横断山区湿地优势植物水葱种子繁殖性状对环境变化的响应1)
2018-03-27王志保田昆管东旭李晖张依南冯春慧孙梅
王志保 田昆 管东旭 李晖 张依南 冯春慧 孙梅
(国家高原湿地研究中心(西南林业大学),昆明,650224)
种子繁殖是植物种群维持和扩散的主要途径之一[1],环境变化很容易引起植物种子繁殖结果的改变,导致物种间的竞争共生关系发生改变,对群落的物种组成和生态系统产生巨大的影响[2-5]。种子繁殖对环境条件变化十分敏感,加强植物种子繁殖对环境变化的适应性研究是了解植物生存和发展动态、预测未来植物群落结构变化的基础。过去很多研究发现,环境变化显著影响植物种子繁殖,但不同研究得到的结果不尽相同[6-10]。例如,栓皮栎(Quercusvariabilis)的种长和种宽与最热月均气温呈显著正相关,而与最湿季降水量呈显著负相关[6];夏腊梅(Sinocalycanthuschinensis)的种子形态与光照条件的相关性最大[7];夜间增温导致冬小麦小穗数和每穗种子数均显著降低[8];水稻在生殖期间水分不足会导致产量降低[9-10]等。通过对比这些研究结果的不同趋势发现,植物种子繁殖对环境变化的适应性研究仍需要更多的实例以进一步探讨变化趋势的普遍性规律。
横断山区地处青藏高原东南缘,地势呈西北高东南低,海拔变化从1 000 m上升至5 000 m左右,较大的海拔变化导致该地区环境的区域差异和垂直变化十分明显[11-12]。例如,海拔高度每升高100 m,该地区的温度平均下降0.6 ℃[13];随着海拔高度的升高,降水呈先增多后减少的曲线变化趋势等[14]。不同海拔高度对应的诸多环境因子的变化,必然会对生长在该地区的植物种子繁殖产生影响[15]。高山地区湿地生态系统对环境变化极其敏感,由于其独有且重要的生态功能,这些湿地生态系统是近年来世界各国关注的焦点[16]。加强对这些湿地生态系统优势植物种子繁殖能力的研究,对了解高山地区湿地变化动态具有重要价值。
横断山区湿地生态系统与平原湿地不同,由于受地形条件限制,使其成为一个敏感和脆弱的地带[17]。湖滨带是该类型湿地生态结构的基础,发挥着缓冲带、生物多样性保育和护岸等生态功能[18-19]。湖滨带植物是维持湿地生态系统结构和功能的载体,种子繁殖是该地区湿地植物种群维持与扩散的重要方式[20]。研究其种子繁殖能力对环境变化的响应是掌握该地区湿地生态系统发展动态、科学评估环境变化对该区域湿地生态系统影响的基础。根据实地调查,水葱(Schoenoplectustabernaemontani)是横断山区很多湿地湖滨带的优势植物之一。滇西北著名的九大高原湖泊中,水葱都是其湖滨带优势植物[21]。过去关于环境变化对水葱生长的相关研究相对较多[15,22],但对其种子繁殖影响的研究还未见报道。因此,开展环境变化对高山地区湿地植物水葱种子繁殖性状影响的研究,是掌握湿地植被系统对环境变化响应与适应机制的关键。
