鼠类脑细胞导航机理的移动机器人仿生SLAM综述
2018-03-12陈孟元
陈孟元
(1. 安徽工程大学 安徽省电气传动与控制重点实验室,安徽 芜湖 241000; 2. 中国科学技术大学 精密机械与精密仪器系,安徽 合肥 230027)
从简单重复的劳动中解放出来一直是人类追求的梦想,也是人类创造发明机器人的主要目的之一。机器人可移动性可以进一步扩大其使用范围并能更好地提高其使用效率,但移动机器人在复杂环境中如何模仿人类进行自我导航和路径规划一直是难以解决的问题。
20世纪七八十年代,随着人类智能和移动机器人的出现,人类将两者进行了有效地关联并开始了有益地探索,人工智能领域进入了一个蓬勃发展的新时代。进入21世纪后,被定义为研究感知与行动之间智能连接的移动机器人学被赋予了更多的内涵,随着工业的发展,移动机器人正在从事工业制造、生活家居、通信运输工作,甚至对空间以及深海进行探索,可以说,移动机器人的出现改变了人们的生活,对社会发展的各个方面产生了巨大的影响,为先进技术的发展与社会的进步做出了有益的贡献。
移动机器人在短短几十年间经过了突飞猛进的发展,能够较好地代替人们完成重复性和危险性的劳动,减轻人们的工作强度和危险性,提高生产效率。而同步定位与地图构建(SLAM)问题一直是限制移动机器人在复杂环境下应用的关键原因,移动机器人在没有提前预设好行进路线的陌生环境中需要主动了解实际环境中的路况信息和障碍物的实际位置,同时精确地进行移动机器人自身的定位与实际环境的地图构建。现阶段,移动机器人主要采用自身携带的各种图像采集、陀螺仪、加速度计等先进传感器,通过激光测距、超声测距以及图像匹配完成SLAM工作。
传感器在实际环境中的测量值有可能是错误的,例如,利用激光照射玻璃墙,激光光束可能已经经过了多次反射才反射回探测器。图像采集传感器也有其局限性,例如,普通照相机在区分图像标准色方面效果较差,若采用黑白色的灰度方式表示图像,光线强度的变化又很难表现出来,若采用全景相机或广角镜头采集图像,采集到的图像又会产生畸变。另一方面,即使传感器测量值是正确的,但总存在难以克服的精度问题。以测距和图像匹配方法实现SLAM为例,虽然传感器具有较高精度,可以通过轮子上安装的里程计准确测量机器人的移动距离,但由于路面不平坦和轮子滑动等原因,传感器测量结果不能准确反映机器人的实际移动距离,并且随着移动机器人工作时间的推移累积误差也在不断叠加,这也是不能忽略的重要方面。因此科学家开始寻找更加可靠的定位导航方式。
1 鼠类相关导航脑细胞
当人类还处于原始社会时就已经意识到“飞鸽传书,老马识途”。一些动物具有出类拔萃的导向能力,无论阴晴雨雪,纵使万水千山,这些动物总能知道路在何方,准确找到目的地。经过多年的研究,2014年诺贝尔生理学或医学奖获得者发现了基于动物导航机制的大脑定位系统细胞。
像人们熟知的GPS系统一样,大脑定位系统也是通过采集自身运动的时间、位置信息进行定位导航的。秀丽隐杆线虫仅有几百个神经细胞,却可以通过追寻环境中的嗅觉信号来判断方向。对于蚂蚁和蜜蜂等神经系统更为复杂的动物,它们利用神经细胞实时监测自身的运动方向和速度变化,通过路径整合获得当前所在的位置。这种方法使得动物仅依靠自身神经系统进行导航。
哺乳类动物辨识方向的方法更为先进。人类作为高级的哺乳动物由于受到伦理的约束无法进行大量的实验研究。与人较为接近的哺乳动物,如猩猩、狒狒等,由于数量和国家保护的因素一般也不选用。在生物地图构建和导航领域,人们通常选用哺乳纲中的鼠类动物作为实验模型,由于实验环境和场合广泛,为研究动物的导航性能奠定了良好的基础。此外,鼠类大脑也是动物研究领域使用最多的一块区域。
大脑中不同的神经细胞由于受到刺激产生兴奋导致膜电位的变化,这些同时产生兴奋的神经细胞所组成的图案恰能反映外界环境的空间布局和自身在环境中所处的位置[1]。鼠类大脑中存在3种主要导航细胞:位置细胞、头方向细胞以及网格细胞。其中,位置细胞绘制所处地点的地图;头方向细胞指明方向(将位置细胞和头方向细胞合并形成的一个新细胞类型,位姿细胞);网格细胞通过标记被激活细胞的位置对场景进行重定位。此外,在内嗅皮层还存在边界细胞、速度细胞等能够进行辅助导航的细胞,如图1所示。

图 1 海马体和内嗅皮质构成的系统生物模型Fig. 1 Biological model consist of hippocampus and entorhinal cortex
