不同氮敏感性粳稻品种的氮代谢与光合特性比较
2018-03-13剧成欣周著彪赵步洪王志琴杨建昌
剧成欣 周著彪 赵步洪 王志琴 杨建昌,*
不同氮敏感性粳稻品种的氮代谢与光合特性比较
剧成欣1,2周著彪1赵步洪2王志琴1杨建昌1,*
1扬州大学江苏省作物遗传生理国家重点实验室培育点 / 粮食作物现代产业技术协同创新中心, 江苏扬州 225009;2江苏里下河地区农业科学研究所, 江苏扬州 225007
本文旨在探明不同氮敏感性粳稻品种氮代谢与光合特性。以2个氮敏感高产品种淮稻5号和连粳7号, 2个氮钝感品种宁粳1号和扬粳4038为材料, 设置0和200 kg hm–22个施氮水平, 研究其产量、氮肥利用效率以及地上部生理性状的变化特点。结果表明, 在2种施氮水平下, 氮敏感品种的产量和氮肥利用效率显著高于氮钝感品种。与氮钝感品种相比, 氮敏感品种具有较高的光合速率和氮素积累, 较强的氮代谢酶活性和较高的光合氮素利用效率, 抽穗期茎叶中积累较多的可溶性糖和淀粉, 抽穗至成熟期茎鞘中非结构性碳水化合物向籽粒转运率较高。表明氮敏感品种在较低施氮量下具有较高的生理活性和物质生产效率; 这些特征可作为筛选高产氮敏感水稻品种的重要生理指标。
水稻; 氮敏感性; 产量; 氮肥利用效率; 生理性状
随着气候变暖和生态安全压力的增加, 水稻等作物的高产高效特别是氮素的高效利用研究已成为当前农业领域的研究热点[1-2]。不同水稻品种的氮肥利用效率存在明显的差异, 培育和选用氮敏感品种或氮高效品种是协同提高作物产量和氮肥利用效率的一条重要途径[3-4]。在以往的研究中我们发现[5], 现代品种在施氮量为300 kg hm–2条件下, 不同品种间的产量差异并不显著; 但施氮量在≤200 kg hm–2条件下, 不同品种对氮素的响应表现出显著的差异, 并将在施氮量≤200 kg hm–2下产量和氮肥利用效率较高的品种称为氮敏感品种; 将在高施氮量(>200 kg hm–2)下产量较高的品种称为氮钝感品种[5]。但对于这两类品种的农艺生理特征缺乏深入研究。
作物光合生产能力以及同化物向经济器官的运转能力是作物产量形成的2个关键因素[6-8]。这些植株生理性状不仅影响水稻的光合生产能力以及营养物质分配, 与氮素的吸收利用也存在着密切的关系[9-12]。叶片作为光合作用的主要器官直接影响水稻产量形成和氮素吸收与运转[13-14]。但不同氮敏感性水稻品种植株的生理性状有哪些差异, 氮敏感品种有哪些生理特征, 缺乏深入的研究。本试验以不同氮敏感性水稻品种为材料, 研究了水稻叶片光合速率和氮代谢酶活性的变化, 以及生育后期植株非结构性碳水化合物的运转等地上部生理性状, 以期探明氮敏感品种地上部生理性状特征, 为氮敏感品种和氮高效水稻品种的培育和选用提供依据。
1 材料与方法
1.1 材料与试验设计
2015年和2016年在江苏省里下河地区农业科学研究所夏桥基地, 在前期试验的基础上, 选用2个氮敏感品种淮稻5号和连粳7号, 2个氮钝感品种宁粳1号和扬粳4038[5]。采用裂区设计, 氮肥处理为主区, 品种为裂区(小区), 小区面积为30 m2, 重复3次。设置全生育期施纯氮0和200 kg hm–22个氮肥水平处理, 依据含氮率折算成尿素(含纯氮46%)施用, 其中基肥、分蘖肥、促花肥和保花肥各占40%、20%、20%和20%。5月12日播种, 6月14日移栽, 株行距25 cm×15 cm, 双本栽插, 主区间筑埂并用塑料薄膜包裹。移栽前在各小区施用过磷酸钙(含P2O513%, 纯磷28%) 300 kg hm–2, 氯化钾(含K2O 63%, 纯钾71%) 195 kg hm–2, 分基肥和拔节肥2次施用, 各占60%和40%。除中期搁田外, 全生育期保持浅水层直到收获前1周断水, 严格控制杂草和病虫害。
1.2 叶片光合速率测定
使用美国LI-COR 6400便携式光合测定仪于幼穗分化期测定植株最上一片完全展开叶, 于穗分化始期、抽穗期和抽穗后10 d、20 d和30 d测定剑叶的光合速率。使用红蓝光源, 光量子通量密度(PFD)为1400 μmol m–2s–1, 叶室CO2浓度为380 μmol mol–1, 温度28~30°C, 用3台光合测定仪同时测定, 每处理每次重复测定6片叶。
1.3 叶片氮代谢酶活性和植株含氮量的测定
于穗分化始期和抽穗期, 取每处理10张叶片(幼穗分化期取最上一片完全展开叶, 抽穗期取剑叶), 分别测定叶片中谷氨酰胺合成酶(GS)、谷氨酸合酶(GOGAT)和硝酸还原酶(NR)活性, 以牛血清白蛋白(BSA)为标准测定酶粗液中蛋白质含量[15-18]。于成熟期从各小区取代表性植株5穴, 由分蘖节处分为地上部和地下部2个部分, 将地上部分解为绿叶、茎鞘、穗, 杀青后烘干用于测定干物质重, 保留样本, 粉碎过筛后用凯氏定氮法测定含氮量。氮肥农学利用率(kg kg–1) = (施氮区籽粒产量-氮空白区籽粒产量)/施氮量; 氮肥吸收利用率(%) = (施氮区植株吸氮量-氮空白区植株吸氮量)/施氮量×100; 氮肥生理利用率(kg kg–1) = (施氮区籽粒产量-氮空白区籽粒产量)/(施氮区植株吸氮量-氮空白区植株吸氮量); 氮肥偏生产力(kg kg–1) = 籽粒产量/施氮量。比叶氮含量= 叶片含氮量/比叶面积, 光合氮素利用效率= 叶片光合速率/比叶氮含量。
