和含笑属植物相比桂南木莲花粉活力低。

表3 TTC染色法测定桂南木莲花粉活力
注:表中花粉染色率数据为平均值±标准差;不同字母表示差异显著(P<0.05)。
2.4 访花昆虫的观察
通过对桂南木莲访花昆虫的观察,发现白天蜜蜂是主要访花昆虫,且主要访问闭合阶段和衰败前阶段期的花朵。蜜蜂访花时从花瓣间隙进入花朵(图3A),或从花瓣底部钻入(图3B),身上沾满花粉后再访问其他桂南木莲花朵时进行授粉。极少数甲虫类昆虫是桂南木莲夜间访问者,主要访问初次开放和再次开放的花朵,尤其再次开放花朵散出的花粉正好通过这些甲虫的访花活动授到其他花朵的柱头上,同时还发现甲虫啃食花瓣的现象(图3C)。此外,在桂南木莲花内还发现蚂蚁、蓟马等小型昆虫,据观察,这类昆虫不是桂南木莲的有效传粉昆虫。
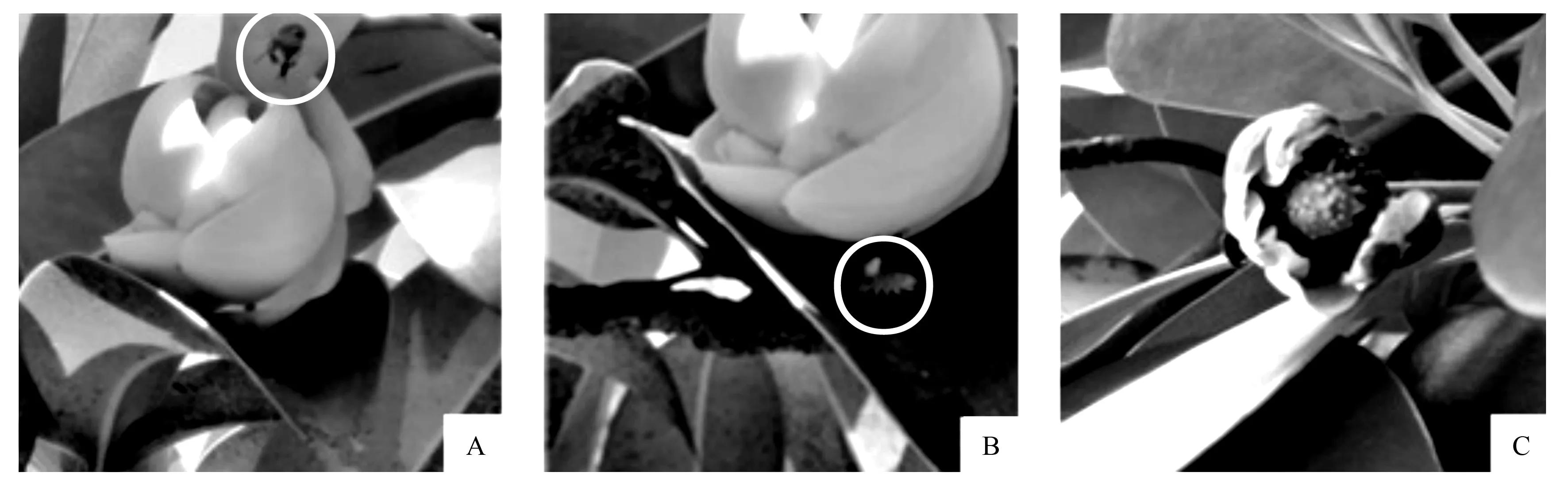
A.蜜蜂从花瓣间隙飞入;B.蜜蜂从花瓣底部钻入;C.被甲虫啃食的花朵。
2.5 杂交指数的测定
参照Dafni[13]201-211的标准,桂南木莲花朵直径(6.2±1.5)cm>6 mm,记为3;花为两性花,雌蕊先熟,记为0;雌雄蕊空间分离,且花药高于柱头,记为1。因此,桂南木莲的杂交指数OC,I为4,繁育系统为部分自交亲和,异交,需要传粉者。
2.6 花粉/胚珠比的估算
桂南木莲的单花平均花粉数为3 999 500±1 530,胚珠平均数为321.75±15,平均P/O=12 430±120(在2 108.0~195 525.0之间),判定其繁育系统为专性异交。
2.7 套袋试验
桂南木莲套袋处理结实表明(表4),桂南木莲自然传粉条件下座果率和结籽率均较低,分别为15.00%和2.29%±0.13%,小核果败育严重(图4A);去雄、套袋不结实,说明不存在无融合生殖现象;不去雄、套袋和同株异花授粉的结实率为0,说明自交不亲和(图4B);桂南木莲异株异花授粉座果率较高,为65.00%,果实发育良好(图4C),但结籽率仅为11.32%±1.06/%,通过解剖果实发现心皮内胚珠败育严重,单个心皮内种子2~3粒。由此判断桂南木莲的繁育系统为异交型,需要传粉媒介。
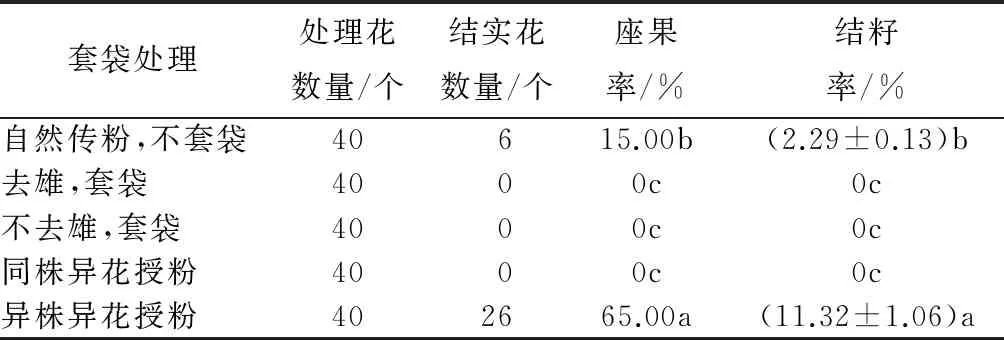
表4 桂南木莲套袋处理结实情况
注:表中结籽率数据为平均值±标准差;同列不同字母表示差异显著(P<0.05)。

A.自然传粉,不套袋发育的果实;B.同株异花授粉不亲和,雄蕊败育;C异株异花授粉发育的果实;D.单心皮内两粒种子,胚珠败育严重。
3 结论与讨论
异交传粉机制的多样性是花结构和传粉媒介相互适应的结果。为适应昆虫的传粉,植物形成与授粉昆虫活动规律相协调的开花动态。例如,高山龙胆科植物为节约能量以适应低频率的昆虫访花,形成了有规律的昼夜开放和闭合的特征及时间长的花寿命[16]。克鲁兹王莲花器官呈现出有规律的开放和闭合,也是适应其传粉昆虫的需要[17]。Gerhard et al.[18]观察到巴西木兰科植物Magnoliaovata夜晚开放、二次开合的开花特征,发现这与其特定授粉甲虫在晚上活动有关,且夜晚雌蕊特征和雄蕊特征表达时花朵所散发的香味只能吸引这种甲虫访花。本研究中,桂南木莲两次开放都是在上半夜进行,初次开放阶段柱头可授性较强,雄蕊未散粉,可为异交提供无污染的柱头,再次开放阶段雄蕊群才开展散粉则为异交提供较高活力的花粉,试验中观察到的甲虫正好是花粉传播的媒介。此外,桂南木莲在连续天晴时迅速进入盛花期,更是为这种传粉机制实现生殖成功提供柱头和花粉的保障。而桂南木莲花朵下半夜闭合,从而推测其目的是,节约能量、保护雌蕊和为授在柱头上的花粉萌发形成合适的微环境。