在哺乳类动物中枢神经系统的大脑皮质中,存在一种形状类似于海马,功能上负责短期记忆储存和学习的区域,被称为海马体。在日常中的一个记忆片段,比如一个电话号码在短时间内被重复提及,经过海马体可以将其转存入大脑皮层,成为永久记忆。1971年伦敦大学学院(university college London)的美国科学家O’Keefe等[2]在海马脑区发现了一种特殊的神经细胞,这种细胞在鼠类经过空间某特定位置时其中的一些细胞会产生兴奋,而经过另一个位置时另一些细胞会产生兴奋,他将这种细胞命名为位置细胞。1984年纽约大学的James Ranck等[3]发现一组与动物的方向感知直接相关的神经细胞并命名为头方向细胞,头方向细胞能够辨别头部的朝向。当头部朝向北方的时候,一组细胞会兴奋;而头部转向南方时,另一组细胞兴奋,通过这两种细胞的兴奋状态能够准确判断出在某一时间老鼠所处的精确位置。除此之外,内嗅皮质也参与整个信号的传递过程,内嗅皮质层从网格细胞处将有关方向和移动距离的信息传递给海马体,如图2所示。内嗅皮质将关于动物运动的方向和距离信息通过不同的神经通路传递至海马体中的齿状回(dentate gyrus,DG)、CA3和位置细胞所在的CA1等区域,由此产生的大脑认知地图可以帮助动物更好地规划未来的“旅程”。

图 2 海马体和内嗅皮层横截面视图Fig. 2 Section lateral of hippocampus and entorhinal cortex
老鼠通过各种感官从环境中获取外界的特征信息,而位置细胞则能够和海马体中其他细胞合作,将输入的特征信息与过往记录到的特征信息加以比对。一旦信息能够匹配成功,与匹配位置相对应的特定位置细胞就会变得活跃。通过这种方法,大脑能够将特定的特征信息与特定的空间位置联系起来,形成空间位置记忆,再通过与头方向细胞协作,可以构建位置细胞和头方向细胞所表征的海马神经人工模型。图3表示老鼠在空旷房间的运动轨迹,圆锥形表示老鼠,圆斑表示理论上细胞的活跃位置,圆点表示记录下的某一批位置细胞活跃时的位置。实验表明,分离的位置细胞和头方向细胞所表征的海马神经人工模型无法长期跟踪老鼠的位置,而将位置细胞和头方向细胞合并形成的一个被称为位姿细胞的新细胞类型则对老鼠起到良好的导航作用。
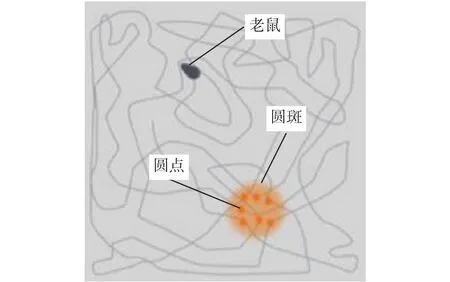
图 3 老鼠运动轨迹和海马体位置细胞分布Fig. 3 The trajectory of rats and the distribution of hippocampus
2005年挪威大学科学与技术学院的May-Britt Moser[4-5]在海马区以外的内嗅皮质的脑区里发现了一种全新的神经细胞并将其命名为网格细胞。图4为老鼠在空旷房间中运动时,老鼠运动轨迹和某一个网格细胞的活跃位置,可以看出这些被激活细胞的位置呈现均匀六边形分布。也就是说,任意一个网格细胞的发放场在空间中均匀分布,并且呈现出一种蜂巢式的六边形网格状。虽然网格细胞的活跃也和动物所处的位置有关,但是与位置细胞不同,网格细胞的活跃并不依赖于外界输入的特征信息。

图 4 老鼠运动轨迹和内嗅皮质网格细胞分布Fig. 4 The trajectory of rats and the distribution of grid cells in entorhinal cortex
2008年,研究者们从内嗅皮质中发现一种新的细胞类型,当动物接近墙壁、围场边缘或是其他隔断时,这些细胞就会放电,这种细胞能够通过细胞活性计算自身到达边界的距离,研究者们将其命名为边界细胞。2015年,在获得诺贝尔生理学或医学奖后,May-Britt Moser和Edvard Moser夫妇继续发现一些神经细胞能够随移动速度的提升成比例地提升放电率,通过查看这种细胞的放电频率便能够判断一个动物在给定时间点上的移动速度,研究者们将其命名为速度细胞。
研究表明,在基于鼠类海马体的移动机器人导航研究基础上,引入速度细胞能够更精准的实现移动机器人速度和角度的测量,融合边界细胞能够实现突发障碍物等复杂环境的导航[6]。
2 国内外研究现状及分析
在过去30多年内,对SLAM问题的探索研究大多是基于数学概率的方法,其中最成功的3种概率法为卡尔曼滤波(Kalman filter,KF)算法、最大期望 (expectation maximisation,EM)算法[7]和粒子滤波(particle filter,PF)算法。卡尔曼滤波算法只适用于线性系统,具有很大的局限性;经典的扩展卡尔曼滤波(extended Kalman filter,EKF)[8-9]通过对非线性函数泰勒展开式一阶线性化截断将非线性问题线性化,可以应用于有非线性特征的简单系统中;无迹卡尔曼滤波(unscented Kalman filter,UKF)[10-11]通过无迹(unscented transformation,UT)变换处理均值和协方差的非线性传递,计算精度较高。