1.4 非结构性碳水化合物测定
于抽穗期和成熟期取各处理12穴稻株, 依据Yoshida等[19]方法测定茎鞘中非结构性碳水化合物。依据Somogyi[20]的方法蔗糖测定浓度。依据Pucher等[21]的方法分析可溶性糖和淀粉, 非结构性碳水化合物运转率(%) = (抽穗期积累量-成熟期积累量)/抽穗期积累量×100。
1.5 考种计产
成熟期取各小区1 m2考察实际穗数, 取12穴水稻刮粒后测定穗粒数、结实率与千粒重, 采用水漂法测定结实率, 将稻粒置密度为1.1×103kg m–3的生理盐水中, 沉入水底者是饱粒, 其余为空瘪粒。从每个小区实收5 m2稻株脱粒计产。
1.6 数据分析
用Microsoft Excel软件整理数据, SAS软件统计分析数据, SigmaPlot 10.0绘图。由于2年试验结果趋势一致, 且同一年代品种各测定指标在年度间的差异不显著(<1)。因此, 本文除产量性状和氮肥利用效率数据列出2年数据外, 其余生理测定数据用2016年试验结果表示。
2 结果与分析
2.1 产量与氮肥利用效率
如表1所示, 在同一施氮量处理下, 2个氮敏感品种的产量显著高于2个氮钝感水稻品种, 但在2个氮敏感品种间或在2个氮钝感品种间的产量差异不显著, 两年的结果趋势一致(表1)。从产量构成因素看, 氮敏感品种产量较高的原因是穗数和穗粒数的乘积较高, 穗数和穗粒数的协调增长使得总颖花量较大, 保证库容量, 这是氮敏感品种产量较高的重要原因。与产量的变化趋势类似, 在2个施氮量处理下, 氮敏感品种的氮肥吸收利用效率、农学利用效率、生理利用效率和偏生产力显著高于氮钝感品种(表2)。
2.2 叶片光合速率
由图1可知, 在不施氮量处理下, 叶片光合速率在2个氮敏感品种间或2个氮钝感品种间差异不显著, 但2个氮敏感品种的叶片光合速率显著高于2个氮钝感品种(图1-A)。在施氮量为200 kg hm–2时, 不同氮敏感性水稻品种间的差异趋势与不施氮处理下表现一致(图1-B)。
2.3 氮素积累量
氮敏感品种的氮素积累量在分蘖中期与氮钝感品种相比没有显著差异, 但是在抽穗期和成熟期, 氮敏感品种的氮素积累量显著高于氮钝感品种。2个氮敏感品种间或2个氮钝感品种间的氮素积累量差异不显著(图2)。
氮素阶段积累量表现为自移栽至穗分化始期, 氮敏感品种与氮钝感品种间没有显著差异, 但是自穗分化始期至抽穗期和自抽穗期至成熟期, 氮敏感水稻品种显著高于氮钝感品种(图3)。
2.4 氮代谢酶活性
化始期和抽穗期剑叶中的氮代谢关键酶活性在不同氮敏感性水稻品种间有显著差异(表3)。在不同生育时期和不同施氮量下, 氮敏感品种剑叶中硝酸还原酶活性均显著高于氮钝感品种。氮敏感品种的谷氨酰胺合成酶和谷氨酸合酶活性在不施氮处理下, 2个生育时期均显著高于氮钝感品种; 但是在施氮量为200 kg hm–2时, 仅在穗分化始期氮敏感品种显著高于氮钝感品种, 在抽穗期, 谷氨酰胺合成酶和谷氨酸合酶活性在不同氮敏感性水稻品种间差异不显著(表3)。

表1 不同水稻品种产量及其构成因素
HD-5: 淮稻5号; LJ-7: 连粳7号; NJ-1: 宁粳1号; YJ-4: 扬粳4038。同栏同年同一施氮量下比较, 标以不同字母的值在=0.05水平上差异显著。
HD-5: Huaidao 5; LJ-7: Lianjing 7; NJ-1: Ningjing 1; YJ-4: Yangjing 4038. Values within the same column under same year and same nitrogen rate followed by a different letter are significantly different at the 0.05 probability level.

表2 不同水稻品种氮肥利用效率
RE: 吸收利用效率; AE: 农学利用效率; PE: 生理利用效率; PFP: 氮肥偏生产力。HD-5: 淮稻5号; LJ-7: 连粳7号; NJ-1: 宁粳1号; YJ-4: 扬粳4038。同栏同年比较, 标以不同字母的值在=0.05水平上差异显著。
RE: recovery nitrogen use efficiency; AE: agronomic nitrogen use efficiency; PE: physiological nitrogen use efficiency; PFP: partial factory productivity. HD-5: Huaidao 5; LJ-7: Lianjing 7; NJ-1: Ningjing 1; YJ-4: Yangjing 4038. Values within the same column under same year followed by a different letter are significantly different at the 0.05 probability level.