本研究利用云南横断山区独特的山区地形以及气候条件,选取该地区典型的纳帕海湿地、拉市海湿地和滇池流域为采样地点,以其湖滨带优势植物水葱为研究对象,探讨水葱种子繁殖性状包括穗生长特征、种子产量及种子形态在不同采样地点的差异; 环境因子与这些种子繁殖性状的关系。通过研究,拟掌握高山地区湿地植物种子繁殖对环境变化的响应策略,为气候变化背景下科学评估高山地区湿地动态提供理论依据。
1 研究地概况
本研究选取环境差异较为明显的纳帕海湿地、拉市海湿地和滇池流域作为采样地点,这些湿地均为我国低纬度高海拔 、在空间上具有典型的“面山—湖滨—湖盆”生态结构特征的封闭与半封闭季节性湖泊湿地类型[23-24]。
纳帕海湿地位于云南香格里拉市境内,平均海拔3 274 m,面积共计3 100 hm2,属寒温带高原季风气候区,干湿季分明,年均降水量619.9 mm,其中,6—9月份为雨季,湖水上涨、水深可达4~5 m;10月份—次年5月份为旱季,平均水深不足20 cm,形成大面积的浅水沼泽和沼泽化草甸。纳帕海年均气温5.4 ℃,最热月为7月份,月均气温13.2 ℃,最冷月为1月份,月均气温-3.7 ℃[23]。
拉市海湿地位于丽江市玉龙县城西,平均海拔2 445 m,是长江上游典型的高原湿地[7]。最大水深7.5 m,平均水深4.5 m,常年水域面积为933.4 hm2。属于暖温带气候,干湿季分明,年均气温11.8 ℃,最热月平均气温18 ℃,最冷月平均气温3.9 ℃[25]。
滇池流域位于云南中部,平均海拔1 943 m,湖面面积330 km2,属于亚热带高原山地季风气候,干湿季分明。年温差为全国最小,年均气温15 ℃,年均降水量1 450 mm,无霜期在240 d以上[15]。
2 材料与方法
2.1 材料及性状测定
于2016年10月份植物生长末期,对3个采样地点不同环境条件下生长的水葱穗部进行采样(每个样地均不少于50株),装进封口袋统一编号并放进保鲜箱中迅速带回实验室进行分析。用精度为0.1 cm的直尺量取水葱的穗长,并记录每穗的小穗数,同时记录每穗种子产量。利用体视显微镜(德国徕卡,LEICA M165 C)对种子外观进行观察拍照并统一编号,用ImagJ图片处理软件(v.1.48;http://rsb.info.nih.gov/ij/)对种子照片进行测量,得到种子长和种子宽,并计算种子的长宽比。
2.2 研究地气候因子收集
从全球网格气候数据(精度:0.166×0.166; http://www.paleo.bris.ac.uk/)提取年均温、最暖月均温、最冷月均温、年平均降水量、生长季降水量、3个连续最湿月降水量、3个连续最干月降水量、年平均相对湿度、饱和大气压差等主要环境因子(表1)。
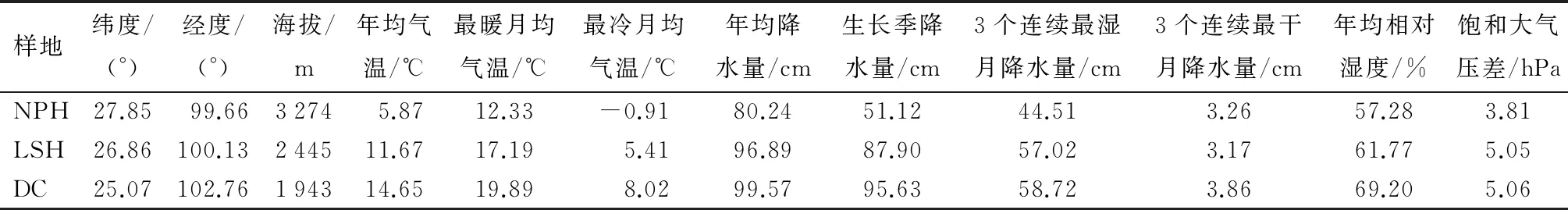
表1 不同地区环境因子数据
注:NPH.纳帕海湿地;LSH.拉市海湿地;DC.滇池流域。
2.3 数据分析
