喜树光合速率日变化及叶片中叶绿素含量季节性变化研究
2018-02-24刘波杨梓桐刘曦子苏禄晖朱丽红傅颖杨意
刘波 杨梓桐 刘曦子 苏禄晖 朱丽红 傅颖 杨意

摘要:指出了喜树为我国特有抗癌植物。对喜树光合速率(Pn)日变化规律、光合速率与光合有效辐射的关系以及喜树嫩叶和成熟叶中叶绿素含量的季节变化进行了研究。结果表明:晴天条件下,喜树光合速率(Pn)日变化规律为双峰曲線,无典型“午休”特征,喜树光合速率随光合有效辐射上升到一定程度后,其光合速率将不随光合有效辐射强度的增大而增加;在整个生长期内,喜树嫩叶的叶绿素含量均低于相应的成熟叶的叶绿素含量,嫩叶与成熟叶的叶绿素含量变化趋势不尽相同。嫩叶叶绿素含量变化为单峰曲线,7月上旬含量达到最高值;成熟叶的叶绿素含量呈双峰曲线变化,6月下旬达到最大值,8月下旬又出现第二高峰。
关键词:喜树;光合速率;日变化;叶绿素;含量;季节变化
中图分类号:S79
文献标识码:A
文章编号:1674-9944(2018)03-0056-04
1 引言
喜树(CamptothecaacuminataDecne.)是我国特有的珙桐科(Nyssaceae)喜树属(CamptothecaDecne.)高大落叶乔木,主要分布于我国长江流域、西南各省区及印度部分地区,属国家二级保护树种[1.2]。喜树根、茎、叶等各部分器官中提取的喜树碱( Camptothecin,CPT)是迄今为止发现的唯一一种拓扑异构酶I( Topoisomer-ase I)的抑制剂,具有广谱抗癌活性,可用于胃癌、直肠癌、慢性粒细胞性白血病和急性淋巴性白血病的治疗[3-5]。进入20世纪90年代后,美国、日本和英国等国家先后开展了对喜树的研究和开发,喜树成为继红豆杉之后第二个重要的木本抗癌药用植物,成为世界性热门研究课题。目前喜树碱源于喜树各组织中分离获得[6]。
近年来,国内对喜树和喜树碱的研究取得了一定的成果[7-15],但对喜树生理特性如光合特性、蒸腾特性和叶绿素含量季节性变化等方面的研究较少[16]。本研究对药用植物喜树光合特性及叶绿素含量季节性变化进行了探讨,以期为药用喜树栽培提供参考。
2 材料与方法
2.1 材料
材料为一年生喜树,种源为四川。以植株成熟叶作(枝条中下部叶,叶片深绿色)为试验材料。
2017年4~11月,于每月中旬、月底的采集喜树嫩叶和成熟叶。采集时间均为早上9:00。
2.2 仪器与试剂
仪器:岛津UV1780型紫外一可见分光光度计(日本岛津公司)、TG328A型电子天平(上海天平仪器厂)、研钵(天津玻璃仪器厂)、玻璃漏斗(天津玻璃仪器厂)。
试剂:80%丙酮、石英砂、碳酸钙粉、蒸馏水。
2.3 喜树光合特性测定
2.3.1 喜树光合速率日变化测定
2017年8月,选择一晴天,用L1-6400便携式光合作用测定仪进行喜树叶片光合速率(μmol CO2.m-2s-l)日变化测定。测定时间为早6:00~18:00,每隔1h测定1次。