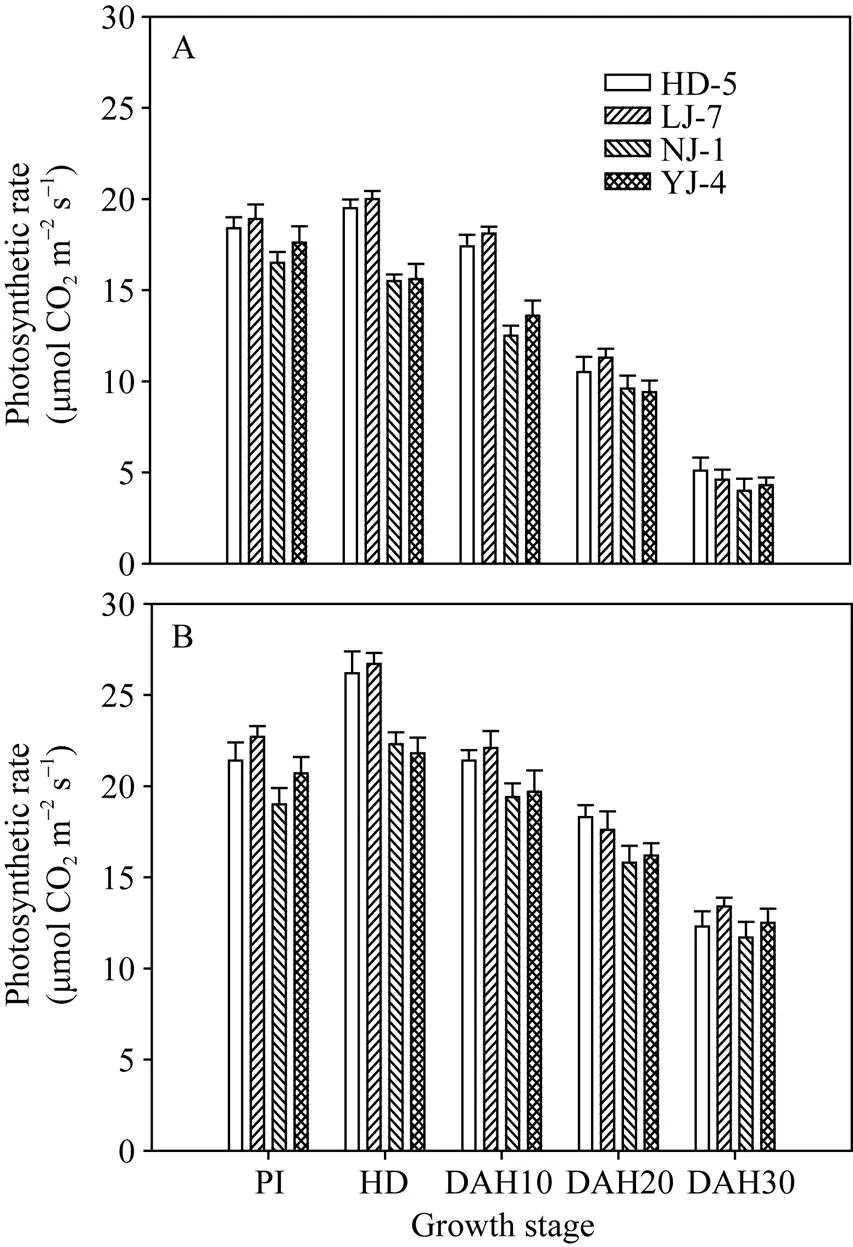
图1 不同水稻品种在0N(A)和200 kg N hm–2(B)处理下的叶片光合速率
PI: 穗分化始期; HD: 抽穗期; DAH10: 抽穗后10 d; DAH20: 抽穗后20 d; DAH30: 抽穗后30 d; HD-5: 淮稻5号; LJ-7: 连粳7号; NJ-1: 宁粳1号; YJ-4: 扬粳4038。
PI: panicle initiation; HD: heading; DAH10: 10 days after heading; DAH20: 20 days after heading; DAH30: 30 days after heading; HD-5: Huaidao 5; LJ-7: Lianjing 7; NJ-1: Ningjing 1; YJ-4: Yangjing 4038.
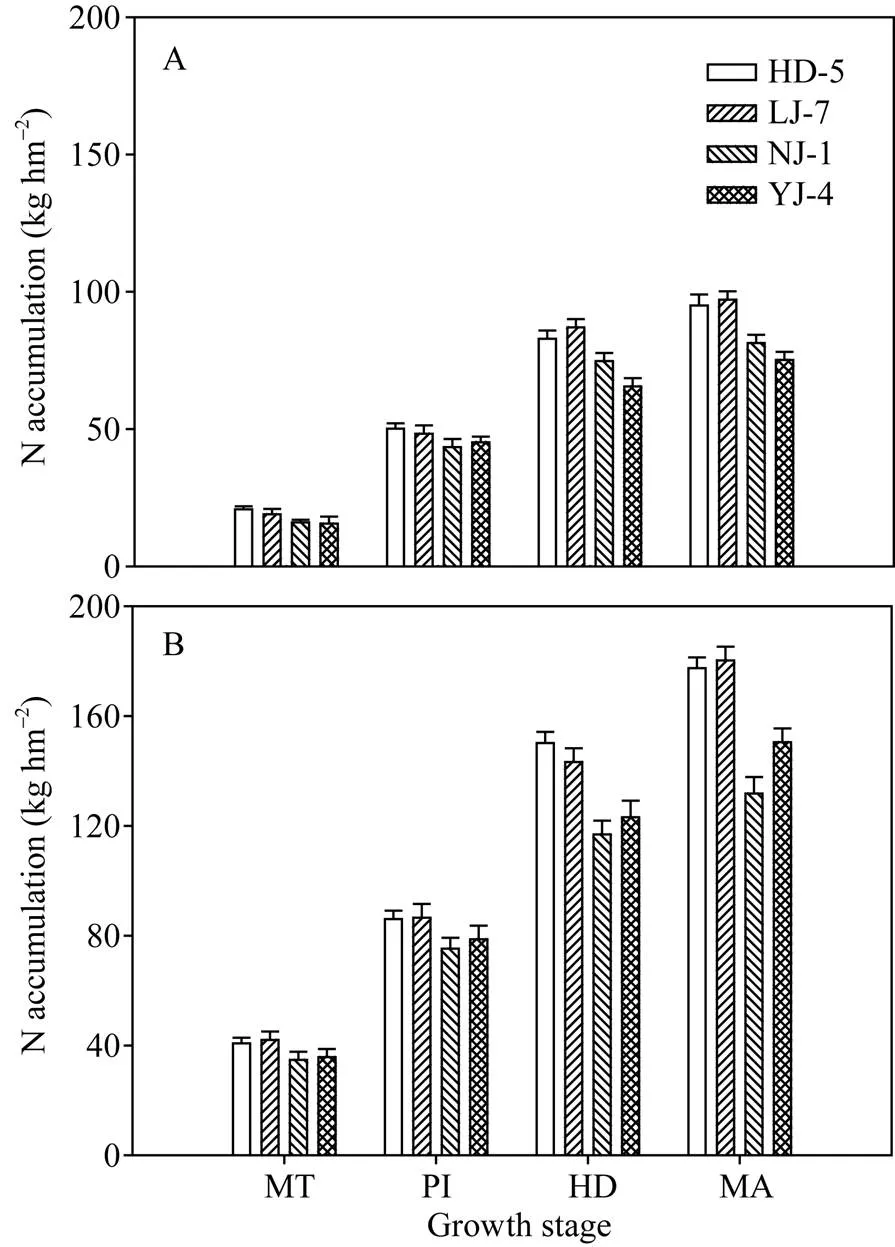
图2 不同水稻品种在0N (A)和200 kg N hm–2(B)处理下的氮素积累量
MT: 分蘖中期; PI: 穗分化始期; HD: 抽穗期; MA: 成熟期; HD-5: 淮稻5号; LJ-7: 连粳7号; NJ-1: 宁粳1号; YJ-4: 扬粳4038。
MT: middle tillering; PI: panicle initiation; HD: heading; MA: maturity; HD-5: Huaidao 5; LJ-7: Lianjing 7; NJ-1: Ningjing 1; YJ-4: Yangjing 4038.