花朝向的花部特征也是植物与环境相适应的结果,其中传粉者在花朝向的形成中扮演着重要角色[9]。植物会通过花朵下垂来提高昆虫访花频率,提高授粉成功率[19],同时避免花粉因阳光直射而降低活力,延长雄性特征表达时间[20]。本研究中人工套袋试验结果显示桂南木莲存在自交不亲和现象,结合较低的花粉活力测定结果和白天蜜蜂的访花行为,判断桂南木莲花朵下垂、花被片合拢的花部特征并不是为其自交提供条件,而是为了将花粉散落在合拢的花被片上,延长雄性特征表达时间,这也是为昆虫传粉提供充足的花粉源。综上所述,桂南木莲花朵下垂、二次开合的花部特征均是对其昆虫传粉机制的适应。
桂南木莲花量大,连续晴天时进入盛花期,在单株和群体水平上开花同步性强,属于“大量、集中式开放”模式,这种模式能吸引更多的昆虫访花传粉,保障植株的生殖成功。同时桂南木莲群体花期在雨水多的季节,这种集中开放模式还能减轻恶劣天气对其群体繁衍的不利影响。但桂南木莲为避免自交导致的败育,雄蕊特意延迟对外散粉时间,因此在单花水平上开花同步性差。另外,桂南木莲单花虽然能产生大量花粉,但在柱头可授性最强期间,柱头被花被片包围,难以接受虫媒传粉以外的其他途径授粉,因此,受花粉传递的限制,在自然条件下植株的结实率低。
在测定其繁育系统时,花粉/胚珠比和套袋试验测定的结果与杂交指数测定的结果不一致。这可能与用杂交指数测定植物繁育系统时,只考虑到花的外部特征对植物生殖系统的影响,对授粉后花粉萌发和生长、雌雄配子结合及胚胎发育等未做考虑有关。桂南木莲作为原始的被子植物,开花过程中花被片长期包裹雄蕊和雌蕊以及雄蕊位于雌蕊之上的花结构,很容易发生自花授粉造成胚胎败育。且自交谱系缺乏对新环境的适应性和形成新物种的潜力,易引起自交衰退而导致物种灭绝,所以自交不能作为植物长期的生殖策略[16]。因此,在种群长期的繁衍中,桂南木莲为避免自交衰退依旧保持原始专性异交的繁育系统。
此外,本文研究结果与测定大果木莲(Manglietiagradis)繁育系统时的结果有所不同,这说明同一分类群(种、属、族、科)中繁育系统会产生多样性表达,从而推测是花部特征不同的植物(桂南木莲花下垂合拢,大果木莲花上举开放)为适应传粉环境形成的不同适应机制。
综上所述,桂南木莲的繁育系统是异交,昆虫传粉,有效传粉昆虫是甲虫和蜜蜂。在自然条件下其结实率低的内因是对外散出的花粉活力低,外因是缺少有效的传粉昆虫。
桂南木莲结实率低与其专性异交的繁育系统密切相关。花粉活力低,需要传粉媒介,自然条件下种群多星散分布,这些都会影响传粉的有效性[21],制约桂南木莲自然种群的繁衍更新。因此,应加强对其人工栽培技术的研究,通过人工栽种的手段扩大种群数量,结合花期释放传粉昆虫的方法,提高自然条件下桂南木莲异株异花授粉的概率。此外,还应加强对现有桂南木莲自然种群的保护,减轻人为活动对种群的影响,进而达到种质资源保护的目的。
[1] 何亚平,刘建全.植物繁育系统研究的最新进展和评述[J].植物生态学报,2003,49(2):151-163.
[2] 杨旭,杨志玲,王洁,等.濒危植物凹叶厚朴的花部综合特征和繁育系统[J].生态学杂志,2012,31(3):551-556.
[3] 黄双全,郭友好.传粉生物学的研究进展[J].科学通报,2000,51(3):225-237.
[4] HEUSCHEN B, GUMBERT A, LUNAU K. A generalised mimicry system involving angiosperm flower colour,pollen and bumblebee’s innate colour perferences[J]. Plant Systematics & Evolution,2005,252(3):121-137.
[5] THOMPOSON J D. How to visitation patterns vary among pollinators in reation to floral display design in a generalist polliantion systen?[J]. Oecologia,2001,126(3):386-394.