在植株中部选择3片成熟叶进行测定,每片叶重复测定3次。
2.3.2 光一光合曲线的测定
用LI- COR公司的LED人T光源,使有效辐射从(Oμmol photons·m-2 s-1)逐渐增加至最大光有效辐射,同时测定相应净光合速率,绘制光一光合曲线,计算光补偿点(O μmolphotons·m-2 s-1)、光饱和点(O μmolphotons·m-2 s-1),根据在低于150~200μmol photons·m-2 s-1光合有效辐射下的光一光合曲线的初始斜率,计算表观量子效率(0 μmol CO2mol-1·Pho-tons)[19]。
2.4 叶绿素含量测定
按Aronon(1949)法测定。称取去脉叶片0.5g,洗净并吸干,剪成1cm的方块,分别置于预冷的研钵中,加入预冷的2mL80%丙酮和少量碳酸钙及适量石英砂,避光快速研磨成均浆,过滤,将滤液转入25mL容量瓶中,用80%丙酮定容至刻度,摇匀,静置20min,取叶绿素提取液,以80%丙酮为对照,岛津UV1780型紫外可见分光光度计在波长为665nm和645nm下测试吸收度。叶绿素a、叶绿素b和总叶绿素含量(mg/LFW)。
叶绿素a含量Ca(mg/L FW)=12.5/1000x10/2.5x1/0.5(12.7D665-2.69D645)
=0.1×(12.7D665-2.69D645)
叶绿素b含量Cb(mg/L FW)=0.1×(22.9D645-4.68D665)
总叶绿素含量CT(mg/L FW)=0.1×(20.21D645+8.02D665)
式中,D665:叶绿素溶液在波长为665nm时的光密度;D645:叶绿素溶液在波长为645nm时的光密度。
3 结果与分析
3.1 喜树叶片光合速率日变化
晴天喜树光合速率( Pn)日变化如图1所示。晴天时Pn日变化为不对称的双峰曲线,最大值出现在上午8:00前后,为11.3102μmolCO2·m-2 s-l。此后Pn逐渐下降,到10:00谷低值,为8.6410 μmolCO2·m-2s-1,到下行14:00 Pn出现第二次高峰,为10.1214μmolCO2·m-2s-l,升高幅度为1.4804μmolCO2·m-2s-1,保持短时间的高峰后Pn急剧下降,到下午18:00趋近于零。比较两个峰值发现,下午的峰值较上午峰值低。一天中的极差达13.8702μmol CO2·m-2s-1。
光合作用的日变化是外界环境条件相互用用以及环境条件与植物本身因素相互制约的综合表现[17]。喜树光合速率最高点出现在上午8:00左右,主要是此时其光合作用随光强增强而加强。一方面光强增加,温度亦逐渐升高,在清晨,C02浓度亦相对较高;另一方面,光照使叶片的气孔开放,有利于C02经气孔进入叶组织内部。这些均利于光合作用进行。但随着光强和温度的继续升高,空气相对湿度降低,叶片内外水汽压差增大,植物蒸腾急升高,进而影响光合作用[18]。另外,失水也使光合产物运输的速度减弱.因而抑制光合作用[17]。下午光合速率出现回升,可能是因太阳西斜,光强和温度均降低,且气孔关闭进而叶片蒸腾降低,缓和了水分消耗和吸收的矛盾,使植物水分状况有所改善的结果[18]。