Arasaratnam等提出的容积卡尔曼滤波(cubature Kalman filter,CKF)算法结构简单[12],估计精度高,进一步提升了系统的稳定性。粒子滤波也称为蒙特卡罗定位 (Monte Carlo localisation,MCL)[13-14],这种算法利用带权重粒子的分布来估算要求的位置分布,故无需考虑后验分布所带来的形状或特征约束。与标准MCL算法一样,快速同步定位和地图构建系统(FastSLAM)算法通过粒子滤波器估计机器人路径下一时刻的位置[15]。这些概率算法的优势是能够处理传感器与环境的模糊性,有很好的SLAM性能,但这些方法是对当前采集到的环境进行数学建模并进行优化,因此不能完全解决全局地图构建和导航问题。
相比之下,许多生物虽然不具备高精度传感器,但仍具有较强的导航能力,并且能够解决全局SLAM问题。因为生物对其地图的构建依赖于鲁棒性处理策略而不是对环境的精确描述,Wehner等[16]发现蚂蚁返回蚁穴邻近区域后,再利用一整套搜索策略找到蚁穴,这种小尺寸和有限复杂度的环境有效降低了地图构建和导航的难度。但这些成熟的生物学导航算法模型仍具有局限性,例如蚁群算法计算量大、搜索时间长、易陷于局部最优解等问题无法得到有效解决。
拥有海马体的哺乳动物,如鼠类是人们研究较多的一种动物,人们对其导航、环境探索和真实能力的神经机制也理解得较为深入[17-21]。Skaggs等[22]利用两套旋转细胞群,通过Hebbian学习规则校正局部场景细胞和头方向细胞的连接,头方向细胞形成的外环是活跃细胞编码方向,内部两环是反应角速度的前庭细胞,如图5所示。Stringer等[23-25]通过头方向细胞、视觉输入细胞和前向速度细胞互相激励来描述位置细胞活性,从而描述环境中物体运动的状态,如图6所示。这种包含分离头方向位置表征系统的海马神经人工模型,尽管可以支持位置和多方向假设,但由于不能随时间推移支持多位姿假设,故会造成不完全重定位或重定向,从而导致无法长期跟踪移动机器人的位置。

图 5 鼠类头方向系统的吸引子网络模型Fig. 5 Attractor network model of head direction system of rats
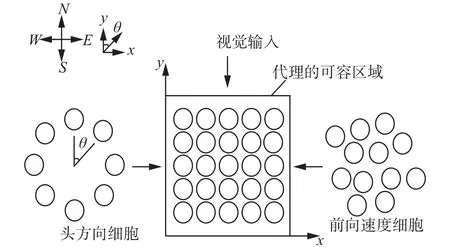
图 6 位置细胞的二维连续吸引子模型Fig. 6 Two-dimensional continuous attractive model of pose cells
针对上述海马模型的缺陷,研究者们提出一种基于复合位姿表征的啮齿动物海马区扩展模型,将路径积分和视觉关联过程集成到位姿细胞模型中,使移动机器人具有更新预测状态的能力。实验表明,啮齿动物海马区扩展模型对于某一环境产生了稳定的、一致的、具有正确拓扑的地图,但是面对范围更广更复杂的环境导致路径积分性能下降时,会出现冲突和多重表征两种附加现象[26-29]。
后人基于此对啮齿动物海马区扩展模型进行改进,一种借鉴策略是直接根据移动机器人位姿的核心表征,以及存储在位姿感知细胞及局部场景细胞网络中的环境表征对应关系,建立存储环境空间、视觉、时间、行为和变化等信息的沿途构图[30-33]。该算法通过同时保留位姿感知细胞地图拓扑连通性和保持细胞之间的空间信息,解决了非连续、冲突和多重表征这3个问题。Milford等验证了这种具有环境探索、目标记忆和自适应改变的扩展啮齿动物海马区模型能在不同天气和地形下稳定导航。文献[34]提出一种新型的基于自组织特征图的神经网络模型并将其引入到局部场景细胞,提高了系统的实时性和准确性。文献[35]通过飞行器进行场景回忆,验证了沿途构图算法的可靠性。对于车载导航系统,由于视觉里程计误差较大,即使通过啮齿动物海马区扩展模型仿生机制的修正也仍无法准确导航,张潇等[36]在此基础上引入光学双轴传感器和MIMU信息,建立了航位推算模型。文献[37]提出将FAB-MAP(fast appearance-based mapping)引入啮齿动物海马区扩展模型的视觉里程计,这种基于历史模型的闭合检测算法可以过滤视觉里程计的误报信息,进一步提高该模型的稳定性,但通过逐个对比当前场景与历史场景的相似性,实施闭环检测的方法效率较低,不能满足系统实时性要求。研究提出的基于实时关键帧匹配的闭环检测模型在保证SLAM稳定的前提下能够提高闭环检测实时性。