图3 不同水稻品种在0N(A)和200 kg N hm–2(B)处理下的氮素阶段积累量
TS: 移栽期; MT: 分蘖中期; PI: 穗分化始期; HD: 抽穗期; MA: 成熟期; HD-5: 淮稻5号; LJ-7: 连粳7号; NJ-1: 宁粳1号; YJ-4: 扬粳4038。
TS: transplanting; MT: middle tillering; PI: panicle initiation; HD: heading; MA: maturity; HD-5: Huaidao 5; LJ-7: Lianjing 7; NJ-1: Ningjing 1; YJ-4: Yangjing 4038.
2.5 光合氮素利用效率
不同氮敏感性水稻品种的叶片含氮量差异显著,但在氮敏感品种和氮钝感品种间并无明显的变化规律。氮敏感品种的比叶氮含量显著高于氮钝感品种。光合氮素利用效率是叶片光合速率与比叶氮的比值, 表示单位氮素的光合生产能力。氮敏感品种的光合氮素利用效率在2个施氮量处理下均显著高于氮钝感品种(表4)。2个氮钝感品种的光合氮素利用效率在施氮量为200 kg hm–2时差异不显著, 但在不施氮处理下表现出显著差异, 其原因可能归于不同品种叶片氮含量的差异。
2.6 非结构性碳水化合物
性糖包括蔗糖和可溶性单糖, 非结构性碳水化合物的总量即可溶性糖与淀粉的含量相加。在抽穗期和2个施氮量处理下的可溶性糖、蔗糖和淀粉含量, 氮敏感水稻品种均显著高于氮钝感品种(表5)。在成熟期, 除了淀粉的含量差异显著以外, 可溶性总糖和蔗糖含量在不同氮敏感性水稻品种间无显著差异。在抽穗期, 氮敏感品种较氮钝感品种在茎鞘中积累更多的非结构性碳水化合物。在成熟期, 不同氮敏感性水稻品种间同化物积累差异不显著。氮敏感品种非结构性碳水化合物转运率显著高于氮钝感品种(表6), 表明氮敏感品种的物质生产效率较高。

表3 不同水稻品种氮代谢关键酶活性
NR: 硝酸还原酶; GS: 谷氨酰胺合成酶; GOGAT: 谷氨酸合酶; PI: 穗分化始期; HD: 抽穗期。HD-5: 淮稻5号; LJ-7: 连粳7号; NJ-1: 宁粳1号; YJ-4: 扬粳4038。同栏同一施氮量下比较, 标以不同字母的值在=0.05水平上差异显著。
NR: nitrate reductase; GS: glutamine synthetase; GOGAT: Glutamate synthase; PI: panicle initiation; HD: heading. HD-5: Huaidao 5; LJ-7: Lianjing 7; NJ-1: Ningjing 1; YJ-4: Yangjing 4038. Values within the same column and same nitrogen rate followed by a different letter are significantly different at the 0.05 probability level.

表4 不同水稻品种的叶片氮含量、比叶氮含量和光合氮素利用效率
LNC: 叶片含氮量; SLNC: 比叶氮含量; PUNE: 光合氮素利用效率。HD-5: 淮稻5号; LJ-7: 连粳7号; NJ-1: 宁粳1号; YJ-4: 扬粳4038。同栏同一施氮量下比较, 标以不同字母的值在=0.05水平上差异显著。
LNC: leaf nitrogen content; SLNC: specific leaf nitrogen content; PUNE: photosynthetic nitrogen use efficiency. HD-5: Huaidao 5; LJ-7: Lianjing 7; NJ-1: Ningjing 1; YJ-4: Yangjing 4038. Values within the same column and same nitrogen rate followed by a different letter are significantly different at the 0.05 probability level.
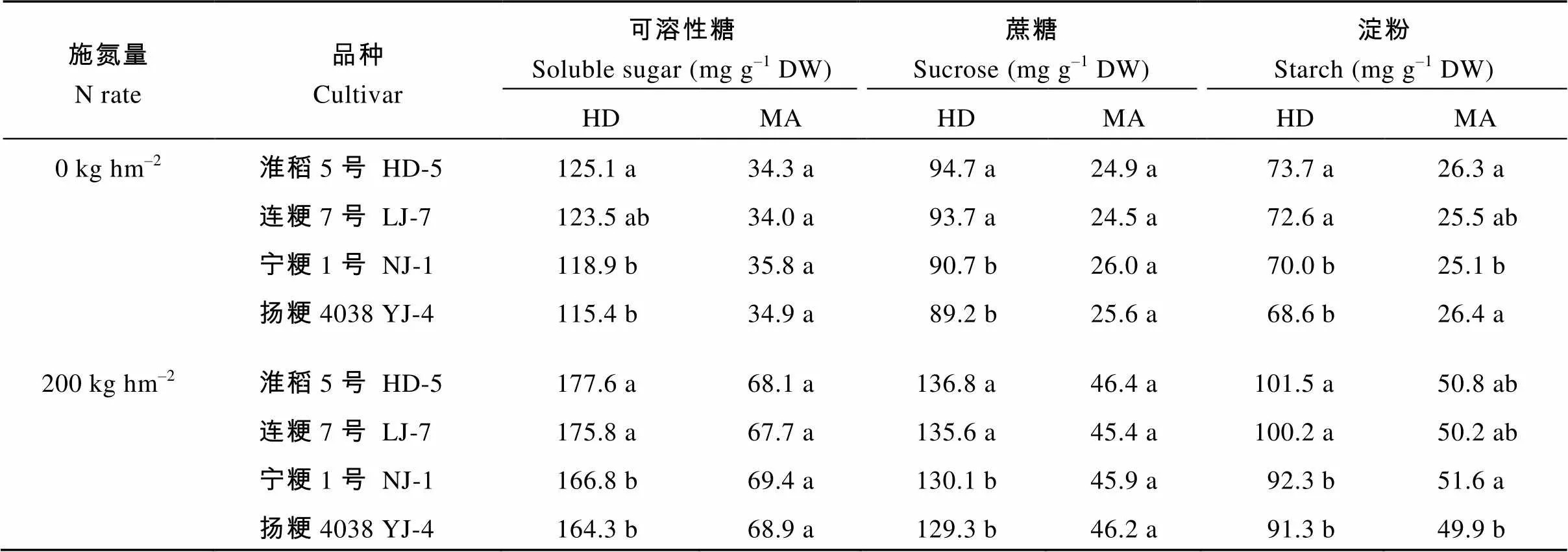
表5 不同水稻品种茎鞘中可溶性糖、蔗糖和淀粉含量
HD-5: 淮稻5号; LJ-7: 连粳7号; NJ-1: 宁粳1号; YJ-4: 扬粳4038。同栏同一施氮量下比较, 标以不同字母的值在=0.05水平上差异显著。
HD-5: Huaidao 5; LJ-7: Lianjing 7; NJ-1: Ningjing 1; YJ-4: Yangjing 4038. Values within the same column and same nitrogen rate followed by a different letter are significantly different at the 0.05 probability level.