[6] BLOCH D, ERHARDT A. Selection toward shorter flowers by butterfies whose probosces are shorter than f-loral tubes[J]. Ecology,2008,89(9):2453-2460.
[7] NEAL P R, DAFIN A, GIURFA M. Floral sysmetry and its role in plant-pollinator systems: terminology, distr-ibution, and bypotheses[J]. Annual Review of Ecology & Systematics,2003,29(1):345-379.
[8] SARGENT R D. Floral symmetry affects speciation rates in angiosperms[J]. Proceedings of the Royal Society Biological Sciences,2004,271:603-608.
[9] 侯勤正,任昱兰,文静,等.灌木铁线莲繁育系统特征及其花下垂现象的适应性意义[J].西北植物学报,2016,36(11):2283-2290.
[10] 徐奎源,徐永星.桂南木莲的引种与栽培技术[J].林业实用技术,2004,47(10):41-42.
[11] 罗仲春.桂南木莲[N].中国花卉报,2003-10-07(T00).
[12] 杨成华,孔志红.9种木兰科种子的场圃发芽率试验[J].贵州林业科技,2003,31(3):19-21,43.
[13] DAFNIA.Pollinationecology:Apracticalapproach[M].Oxford:OxfordUnivertyPress,1992.
[14] 付玉嫔,陈少瑜,吴涛.濒危植物大果木莲与中缅木莲的花部特征及繁育系统比较[J].东北林业大学学报,2010,38(4):6-10.
[15] GRUDEN R W. Pollen-ovule: a conservative indicator of breeding systems in flowering Plants[J]. Evolution,1977,35(1):1-6.
[16] 何亚平,刘建全.植物繁育系统研究的最新进展和评述[J].植物生态学报,2003,49(2):151-163.
[17] 周庆源,傅德志,靳晓白.克鲁兹王莲(睡莲科)的开花生物学研究[J].西北植物学报,2006,27(8):1526-1533.
[18] GERHARD G, ILSE S, ROGER S, et al. Pollination ecology ofMagnoliaovatamay explain the overall lar-ge flower size of the genus[J]. Flora,2012,207(2):107-118.
[19] SUNS G, YAOCY Y. Increased seed set in down slope-facing flower ofLiliumduchartrei[J]. Journal of Systematics & Evolution,2013,51(4):405-412.
[20] CHAN Z, IRWIN R E, YUN W, et al. Selective seed abortion induced by nectar robbing in the selfing pl-antComastomapulmarium[J]. New Phytologist,2011,192(1):249-255.
[21] 李宗艳,郭荣.木莲属濒危植物致濒原因及繁殖生物学研究进展[J].生命科学研究,2014,18(1):90-94.