采用SPSS统计分析软件(v.19.0; https://www.ibm.com/analytics/us/en/technology/spss/),并应用单因素方差分析(one-way ANVOA)对不同采样地点水葱种子繁殖性状进行差异性检验,显著水平P<0.05。用R统计分析软件(v.3.01; https://www.r-project.org/)构建逐步回归分析模型 ,对影响水葱种子繁殖性状的主要环境因子(年均温、最暖月均温、最冷月均温、年平均降水量、生长季降水量、3个连续最湿月降水量、3个连续最干月降水量、年平均相对湿度、饱和大气压差)进行筛选,明确决定每个性状的主要环境因子以及性状与环境因子间的关系。
3 结果与分析
3.1 不同采样地点水葱穗特征的差异
水葱穗特征在纳帕海湿地、拉市海湿地、滇池流域3个采样地点间变化显著(表2),相比于纳帕海湿地,拉市海湿地和滇池流域的穗长和小穗数均显著增加(P<0.001,表2),穗长分别增加了40.92%、50.88%;小穗数分别增加了19.98%、39.20%。
3.2 不同采样地点水葱种子产量和种子形态的差异
水葱每穗种子产量和种子形态在纳帕海湿地、拉市海湿地、滇池流域3个采样地点间均存在显著差异,相比于纳帕海湿地,拉市海湿地和滇池流域的每穗种子产量、种长、种长/种宽均显著增加,每穗种子产量分别增加了51.37%、104.56%,种长分别增加了9.67%、24.7%,种长/种宽分别增加了11.26%、26.7%;而种宽无显著变化(表2)。

表2 不同采样地点水葱种子繁殖性状
注:表中数据为平均值±标准差;NPH.纳帕海湿地;LSH.拉市海湿地;DC.滇池流域;*** 表示P<0.001;同列不同字母表示差异显著(P<0.05);n=40。
3.3 环境因子和种子繁殖性状间的关系
水葱种子繁殖参数与年均温、最暖月均温、最冷月均温、年平均降水量、生长季降水量、3个连续最湿月降水量、3个连续最干月降水量、年平均相对湿度、饱和大气压差的逐步回归模型表明,水葱种子繁殖性状主要受年均温和最暖月均温两个环境因子的影响,并且这些种子繁殖性状与环境因子间均存在正相关关系,而与降水量和饱和大气压差间的相关性不显著 (表3)。其中,穗长主要受年均温和最暖月均温的共同影响,小穗数和种长主要受年均温的影响,每穗种子产量和种长/种宽主要受最暖月均温的影响,而种宽与本研究所有环境因子间的相关性均不显著(表3)。
表3基于逐步回归模型的水葱种子繁殖性状参数与气候因子的相关性
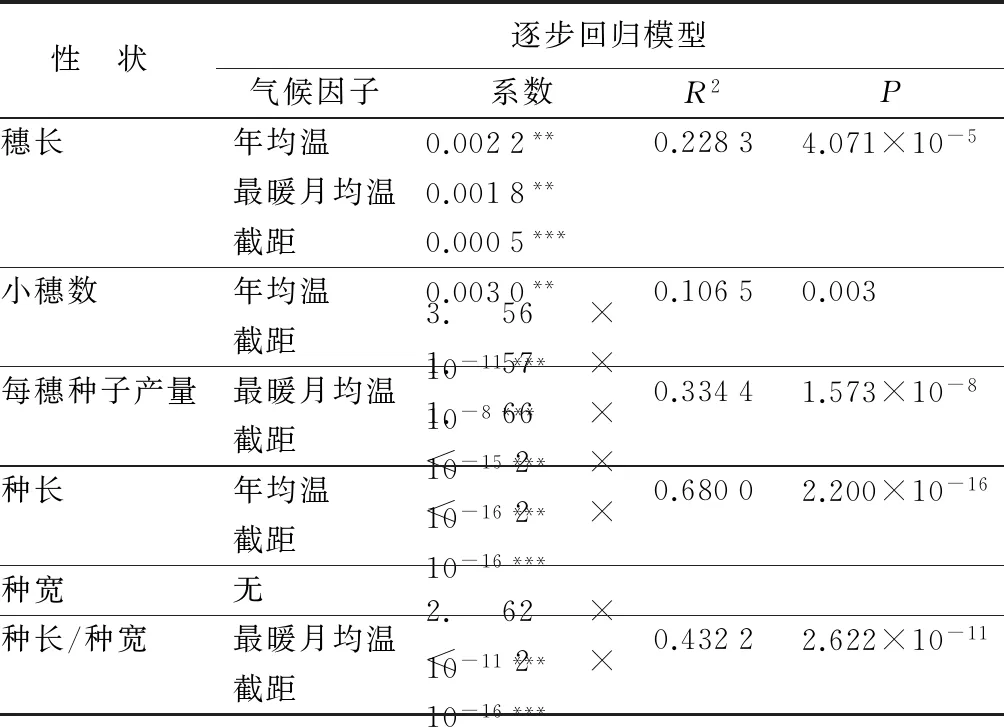
性 状逐步回归模型气候因子系数R2P穗长年均温0.0022**0.22834.071×10-5最暖月均温0.0018**截距0.0005***小穗数年均温0.0030**0.10650.003截距3.56×10-11***每穗种子产量最暖月均温1.57×10-8***0.33441.573×10-8截距1.66×10-15***种长年均温<2×10-16***0.68002.200×10-16截距<2×10-16***种宽无种长/种宽最暖月均温2.62×10-11***0.43222.622×10-11截距<2×10-16***
注:** 为P<0.01;*** 为P<0.001;n=40。
4 结论与讨论