3 讨论
水稻氮高效主要体现在2个方面, 即高效吸收和高效利用, 前者是前提, 后者是宗旨[11-12]。评价氮肥效率的指标众多[12,15]。本研究表明, 氮敏感性水稻品种的产量和氮肥利用效率在2个施氮水平下均表现出明显的品种间差异, 氮敏感品种的氮肥吸收利用效率、农学利用效率、生理利用效率和偏生产力均显著高于氮钝感品种。表明选用氮敏感性品种可以获得高产与氮肥高效利用的效果。
光合作用是植物生物产量和经济产量的基础, 作物产量的90%~95%直接或间接来自光合作用[22-23]。本研究表明, 与氮钝感品种相比, 氮敏感品种在整个生育期具有较高的叶片光合速率。在籽粒灌浆期, 较高的叶片光合速率, 可以促进籽粒灌浆。特别需要指出的是, 氮敏感品种较氮钝感品种有较高的氮素光合利用效率。光合氮素利用效率是叶片光合速率与比叶氮含量的比值。氮素光合利用效率较高, 表明产生单位干物质所需的氮素较少, 这是氮敏感品种氮素高效利用的一个重要生理机制。
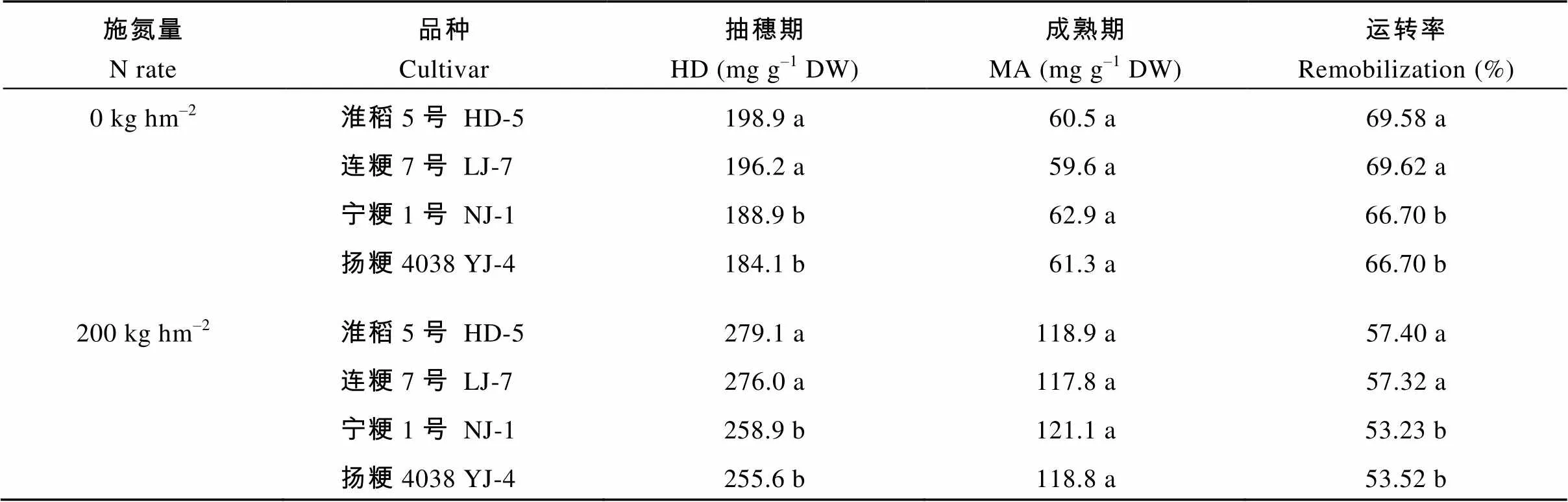
表6 不同水稻品种非结构性碳水化合物的运转
HD-5: 淮稻5号; LJ-7: 连粳7号; NJ-1: 宁粳1号; YJ-4: 扬粳4038。同栏同一施氮量下比较, 标以不同字母的值在=0.05水平上差异显著。
HD-5: Huaidao 5; LJ-7: Lianjing 7; NJ-1: Ningjing 1; YJ-4: Yangjing 4038. Values within the same column and same nitrogen rate followed by a different letter are significantly different at the 0.05 probability level.