仅仅依赖纯视觉的导航算法并不能适应长时间复杂环境下的导航,研究者们将现有的传感器技术应用于已知的几种导航细胞下进行导航。许曈等[38]提出将姿态策略模块引入啮齿动物海马区扩展模型中,对突发障碍物的干扰提出了一个切实可行的建议,Berkvens等[39]将无源 RFID、有源 RFID、WI-FI、地磁传感器分别引入多组实验中,结果表明,地磁通量引入的啮齿动物海马区扩展模型算法可以明显减少射频技术沿途构图的平均误差,并在很大程度上确保无错误路径的产生,但仍有问题有待解决,如由于前一时刻定位产生错误导致预期位置的错误判断、沿途构图的平均误差较大等问题。这些利用现有传感器来解决移动机器人复杂环境下的导航问题,一方面对传感器的性能要求较高,另一方面,各类传感器也会受到复杂环境的干扰。研究提出一种复合鼠类导航细胞模型,融合速度细胞可更精准实现移动机器人速度和旋转角度的测量,融合边界细胞以实现突发障碍物的判别。
现有的SLAM理论经过30余年的发展,基于数学概率方法和从距离到拓扑的地图表征法已经较好地完成了“定位”和“建图”过程,但都仍然存在一定的局限性。由于实际环境十分复杂,例如光线变化、太阳东升西落、不断的有人进出等。即使让一个机器人以5 cm/s的速度在安静的空间里慢慢移动,这种论文中看起来可行的算法,在实际环境中往往捉襟见肘,处处碰壁,因此,SLAM还未走进人们的实际生活。这种局限性是数学模型本身带来的,与改进算法无关。因此,在考虑人类能如此轻松地完成导航的基础上,提出构建一种仿动物大脑细胞导航机制下的同步定位与地图构建方法。
本研究受到2014年“诺贝尔生理学或医学奖”启发,拟将发现的局部场景细胞、位姿细胞(位置细胞、头方向细胞)、网格细胞、速度细胞、边界细胞等具有定位导航功能的细胞模型应用于SLAM研究中。分别研究5种导航细胞各自的工作机理和数学表达,然后将这5种导航细胞按照动物导航机制下大脑的工作原理建立数学模型,最终进行实验验证。
3 未来深入研究的方法建议
提出一种未来深入研究的方法建议,拟将包含局部场景细胞(view cells)、位姿细胞(pose cells)、网格细胞(grid cells)、速度细胞(speed cells)、边界细胞(border cells)等具有定位导航功能的鼠类脑细胞模型应用于SLAM研究中,构建一种衍生SLAM算法(BVGSP-SLAM)实现复杂场景下的导航。融入局部场景细胞、位姿细胞和网格细胞模型,尽可能减小光线变化对视觉里程计产生的影响;在此基础上引入速度细胞和边界细胞,对突发障碍物和自身速度进行进一步判别。
局部场景细胞获取当前场景,位姿细胞获取当前状态,获得具有一定导航能力的仿生导航系统VP-SLAM,但这种系统会受到现实场景中人物走动、光线变化等原因造成导航性能下降。为进一步提升系统的鲁棒性,融入网格细胞,构建了实时关键帧匹配的闭环检测模型GVP-SLAM,避免因光线变化导致SLAM不稳定,从而既可大幅提高复杂环境下导航的精准性,又具有较好的实时性。在提出带实时闭环检测的鼠类导航细胞模型基础上,引入边界细胞和速度细胞两种导航细胞的导航原理,提出一种带实时闭环检测的复合鼠类导航细胞模型BVGSP-SLAM。该模型通过融合速度细胞更精准地实现移动机器人速度的测量,避免基于局部场景细胞在突发障碍物影响下的判别失效;通过融合边界细胞,实现移动机器人对移动障碍物的实时避障,进一步提高了SLAM的稳定性,技术路线图如图7所示。
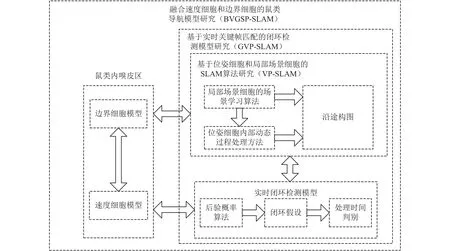
图 7 融入鼠类脑细胞导航机制的移动机器人衍生SLAM方法技术路线图Fig. 7 Technology roadmap of derivative SLAM on mobile robots integrated with brain cells of rats
3.1 基于位姿细胞和局部场景细胞的SLAM算法(VP-SLAM)
局部场景细胞在环境中学习独特场景,模型化头方向细胞和位置细胞的竞争性吸引子网络结构所形成的位姿感知细胞表征当前位置,局部场景细胞和位姿细胞协同完成拓扑化经验图的绘制。在此过程中,抽象出融合头方向与位置模型特性的信息需要通过某种关联算法进行姿态表达,在相机图像信息经过处理形成局部场景后,需选择合适的视觉SLAM算法进行图像处理。
现有基于鼠类导航策略的扩展海马模型,其分别由局部场景细胞在环境中学习独特场景,由头方向细胞和位置细胞合并形成的位姿细胞表征当前位置,以及用节点与链路编码局部场景细胞和位姿细胞构建拓扑化的经验图,如图8所示。通过(x, y,θ)关联一维头方向细胞模型θ与二维位置细胞模型(x, y),实现位姿细胞的构建,其中,连续吸引子网络(continuous attractive network,CAN)控制着位姿感知网络内部的活动。其动态过程经历3个阶段:兴奋度更新阶段、对所有细胞的全局抑制阶段以及对位姿感知细胞活动的归一化阶段。
对于局部场景信息的处理,通过Hessian矩阵行列式对图像中的像素点进行分析,构造快速鲁棒特征(speeded up robust feature,SURF)的特征点描述算子。其中,每个特征点采用64维向量的描述子进行匹配。