探讨了湖滨带优势植物水葱种子繁殖性状在不同采样地点间的差异以及环境变化对种子繁殖的影响。结果表明,水葱种子繁殖性状在不同采样地点间差异显著,海拔高度降低引起的环境因子变化显著促进了水葱的种子繁殖能力,年均气温和最暖月均气温是影响水葱种子繁殖性状的主要因素,而降水和饱和大气压差对水葱种子繁殖性状的影响相对较小。
水葱种子繁殖性状在不同采样地点间的变化显著,海拔高度降低引起的环境变化致使水葱穗长、小穗数、每穗种子产量、种长、种长/种宽均显著升高,但对种宽没有显著影响 。过去针对该区域草本植物种子大小的研究也得到相似结论,如青藏高原东缘整个区系内草本植物的种子大小随着海拔高度的降低均呈显著增大趋势[26-27],表明种子繁殖能力与海拔高度的关系是一个普遍规律,随着海拔的升高,种子繁殖能力趋于降低。在高山地区,海拔升高,植物生长的环境条件更加苛刻,引起生长季缩短、光合能力下降,进而导致植物有机物积累降低,可能是导致植物对种子繁殖能力投资减少的重要原因[28]。这可能也是水葱种子繁殖能力在不同采样地点间的差异与这些采样地点环境因子变化密切相关的原因。
植物种子繁殖能力在不同采样地点间的差异主要由于不同采样地点间的生长环境不同,反映了植物对不同环境条件的适应策略[29]。本研究中,水葱的种子繁殖能力主要受温度条件变化的影响,与温度呈显著正相关,而受降水和饱和大气压差的影响较小 。过去一些相关的研究也得到类似的结果,例如,植物的种子产量、种子形态主要受温度等环境条件的影响[30-32];模拟增温提高了北美车前(Plantagovirginica)的光合同化能力,并增加了对植物繁殖投资的比例,促进了穗生长及植物种子繁殖能力[33];温度升高使得植物生长积温增加,生长期延长,叶片功能增强,光合作用增强,进而导致植物生殖产量的增加[9,34];随着温度升高,植物分蘖速度加快,导致种子实粒数增加,促进植物产量增加[35]等。种子繁殖性状与温度因子间呈显著正相关关系 ,可能是由于温度升高增加了植物的光合能力和生长速率,使植物的开花、结实等物候期提前,并延长植物的生长期,促进了植物的营养生长和碳积累,进而导致植物的生长和对植物各部分碳投资比例发生改变[36]。较高的碳积累,促使植物提高对各部分的投资比例,植物种子繁殖供应的能量也相应增加,使得植物的穗生长、种子产量、种子大小等反映植物种子繁殖能力的性状均呈增大趋势。
此外,研究结果表明,年均气温和最暖月均气温是影响横断山区湿地湖滨带优势植物的最主要温度因子。年均温反映全年的温度状况,一般来说,植物性状在发展过程中受基因控制,如果在植物性状的整个发展阶段均存在性状发生的相关基因表达,则该性状的发展持续时间较长,将受年均气温的显著影响。相反,如果性状的发展在植物生长期的短时间内迅速完成,则该性状可能受生长季温度或者与生长季密切相关的某些重要温度因子(如最暖月均温)影响较大。因此,推断穗形态(穗长和小穗数)以及种子长度的发展过程持续的时间较长,可能包含了植物的整个生长周期甚至更长。种子产量和种子长宽比的发展过程持续的时间较短,由于这两个性状主要受最暖月均温的影响,因此推测这两个性状的发展过程主要位于生长季中的最暖月,即在横断山区位于7月份。