有研究认为, 地上部与根系存在着相互作用[24], 地上部生长发育良好可以保证充足的营养物质向根部输送, 从而保持活跃的根系功能; 活跃的根系又反过来为地上部提供充分的水分和植物激素, 进而改善地上部的生长, 促进植株生物产量的提高[25-26]。因此, 叶片光合性状的提高也为地下部根系生长提供了充足的碳源。氮敏感品种具有较高的叶片光合速率, 特别是较高的氮素光合利用效率, 这是其获得高产和养分高效利用的重要生理原因。
有研究表明, 不同氮敏感性水稻品种在成熟期的氮素积累量具有很大差异, 但关于不同水稻品种在全生育期中的氮素积累动态变化及其不同生育阶段的氮素积累的研究较少[27-29]。本研究观察到, 在分蘖中期和穗分化始期, 氮敏感品种的氮素积累量与氮钝感品种相比没有明显的优势, 但在抽穗期和成熟期, 氮敏感品种的氮素积累量显著高于氮钝感品种。通过分析不同氮敏感性水稻品种的氮素阶段积累特性发现, 造成这一差异的主要原因在于, 自穗分化始期至成熟期氮敏感品种的氮素积累量显著高于氮钝感品种。自穗分化始期至成熟期氮素积累量高, 有利于水稻的颖花分化和发育, 有利于叶片光合作用和提高库的生理活性(细胞分裂素等激素含量, 蔗糖-淀粉代谢途径关键酶活性等), 有利于籽粒灌浆充实[5, 30-31]。
水稻对氮素的吸收与利用必须经一系列氮代谢酶参与的反应和转化来完成。硝酸还原酶是植物器官中硝态氮还原同化过程中第一个酶和限速酶, 谷氨酰胺合成酶-谷氨酸合酶循环是植物体内氨同化的主要途径, 是整个氮代谢的中心, 上述酶类对水稻氮素吸收有重要的调控作用[32-34]。本研究表明, 不同氮敏感性水稻品种的氮代谢相关酶活性存在显著差异, 氮敏感品种的氮代谢相关酶活性在穗分化始期和抽穗期显著高于氮钝感品种, 说明氮代谢酶活性的增强是氮敏感水稻品种氮肥利用效率较高一个重要生理机制。但当氮肥过高时会对结实期水稻植株中蛋白质的降解及运转造成不利影响[13], 各氮代谢酶活性对不同施氮水平的响应是否存在一定的协同关系还有待进一步研究。
水稻的氮代谢与糖代谢间具有十分密切的联系[35]。本研究表明, 与氮钝感品种相比, 氮敏感品种在抽穗期茎鞘中能够贮存更多的非结构性碳水化合物, 且灌浆期碳水化合物的运转率显著高于氮钝感品种。抽穗期茎鞘中贮存的非结构性碳水化合物多, 有利于形成大穗, 有利于籽粒灌浆早期胚乳发育, 增大库强; 抽穗后茎鞘中贮存的非结构性碳水化合物向籽粒转运率高, 可以促进籽粒灌浆, 提高结实率和粒重[4,35]。我们在以往的研究中还观察到, 促进茎中碳同化物向籽粒转运, 同时也可以促进营养器官的氮素向籽粒转运, 进而提高氮收获指数和氮肥利用效率[5,13,23]。此外, 地上部糖代谢还与地下部的生长发育关系密切, 地上部提供充足的碳水化合物运输到地下部才能维持地下部的根系活性[36-37]。因此, 提高抽穗期水稻茎鞘中非结构性碳水化合物的积累, 在灌浆期促进其向籽粒转运是促进水稻籽粒灌浆, 进而提高产量和氮肥利用效率的重要途径。
4 结论
在施氮量≤200 kg hm–2条件下, 氮敏感水稻品种的产量和氮肥利用效率均显著高于氮钝感品种。与氮钝感品种相比, 氮敏感品种具有较高的光合速率和氮素积累, 较强的氮代谢酶活性和较高的光合氮素利用效率, 抽穗期茎鞘中积累较多的可溶性糖和淀粉, 籽粒灌浆期茎鞘中非结构性碳水化合物向籽粒转运率较高。这些表现是氮敏感品种产量和氮肥利用效率高的重要生理原因, 上述生理性状可作为筛选氮敏感水稻品种的重要指标。
[1] FAOSTAT. FAO Statistical Databases, Food and Agriculture Organization (FAO) of the United Nations, Rome, 2016. http:// www.fao.org/
[2] Normile D. Reinventing rice to feed the world., 2008, 321: 330–333
[3] Kant S, Bi Y M, Rothstein S J. Understanding plant response to nitrogen limitation for the improvement of crop nitrogen use efficiency., 2011, 62: 1490–1509
[4] Haegele J W, Cook K A, Nichols D M, Below F E. Changes in nitrogen use traits associated with genetic improvement for grain yield of maize hybrids released in different decades., 2013, 53: 1256–1268
[5] Ju C X, Roland J B, Wang Z Q, Zhang H, Liu L J, Yang J C, Zhang J H. Root and shoot traits for rice varieties with higher grain yield and higher nitrogen use efficiency at lower nitrogen rates application., 2015, 175: 47–55
[6] 黄元财, 王术, 吴晓冬, 王祥九, 刘向英, 王德伟. 肥水条件对不同类型水稻干物质积累与分配的影响. 沈阳农业大学学报, 2004, 35: 346–349 Huang Y C, Wang S, Wu X D, Wang X J, Liu X Y, Wang D W. Effect of fertilizer and water conditions on dry matter accumulation and partition of different types of rice., 2004, 35: 346–349 (in Chinese with English abstract)
[7] 霍中洋, 姚义, 张洪程, 夏炎, 倪晓诚, 戴其根, 许轲, 魏海燕. 播期对直播稻光合物质生产特征的影响. 中国农业科学, 2012, 45: 2592–2606 Huo Z Y, Yao Y, Zhang H C, Xia Y, Ni X C, Dai Q G, Xu K, Wei H Y. Effect of sowing date on characteristics of photosynthesis and matter production of direct seeding rice., 2012, 45: 2592–2606 (in Chinese with English abstract)
[8] 董桂春, 王熠, 于小凤, 周娟, 彭斌, 李进前, 田昊, 张燕, 袁秋梅, 王余龙. 不同生育期水稻品种氮素吸收利用的差异. 中国农业科学, 2011, 44: 4570–4582 Dong G C, Wang Y, Yu X F, Zhou J, Peng B, Li J Q, Tian H, Zhang Y, Yuan Q M, Wang Y L. Differences of nitrogen uptake and utilization of conventional rice varieties with different growth duration., 2011, 44: 4570–4582 (in Chinese with English abstract)
[9] 张岳芳, 王余龙, 张传胜, 董桂春, 杨连新, 黄建晔, 陈培峰, 龚克成.籼稻品种的氮素累积量与根系性状的关系. 作物学报, 2006, 32: 1121–1129 Zhang Y F, Wang Y L, Zhang C S, Dong G C, Yang L X, Huang J Y, Chen P F, Gong K C. Relationship between N accumulation and root traits in conventionalrice varieties (L.)., 2006, 32: 1121–1129 (in Chinese with English abstract)
[10] 于小凤, 王坚纲, 李进前, 王熠, 袁秋梅, 仲军, 陈琛, 田昊, 张燕, 黄建晔, 王余龙, 董桂春. 不同氮素吸收类型梗稻品种吸氮能力的差异及原因分析. 中国水稻科学, 2012, 26: 331–340 Yu X F, Wang J G, Li J Q, Wang Y, Yuan Q M, Zhong J, Chen C, Tian H, Zhang Y, Huang J Y, Wang Y L, Dong G C. Difference in nitrogen absorption ability and its reason in conventionalrice cultivars with different nitrogen uptake efficiencies., 2012, 26: 331–340 (in Chinese with English abstract)
[11] 熊洁.水稻源库结构、稻米品质以及对氮响应的基因型差异.南京农业大学博士学位论文, 江苏南京, 2011 Xiong J. The Genetic Differences in Source-sink Structure, Rice Quality and Responses to Nitrogen. PhD Dissertation of Nanjing Agricultural University. Nanjing, China, 2011
[12] 陈琛. 氮高效吸收高产类型遗传群体水稻株系的基本特点. 扬州大学博士学位论文, 江苏扬州, 2016 Chen C. Basic Characteristics of Nitrogen Efficient Absorption and High Yield Rice in Genetic Populations. PhD Dissertation of Yangzhou University. Yangzhou, China, 2016