3.2 基于实时关键帧匹配的闭环检测模型(GVP-SLAM)
视觉里程计漂移会形成累计误差,通过闭环检测可以修正复杂环境下定位导航里程计产生的累计误差。现有IAB-MAP(incremental appearance-based mapping)和FAB-MAP(fast appearance-based mapping)闭环检测算法,虽能胜任复杂环境下的闭环检测问题,但由于它们通过逐个比较当前帧数据与各历史帧数据的相似性,无法满足实时性要求。本文基于的导航细胞模型在借鉴网格细胞场景重定位的基础上设计相关算法提高实时性能,构建的实时关键帧匹配的闭环检测模型能够明显提高系统的实时性能。
在闭环检测过程中,要对采集到的足够多的陌生场景进行信息匹配。一方面,必须通过某种手段对采集到的连续图像和情境重现加以区分以避免误判;另一方面,由于复杂环境的影响,采集的某一场景图像信息可能会发生错误辨识。对于闭环检测方法本身,需通过一种策略收集频次较高和最临近时刻出现的信息作为被匹配对象,在节约时间成本的条件下尽可能地准确匹配当前图像信息。与一般的机器视觉算法不同,上述工作均需基于鼠类导航细胞。
借鉴网格细胞场景重定位并在此基础上设计相关算法提高实时性能,构成一种基于实时关键帧匹配的闭环检测模型,通过局部场景细胞进行实时关键帧匹配,实现闭环检测,通过位姿细胞和局部场景细胞沿途构图。如图8所示,实时关键帧匹配的闭环检测模型通过局部场景细胞进行新场景匹配的判断,并通过局部场景细胞与位姿细胞关联影响沿途构图。

图 8 基于鼠类导航细胞的实时关键帧匹配的闭环检测模型Fig. 8 Closed loop detection model with real-time keyframe matching based on rats
本研究提出的实时关键帧匹配的闭环检测模型具体流程包括定位点的建立、权重更新、贝叶斯估计更新、闭环假设选择、恢复和转换6部分,并引入了3种记忆模式,即工作记忆(working memory,WM)、长期记忆(long-term memory,LTM)和短时记忆(short-term memory,STM),具体策略如下。首先采集陌生场景信息,为避免实际环境的干扰,场景采集时需预先对同一场景多次采集并过滤其中的错误信息,确保场景采集的准确性。再通过设定阈值避免将当前采集到的连续图像误判为闭环。将当前位置时刻至过去某时刻内的信息储存在短时记忆中,并选取所有过去时刻出现频次最高的信息储存在工作记忆中。最后通过当前位置与短时记忆存储位置比较进行新位置的判别和权重更新,将当前位置与工作记忆存储位置比较进行闭环检测,同时,实时更新各记忆本身。此闭环检测模型的流程图如图9所示,各记忆关系图如图10所示。
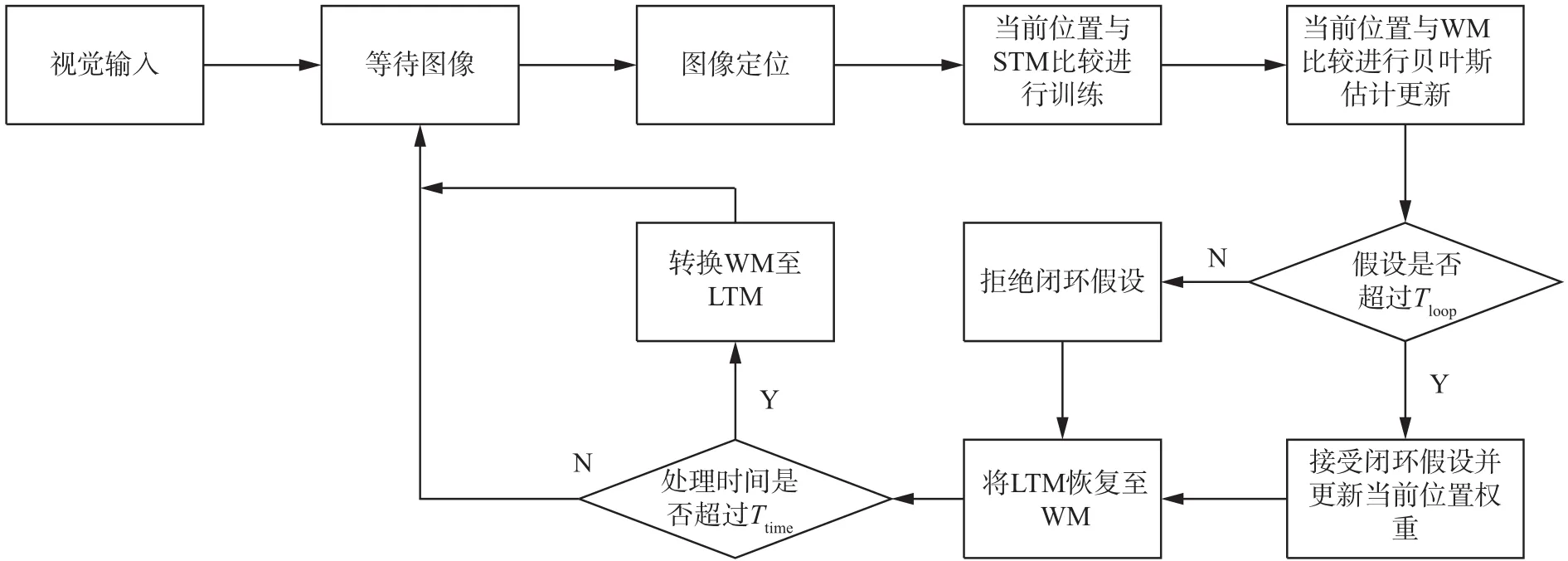
图 9 实时关键帧匹配的闭环检测模型流程图Fig. 9 Flow chart of the closed loop detection of real-time keyframe matching

图 10 实时关键帧匹配的闭环检测中各记忆模式关系图Fig. 10 Diagram of memory pattern in the closed loop detection of real-time keyframe matching
3.3 融合速度细胞和边界细胞的鼠类导航模型(BVGSP-SLAM)
现有的位姿细胞无法适应外界多变的复杂环境,如移动障碍物的出现会导致视觉里程计速度的错误判别。根据发现的内嗅皮层中存在的速度细胞和边界细胞,针对室内复杂环境的特点,利用速度细胞和边界细胞的导航功能对移动机器人角度和速度等信息进行采集分析,输出到终端设备并与鼠类导航细胞的沿途构图进行比对。在3.2节研究的GVP-SLAM算法基础上,引入速度细胞和边界细胞实现移动机器人在突发障碍物等复杂环境下的精确导航,构成复合鼠类导航模型(BVGSP-SLAM)。
在GVP-SLAM算法基础上引入内嗅皮层中的边界细胞和速度细胞进行辅助导航。为探究其效果,将速度细胞和边界细胞对鼠类导航模型的影响进行分别研究。利用手术在老鼠脑部植入电极,通过上位机可观测速度细胞和边界细胞的活性并建立数学模型。
1) 速度细胞活性及数学模型建立