[1] 祝振昌,张利权,肖德荣.上海崇明东滩互花米草种子产量及其萌发对温度的响应[J].生态学报,2011,31(6):1574-1581.
[2] 颜廷芬,丛沛桐,刘兴华,等.环境因子对植物生态位宽度影响程度分析[J].东北林业大学学报,1999,27(1):35-38.
[3] SHERRY R A, ZHOU X, GU S, et al. Divergence of reproductive phenology under climate warming[J]. Proceedings of the National Academy of Sciences of the United States of America,2007,104(1):198-202.
[4] CHUINE I, BEAUBIEN E G. Phenology is a major determinant of tree species range[J]. Ecology Letters,2010,4(5):500-510.
[5] 王国明,叶波.舟山群岛典型植物群落物种组成及多样性[J].生态学杂志,2017,36(2):349-358.
[6] 周旋,何正飚,康宏樟,等.温带-亚热带栓皮栎种子形态的变异及其与环境因子的关系[J].植物生态学报,2013,37(6):481-491.
[7] 张文标,金则新.夏蜡梅果实和种子形态变异及其与环境因子相关性[J].浙江大学学报(理学版),2007,34(6):689-695.
[8] 房世波,谭凯炎,任三学.夜间增温对冬小麦生长和产量影响的实验研究[J].中国农业科学,2010,43(15):3251-3258.
[9] 蒋小雪,金飚.气候变化对植物有性生殖影响的研究进展[J].西北植物学报,2012,32(10):2139-2150.
[10] 武立权.水分供应与稻米品质及产量性状关系的研究[D].合肥:安徽农业大学,2004.
[11] 刘振元,张杰,陈立.青藏高原植被指数最新变化特征及其与气候因子的关系[J].气候与环境研究,2017,22(3):289-300.
[12] SUN M, YANG S J, ZHANG J L, et al. Correlated evolution in traits influencing leaf water balance inDendrobium(Orchidaceae)[J]. Plant Ecology,2014,215(11):1255-1267.
[13] ZHANG S B, ZHOU Z K, HU H, et al. Photosynthetic performances ofQuercuspannosavary with altitude in the Hengduan Mountains, southwest China.[J]. Forest Ecology & Management,2005,212(1):291-301.
[14] 潘建峰,国庆喜,王化儒.山地小气候模型在帽儿山地区气候模拟中的应用[J].东北林业大学学报,2007,35(5):51-54.