[13] 剧成欣, 陶进, 钱夕旸, 顾骏飞, 张耗, 赵步洪, 王志琴, 杨建昌. 不同年代中籼水稻品种的叶片光合性状. 作物学报, 2016, 42: 415–426 Ju C X, Tao J, Qian X Y, Gu J F, Zhang H, Zhao B H, Wang Z Q, Yang J C. Leaf photosynthetic characteristics of mid-seasonrice varieties applied at different decades., 2016, 42: 415–426 (in Chinese with English abstract)
[14] Gu J F, Yin X Y, Paul C S, Tjeerd J S, Wang H Q. Using chromosome introgression lines to map quantitative trait loci for photosynthesis parameters in rice (L.) leaves under drought and well-watered field conditions., 2012, 63: 455–469
[15] 陈露, 张伟杨, 王志琴, 张耗, 刘立军, 杨建昌. 施氮量对江苏不同年代中粳稻品种产量与群体质量的影响. 作物学报, 2014, 40: 1412−1423 Chen L, Zhang W Y, Wang Z Q, Zhang H, Liu L J, Yang J C. Effect of nitrogen application rate on grain yield and population quality of mid-seasonrice cultivars at different decades in Jiangsu province., 2014, 40: 1412−1423 (in Chinese with English abstract)
[16] 赵全志, 陈静蕊, 刘辉, 乔江方, 高桐梅, 杨海霞, 王继红. 水稻氮素同化关键酶活性与叶色变化的关系. 中国农业科学, 2008, 41: 2607−2616 Zhao Q Z, Chen J R, Liu H, Qiao J F, Gao T M, Yang H X, Wang J H. Relationship between activities of nitrogen assimilation enzymes and leaf color of rice., 2008, 41: 2607−2616. (in Chinese with English abstract)
[17] 李合生. 植物生理生化实验原理和技术. 北京: 高等教育出版社, 2000. pp 125−127 Li H S. Principle and Technology of Plant Physiological and Biochemical Experiments. Beijing: Higher Education Press, 2000. pp 125−127 (in Chinese)
[18] 陈露. 江苏不同年代中粳稻品种对施氮量的响应及其生理机制. 扬州大学硕士学位论文, 江苏扬州, 2014 Chen L. Response of the Mid-seasonRice Cultivars Applied at Different Decades in Jiangsu to Nitrogen Rates and Its Physiological Mechanism. MS Thesis of Yangzhou University. Yangzhou, Jiangsu, China, 2014
[19] Yoshida S, Forno D, Cock J, Gomez K. Determination of sugar and starch in plant tissue. In: Yoshida S. ed. Laboratory Manual for Physiological Studies of Rice. The International Rice Research Institute, Philippines, 1976. pp: 46–49
[20] Somogyi M. A new reagent for the determination of sugars., 1945, 160: 61–68
[21] Pucher G W, Leavenworth C S, Vickery H B. Determination of starch in plant tissues., 1948, 20: 850–853
[22] 赵黎明, 李明, 郑殿峰, 顾春梅, 那永光, 解保胜. 水稻光合作用研究进展及其影响因素分析. 北方水稻, 2014, 44(5): 66−71 Zhao L M, Li M, Zheng D F, Gu C M, Na Y G, Xie B S. Analysis of the factors and their effect on the photosynthesis of rice., 2014, 44(5): 66−71 (in Chinese with English abstract)
[23] 邓启云, 袁隆平, 蔡义东, 刘建丰, 赵炳然, 陈立云. 超级杂交稻模式株型的光合优势. 作物学报, 2006, 32: 1287−1293 Deng Q Y, Yuan L P, Cai Y D, Liu J F, Zhao B R, Chen L Y. Photosynthetic advantages of model plant-type in super hybrid rice., 2006, 32: 1287−1293 (in Chinese with English abstract)
[24] 许明, 贾德涛, 马殿荣, 王嘉宇, 苗微, 陈温福. 北方超级粳稻根系生理、叶片光合性能特点及其相互关系. 作物学报, 2010, 36: 1030–1036 Xu M, Jia D T, Ma D R, Wang J Y, Miao W, Chen W F. Correlation of root physiology and leaf photosynthesis characteristics in northern Chinesesuper rice., 2010, 36: 1030–1036 (in Chinese with English abstract)
[25] Yang C M, Yang L Z, Yang Y X, Zhu O Y. Rice root growth and nutrient uptake as influenced by organic manure in continuously and alternately flooded paddy soils., 2004, 70: 67–81
[26] Osaki M, Shinano T, Matsumoto M, Zheng T G, Tadano T. A root-shoot interaction hypothesis for high productivity of field crops., 1997, 43: 1079–1084
[27] Inthapanya P, Sipaseuth, Sihavong P, Sihathep V, Chanphengsay M, Fukai S, Basnayake J. Genotypic performance under fertilized and non-fertilized conditions in rained lowland rice., 2000, 65: 1–14
[28] 董桂春, 陈琛, 袁秋梅, 羊彬, 朱正康, 曹文雅, 仲军, 周娟, 罗刚, 王熠, 黄建晔, 王余龙. 氮肥处理对氮素高效吸收水稻根系性状及氮肥利用率的影响. 生态学报, 2016, 36: 642–651 Dong G C, Chen C, Yuan Q M, Yang B, Zhu Z K, Cao W Y, Zhong J, Zhou J, Luo G, Wang Y, Huang J Y, Wang Y L. The effect of nitrogen fertilizer treatments on root traits and nitrogen use efficiency inrice varieties with high nitrogen absorption efficiency., 2016, 36: 642–651 (in Chinese with English abstract)
[29] 董桂春, 王余龙, 周娟, 张彪, 张传胜, 张岳芳, 杨连新, 黄建晔. 不同氮素籽粒生产效率类型籼稻品种氮素分配与运转的差异. 作物学报, 2009, 35: 149–155 Dong G C, Wang Y L, Zhou J, Zhang B, Zhang C S, Zhang Y F, Yang L X, Huang J Y. Difference of nitrogen accumulation and translocation in conventionalrice cultivars with different nitrogen use efficiency for grain output., 2009, 35: 149–155 (in Chinese with English abstract)
[30] Fageria N K. Yield physiology of rice., 2007, 30: 843–879