在分析速度细胞活性的实验中,将老鼠置于100 cm×100 cm×50 cm的盒子中,通过在随机位置播撒食物屑促使老鼠进行任意运动,如图11所示。为避免局部场景细胞对实验的影响,该实验在黑暗环境中进行;为避免老鼠自身行为对速度细胞产生的影响,本实验忽略所有运动速度小于2 cm/s时老鼠速度细胞活性的变化。首先归一化速度细胞活性;通过线性变换计算出速度细胞的发放率并表示其活性;然后对细胞进行无偏分析,通过无偏估计在实验中调整速度细胞参数,利用尖峰电压大小判断速度细胞的活性强弱,通过一个由发放场和线性滤波器两部分组成的简单线性解码器使速度细胞的活跃度具体化;再将该活性状态信息传递给由头方向细胞和位置细胞融合而成的位姿细胞,进而影响沿途构图。

图 11 黑暗环境下的速度细胞实验及速度细胞分析过程Fig. 11 Speed cells experiment in dark environment and the analysis procedure of speed cells
2) 边界细胞活性及数学模型建立
边界细胞对于障碍物的判别,如图12所示,无论老鼠的头部朝向什么方向,边界细胞的发放率都会随老鼠离障碍物距离的减少而增加。每个边界细胞的感受野通过两个高斯函数的乘积构成,一个函数表示特定的距离,另一个函数表示非自我中心的方向。利用边界细胞的发放率、边界细胞的感受野及其与障碍物的距离信息构成边界细胞发放率变化的表达式,这种细胞的发放率变化状态即表现为细胞的活性状态,再将该活性状态信息通过神经元的感知机模型进行分析,并把所有速度细胞活性信息传递给由头方向细胞和位置细胞融合而成的位姿细胞,进而影响沿途构图。

图 12 老鼠边界细胞的感受野以及老鼠距离障碍物不同距离时发放率强度Fig. 12 Receptive field of border cells and firing rate at various distances of rats
3) 边界细胞与速度细胞模型的关联函数构造构造由边界细胞和速度细胞发放率、感受野以及距离信息的关联函数,采用统计学的方法剔除异常细胞,通过构建权值、中心和幅度3个参数可调的自适应模糊逻辑系统证明该关联函数的稳定性,进而提高BVGSP-SLAM模型的鲁棒性。
3.4 所需突破的关键性问题
复杂室内环境下SLAM存在如下问题:室内光线变化、突发障碍物等对导航系统产生影响,对于移动机器人本身,长时间导航下移动机器人也会存在里程计漂移等现象。本文针对这些问题建立了适合复杂室内环境的基于鼠类导航细胞的SLAM模型,所需突破的关键问题主要包括如下两点。1) 实时关键帧匹配的闭环检测建模现有的闭环检测模型存在复杂度高、实时性差等问题,本文提出了一种实时关键帧匹配的闭环检测模型,该模型通过“长期记忆”、“短时记忆”等记忆单元的协同作用完成关键帧匹配,同时可以通过“长期记忆”和“短时记忆”单元的切换和恢复,降低系统运算的复杂度,这也是后续融入速度细胞模型和边界细胞模型完成实时导航的基础。因此,该问题是研究内容待解决的关键问题之一。2) 边界细胞和速度细胞模型关联函数的构造边界细胞和速度细胞模型分别由其发放率、感受野以及距离信息等动态参数决定,两种模型通过关联函数各自影响沿途构图。采用统计学的方法剔除异常细胞,通过构建权值、中心和幅度3个参数可调的自适应模糊逻辑系统证明该关联函数的稳定性,研究基于边界细胞和速度细胞模型关联的融合方法,将多种调度参数信息融入到控制方法的设计中,这也是沿途构图准确度提高面临的主要问题。因此,该问题是研究内容待解决的另一关键问题。
4 结束语
基于鼠类脑细胞导航机理的模型算法鲁棒性强,能够解决全局SLAM问题。综观这种基于鼠类脑细胞导航机理的模型算法的研究现状,其相关的研究仍处于初级阶段,有很多问题值得进一步的研究,本文提出运用仿生学研究、医工结合、数值计算方法、解析计算方法,结合信息学、控制学、仿生学等基础理论并建立适用于复杂环境下快速、准确、全面的融入鼠类脑细胞导航机制的移动机器人衍生同步定位地图构建方法体系。本文重点探讨了以鼠类脑细胞中边界细胞、局部场景细胞、网格细胞、速度细胞、位姿细胞等具有定位导航功能的细胞模型应用于SLAM研究中,构建一种基于多细胞导航机制的BVGSP-SLAM模型:融入局部场景细胞、位姿细胞和网格细胞以尽可能减少光线变化对视觉里程计的影响,引入边界细胞和速度细胞以对突发障碍物和自身速度进行进一步判别,再将混合细胞算法引入仿生传感系统并在移动机器人平台上进行硬件验证,为移动机器人SLAM研究领域提供了生物细胞学重要解决思路的理论参考。在此基础上如何实现在实验场景更加恶劣的情况下动态调节现有细胞导航模型的参数仍值得进一步研究,如何通过现有的鼠类模型进行算法改进或借鉴其他生物模型特性实现空中与水下的导航研究也将是未来SLAM发展的一个重要研究方向。
[1]TOLMAN E C. Cognitive maps in rats and men[J]. Psychological review, 1948, 55(4): 189–208.