[15] 刘国栋.气候变化对高原湿地优势植物生理生态学特性的影响研究[D].重庆:重庆大学,2016.
[16] GOPAL B. Future of wetlands in tropical and subtropical Asia, especially in the face of climate change[J]. Aquatic Sciences,2013,75(1):39-61.
[17] 董瑜,田昆,郭绪虎,等.不同区域气候条件影响下的纳帕海湿地植物叶绿素荧光特性[J].生态环境学报,2013,22(4):588-594.
[18] RICHARDSON D, HOLMES P, ESLER K, et al. Riparian vegetation: degradation, alien plant invasions, and restoration prospects[J]. Diversity & Distributions,2007,13(1):126-139.
[19] XIAO D, ZHANG C, TIAN K, et al. Development of alpine wetland vegetation and its effect on carbon sequestration after dam construction: A case study of Lashihai in the northwestern Yunnan plateau in China.[J]. Aquatic Botany,2015,126:16-24.
[20] 孟焕,王雪宏,佟守正,等.湿地土壤埋深对芦苇、香蒲种子萌发的影响[J].生态学杂志,2013,32(9):2320-2325.
[21] 李恒.横断山区的湖泊植被[J].植物分类与资源学报,1987,9(3):257-270.
[22] 董瑜,田昆,肖德荣,等.区域气候变化对纳帕海湖滨植物生理生化特性的影响[J].生态学报,2014,34(19):5570-5578.
[23] 肖德荣,田昆,张利权.滇西北高原纳帕海湿地植物多样性与土壤肥力的关系[J].生态学报,2008,28(7):3116-3124.
[24] 胡金明,李杰,袁寒,等.纳帕海湿地季节性景观格局动态变化及其驱动[J].地理研究,2010,29(5):899-908.
[25] 肖德荣,袁华,田昆,等.筑坝扩容下高原湿地拉市海植物群落分布格局及其变化[J].生态学报,2012,32(3):815-822.
[26] 郭淑青,齐威,王玉林,等.青藏高原东缘海拔对植物种子大小的影响[J].草业学报,2010,19(1):50-58.
[27] 杨霞,梁艳,陈学林.青藏高原东缘地区常见植物种子大小变异研究[J].生态科学,2007,26(6):483-489.
[28] MOLES A T, WARTON D I, STEVENS R D, et al. Does a latitudinal gradient in seedling survival favour larger seeds in the tropics?[J]. Ecology Letters,2004,7(10):911-914.
[29] STEPHANIE D C, MARGARETJ S, LAWREN S. Leaf trait diversification and design in seven rare taxa of the hawaiianPlantagoradiation[J]. International Journal of Plant Sciences,2009,170(1):61-75.
[30] 杨继.植物种内形态变异的机制及其研究方法[J].植物科学学报,1991,9(2):185-195.
[31] 张小彦.黄土丘陵沟壑区主要植物种子形态特征及有效性研究[D].杨凌:西北农林科技大学,2010.
[32] MOLES A, ACKERLY D, TWEDDLE J, et al. Global patterns in seed size[J]. Global Ecology & Biogeography,2007,16(1):109-116.
[33] 张万灵,肖宜安,闫小红,等.模拟增温对入侵植物北美车前生长及繁殖投资的影响[J].生态学杂志,2013,32(11):2959-2965.
[34] JIN B, WANG L, WANG J, et al. The effect of experimental warming on leaf functional traits, leaf structure and leaf biochemistry inArabidopsisthaliana[J]. Bmc Plant Biology,2011,11(1):1-10.
[35] 池再香,杨桂兰,杨黎,等.不同海拔高度的光温因子对超级稻陆两优106产量的影响研究[J].贵州气象,2007,31(6):9-10.
[36] 刘振亚,张晓宁,李丽萍,等.模拟增温对滇西北高原典型湿地湖滨带优势植物的光和CO2利用能力的影响[J].生态学报,2017,37(23):7821-7832.