[31] Zhang H, Xue Y G, Wang Z Q, Yang J C, Zhang J H. Morphological and physiological traits of roots and their relationships with shoot growth in “super” rice., 2009, 113: 31−40
[32] 莫良玉, 吴良欢, 陶勤南. 高等植物GS/GOGAT循环研究进展. 植物营养与肥料学报, 2001, 7: 223−231 Mo L Y, Wu L H, Tao Q N. Research advances on GS/GOGAT cycle in higher plants., 2001, 7: 223−231 (in Chinese with English abstract)
[33] 王小纯, 熊淑萍, 马新明, 张娟娟, 王志强. 不同形态氮素对专用型小麦花后氮代谢关键酶活性及籽粒蛋白质含量的影响. 生态学报, 2005, 25: 802−807 Wang X C, Xiong S P, Ma X M, Zhang J J, Wang Z Q. Effect of different nitrogen forms on key enzyme activity involved in nitrogen metabolism and grain protein content in specialty wheat cultivars., 2005, 25: 802−807 (in Chinese with English abstract)
[34] Pang J Y, Palta J A, Rebetzke G J, Milroy S P. Wheat genotypes with high early vigour accumulate more nitrogen and have higher photosynthetic nitrogen use efficiency during early growth., 2014, 41: 215–222
[35] 王维. 适度土壤干旱对稻麦碳氮营养运转的调节作用及其机理. 扬州大学博士学位论文, 江苏扬州, 2003 Wang W. Soil Drying in Regulation to the Remobilization of Carbon and Nitrogen Reserves During Grain-filling Period of Wheat and Rice and Its Physiological Mechanism. PhD Dissertation of Yangzhou University, Yangzhou, China, 2003
[36] Samejima H, Kondo M, Ito O, Nozoe T, Shinano T, Osaki M. Characterization of root systems with respect to morphological traits and nitrogen-absorbing ability in new plant type of tropical rice lines., 2005, 28: 835–850
[37] Samejima H, Kondo M, Ito O, Nozoe T, Shinano T, Osaki M. Root-shoot interaction as a limiting factor of biomass productivity in new tropical rice lines., 2004, 50: 545–554
Comparison in Nitrogen Metabolism and Photosynthetic Characteristics betweenRice Varieties Differing in Nitrogen Sensitivity
JU Cheng-Xin1,2, ZHOU Zhu-Biao1, ZHAO Bu-Hong2, WANG Zhi-Qin1, and YANG Jian-Chang1,*
1Jiangsu Key Laboratory of Crop Genetics and Physiology / Co-innovation Center for Modern Production Technology of Grain Crops, Yangzhou University, Yangzhou 225009, Jiangsu, China;2Lixiahe Region Agricultural Research Institute, Yangzhou 225007, Jiangsu, China
Understanding the physiological characters associated with nitrogen (N) use efficiency (NUE) is very important in selecting and breeding N-efficient varieties. However, the information on such characters is very limited. Fourrice varieties, i.e., two nitrogen sensitive varieties, Huaidao 5 (HD-5) and Lianjing 7 (LJ-7), two nitrogen insensitive varieties, Ningjing 1 (NJ-1) and Yangjing 4038 (YJ-4), were grown in the field, and two N rates, 0 and 200 kg ha−1, were applied during the growing season.The yield components, NUE and aboveground physiological characters were investigated. The nitrogen sensitive varieties produced higher grain yield, exhibited higher NUE than nitrogen insensitive varieties at N rates of 0 and 200 kg ha−1.When compared with the nitrogen insensitive varieties, the nitrogen sensitive varieties had a higher photosynthetic rate and N accumulation, stronger activity of the enzymes involved in N metabolism, higher photosynthetic NUE, more accumulation of soluble sugars and starch in stems and sheaths at heading time, and more remobilization of nonstructural carbohydrate from stems to grains during grain filling. These physiological traits resulting in higher grain yield and nitrogen use efficiency are important and can be used as physiological indexes to select and breed high-yielding and N-sensitive rice varieties.
rice; nitrogen sensitivity; grain yield; nitrogen use efficiency; physiological characteristics
2017-08-28;
2017-11-21;
2017-12-18.
10.3724/SP.J.1006.2018.00405
本研究由国家自然科学基金项目(31461143015, 31471438, 31471447), 国家科技支撑计划项目(2014AA10A605), 国家重点研发计划项目(2016YFD0300206-4), 江苏高校优势学科建设工程项目(PAPD)和扬州大学高端人才支持计划项目(2015-1)资助。
This study was supported the National Natural Science Foundation of China (31461143015, 31471438, 31471447), the National Key Technology Support Program of China (2014AA10A605), the National Key Research and Development Support Program of China (2016YFD0300206-4), the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), and the Top Talent Supporting Program of Yangzhou University (2015-01).
Corresponding author杨建昌, E-mail: jcyang@yzu.edu.cn, Tel: 0514-87979317
E-mail: cxju1124@163.com
http://kns.cnki.net/kcms/detail/11.1809.S.20171218.0920.008.html