[2]O’KEEFE J, DOSTROVSKY J. The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat[J]. Brain research, 1971, 34(1): 171–175.
[3]RANCK J L, LETELLIER L, SHECHTER E, et al. X-ray analysis of the kinetics of Escherichia coli lipid and membrane structural transitions[J]. Biochemistry, 1984, 23(21):4955–4961.
[4]于乃功, 王琳, 李倜, 等. 网格细胞到位置细胞的竞争型神经网络模型[J]. 控制与决策, 2015, 30(8): 1372–1378.YU Naigong, WANG Lin, LI Ti, et al. Competitive neural network model from grid cells to place cells[J]. Control and decision, 2015, 30(8): 1372–1378.
[5]于平, 徐晖, 尹文娟, 等. 网格细胞在空间记忆中的作用[J].心理科学进展, 2009, 17(6): 1228–1233.YU Ping, XU Hui, YIN Wenjuan, et al. The roles of grid cells in spatial memory[J]. Advances in psychological science, 2009, 17(6): 1228–1233.
[6]KROPFF E, CARMICHAEL J E, MOSER M B, et al.Speed cells in the medial entorhinal cortex[J]. Nature, 2015,523(7561): 419–424.
[7]DEMPSTER A P, LAIRD N M, RUBIN D B. Maximum likelihood from incomplete data via the EM algorithm[J].Journal of the royal statistical society, 1977, 39(1): 1–38.
[8]SMITH R C, CHEESEMAN P. On the representation and estimation of spatial uncertainly[J]. International journal of robotics research, 1987, 5(4): 56–68.
[9]SMITH R, SELF M, CHEESEMAN P. Estimating uncertain spatial relationships in robotics[J]. Machine intelligence and pattern recognition, 1988, 5(5): 435–461.
[10]JULIER S J, UHLMANN J K. Unscented filtering and nonlinear estimation[J]. Proceedings of the IEEE, 2004,92(3): 401–422.
[11]JULIER S, UHLMANN J, DURRANT-WHYTE H F. A new method for the nonlinear transformation of means and covariances in filters and estimators[J]. IEEE transactions on automatic control, 2000, 45(3): 477–482.
[12]ARASARATNAM I, HAYKIN S. Cubature Kalman filters[J]. IEEE transactions on automatic control, 2009,54(6): 1254–1269.
[13]NIEDERREITER H. Random number generation and quasi-monte carlo methods[J]. Journal of the american statistical association, 1992, 88(89): 147–153.
[14]THRUN S, FOX D, BURGARD W, et al. Robust Monte Carlo localization for mobile robots[J]. Artificial intelligence, 2001, 128(1): 99–141.
[15]MONTEMERLO M, THRUN S, WHITTAKER W. Conditional particle filters for simultaneous mobile robot localization and people-tracking[C]//Proceedings of 2002 IEEE International Conference on Robotics and Automation.Washington, DC, USA, 2002: 695–701.
[16]WEHNER R, GALLIZZI K, FREI C, et al. Calibration processes in desert ant navigation: vector courses and systematic search[J]. Journal of comparative physiology A, 2002,188(9): 683–693.
[17]刘新玉, 海鑫, 尚志刚, 等. 利用粒子滤波重建位置细胞编码的运动轨迹[J]. 生物化学与生物物理进展, 2016,43(8): 817–826.LIU Xinyu, HAI Xin, SHANG Zhigang, et al. Decoding movement trajectory of hippocampal place cells by particle filter[J]. Progress in biochemistry and biophysics, 2016,43(8): 817–826.
[18]胡波, 隋建峰. 海马位置细胞空间信息处理机制的研究进展[J]. 中华神经医学杂志, 2005, 4(4): 416–418.HU Bo, SUI Jianfeng. Advance of mechanisms of spatial processing for hippocampal place cells[J]. Chinese journal of neuromedicine, 2005, 4(4): 416–418.
[19]王可, 张婷, 王晓民. 大脑中的“定位系统”——2014年诺贝尔生理学或医学奖简介[J]. 首都医科大学学报, 2014,35(5): 671–675.WANG Ke, ZHANG Ting, WANG Xiaomin. “Inner GPS”in the brain——introduction of Nobel Prize in Physiology or Medicine 2014[J]. Journal of capital medical university,2014, 35(5): 671–675.
[20]田莉雯. 基于顶部摄像头和鼠载摄像头的大鼠自动导航系统[D]. 杭州: 浙江大学, 2015.TIAN Liwen. An automatic navigation system based on a rat-mounted camera and a bird’s eye camera[D]. Hangzhou: Zhejiang University, 2015.
[21]查峰, 肖世德, 冯刘中, 等. 移动机器鼠沿墙导航策略与算法研究[J]. 计算机工程, 2012, 38(6): 172–174.ZHA Feng, XIAO Shide, FENG Liuzhong, et al. Research on wall-following navigation strategy and algorithm for mobile mechanical mouse[J]. Computer engineering, 2012,38(6): 172–174.
[22]SKAGGS W E, KNIERIM J J, KUDRIMOTI H S, et al. A model of the neural basis of the rat's sense of direction[J].Advances in neural information processing systems, 1995,7: 173–180.
[23]REDISH A D, ELGA A N, TOURETZKY D S. A coupled attractor model of the rodent head direction system[J]. Network: computation in neural systems, 1997, 7(4): 671–685.
[24]SAMSONOVICH A, MCNAUGHTON B L. Path integration and cognitive mapping in a continuous attractor neural network model[J]. Journal of neuroscience: the official journal of the society for neuroscience, 1997, 17(15): 5900–5920.
[25]STRINGER S M, ROLLS E T, TRAPPENBERG T P, et al. Self-organizing continuous attractor networks and path integration: two-dimensional models of place cells[J]. Network: computation in neural systems, 2002, 13(4): 429–446.
[26]MILFORD M J, WYETH G F, PRASSER D. RatSLAM: a hippocampal model for simultaneous localization and mapping[C]//Proceedings of 2004 IEEE International Conference on Robotics and Automation. New Orleans, LA,USA, 2004: 403–408.
[27]MILFORD M, WYETH G. Persistent navigation and mapping using a biologically inspired SLAM system[J]. International journal of robotics research, 2009, 29(9): 1131–1153.
[28]PRASSER D P, WYETH G, MILFORD M. Experiments in outdoor operation of RatSLAM[C]//Proceedings of 2004 Australasian Conference on Robotics and Automation.Canberra, Australia, 2004: 1–6.
[29]PRASSER D, MILFORD M, WYETH G. Outdoor simultaneous localisation and mapping using RatSLAM[C]//Proceedings of the Results of the 5th International Conference.Berlin, Heidelberg, Germany, 2006: 143–154.
[30]MILFORD M J, PRASSER D, WYETH G. Effect of representation size and visual ambiguity on RatSLAM system performance[C]//Proceedings of 2006 Australasian Conference on Robotics and Automation. Auckland, New Zealand, 2006: 1–8.
[31]MILFORD M, SCHULZ R, PRASSER D, et al. Learning spatial concepts from RatSLAM representations[J]. Robotics and autonomous systems, 2007, 55(5): 403–410.
[32]MILFORD M, WYETH G, PRASSER D. RatSLAM on the edge: revealing a coherent representation from an overloaded rat brain[C]//Proceedings of 2006 IEEE/RSJ International Conference on Intelligent Robots and Systems.Beijing, China, 2006: 4060–4065.
[33]DHANDE O S, HUBERMAN A D. Retinal ganglion cell maps in the brain: implications for visual processing[J].Current Opinion in Neurobiology, 2014, 24(1): 133.
[34]许曈, 凌有铸, 陈孟元. 一种融合DGSOM神经网络的仿生算法研究[J]. 智能系统学报, 2017, 12(3): 405–412.XU Tong, LING Youzhu, CHEN Mengyuan. A bio-inspired algorithm integrated with DGSOM neural network[J]. CAAI transactions on intelligent systems,2017, 12(3): 405–412.
[35]MILFORD M J, SCHILL F, CORKE P, et al. Aerial SLAM with a single camera using visual expectation[C]//Proceedings of 2011 IEEE International Conference on Robotics and Automation. Shanghai, China, 2011:2506–2512.
[36]张潇, 胡小平, 张礼廉,等. 一种改进的RatSLAM仿生导航算法[J]. 导航与控制, 2015, 14(5): 73–79.ZHANG Xiao, HU Xiaoping, ZHANG Lilian, et al. An improved bionic navigation algorithm based on RatSLAM[J].Navigation and control, 2015, 14(5): 73–79.
[37]MADDERN W, GLOVER A, GORDON W, et al. Augmenting RatSLAM using FAB-MAP-based visual data association[C]// Curran Associates, 2013:2–4.
[38]许曈, 凌有铸, 陈孟元, 等. 基于姿态测量模块和闭环检测算法的仿生SLAM研究[J]. 传感技术学报, 2017,30(6): 916–921.XU Tong, LING Youzhu, CHEN Mengyuan, et al. Bio-inspired SLAM based on gesture measuring and closed-loop detection[J]. Chinese journal of sensors and actuators,2017, 30(6): 916–921.
[39]BERKVENS R, WEYN M, PEREMANS H. Asynchronous, electromagnetic sensor fusion in RatSLAM[C]//Proceedings of 2015 IEEE SENSORS. Busan, South Korea,2015: 1–4.