3.2 喜树叶片光一光合曲线
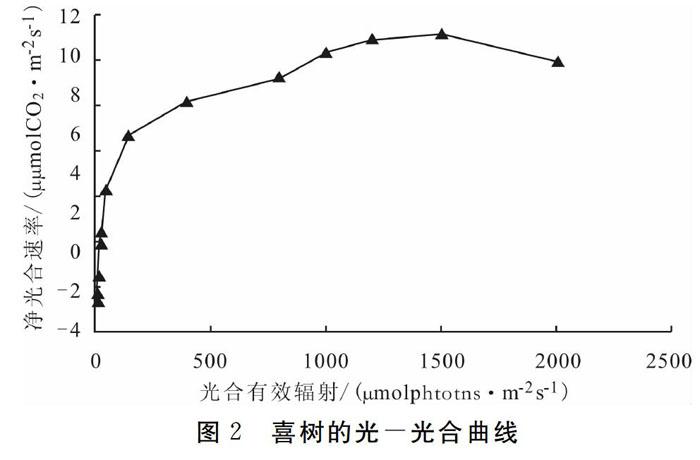
杜占池等[19]对草原植物和作用的光合速率与光合有效辐射的关系进行了研究,认为光一光合曲红可近似用双曲线形式表示。对喜树的光合速率与光合有效辐射进行拟合亦呈双日曲线(图2)。根据光补偿点、半饱和点及饱和點,可把光一光合曲线划分为二个阶段:由光补偿点至半饱和点,是光合速率随光合有效辐射增强迅速增大阶段;半饱和点至饱和点,光合速率随光合有效辐射强度增强逐渐减弱。这二个阶段反映了喜树对光的利用效率。喜树的光补偿点较低,为31.68 μmolphotons·m-2s-l,光饱和点亦较低,为791.93 μmolphotons·m-2s-l,光饱和速率较高,为9.76μmol C02·m-2 s-l,表观量子效率较高,说明喜树利用弱光能力较强,利用强光能力较差,光合的能力较强。
3.3 喜树叶片中叶绿素含量的季节性变化
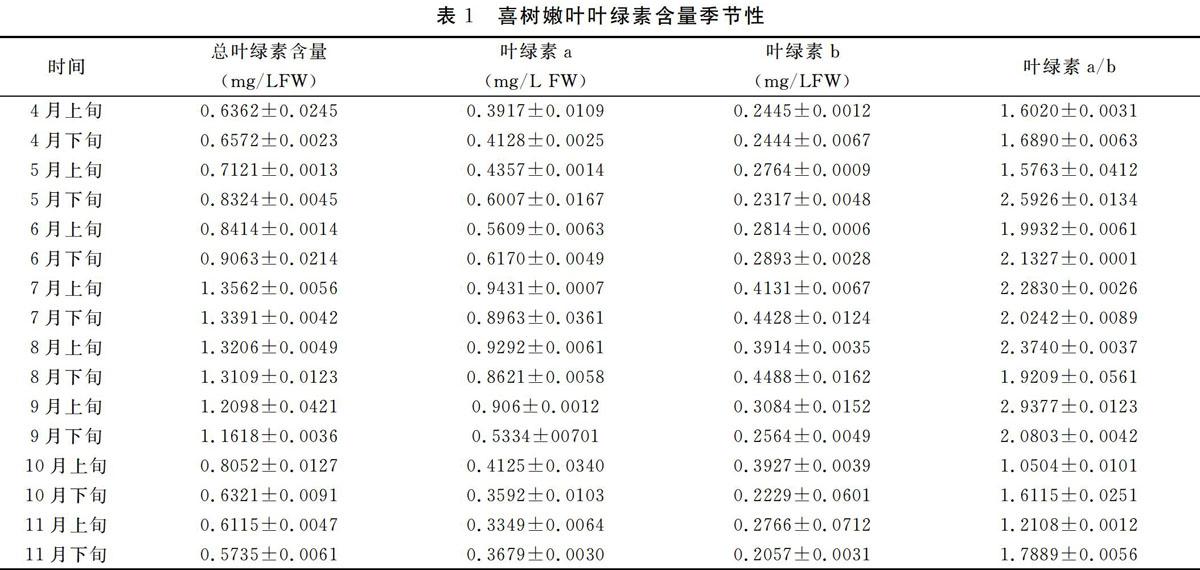
由表1~2可以看出,在整个生长期内,喜树嫩叶的叶绿素含量均低于相应的成熟叶的叶绿素含量,嫩叶与成熟叶的叶绿素含量变化趋势不尽相同。嫩叶叶绿素含量变化为单峰曲线,从4月份开始.嫩叶的总叶绿素含量呈增加趋势,7月上旬达到高大值,为1.3562mg/L FW,后含量逐渐降低,11月底含量达到最低,为0.5735 mg/L FW,且含量低于4月上旬嫩叶叶绿素含量;成熟叶的叶绿素含量呈双峰曲线变化,从4月份开始,叶绿素含量迅速升高,6月下旬达到最大值,达1.9248mg/L FW,后含量逐渐降低,8月下旬出现第二高峰,含量为1.8254 mg/L FW,后叶绿素含量逐渐下降,11月底含量达到最低,为0.6012 mg/L FW,且含量低于4月上旬成熟叶的绿素含量。
由方差分析可知,喜树嫩叶间叶绿素含量差异不显著,成熟叶间叶绿素含量差异不显著,但嫩叶与成熟叶间的叶绿素含量差异极显著((P<0.01)。
3 结论与讨论
植物光合速率日变化及其影响因素是从事植物生理研究的焦点之一,在草本植物方面的研究较多,在多种经济树种的研究亦有报道[20-24]。郑丕尧[25]将植物的晴天Pn日变化分为单峰型、双峰型、严重型和平坦型四种类型。本研究中,在晴天时喜树Pn日变化为双峰曲线,属严重型,不具有典型的“午休”特征。喜树光合速率随光合有效辐射上升到一定程度后,其光合速率将不随光合有效辐射强度的增大而增加。
在整个生长期内,喜树嫩叶的叶绿素含量均低于相应的成熟叶的叶绿素含量,嫩叶与成熟叶的叶绿素含量变化趋势不尽相同。嫩叶叶绿素含量变化为单峰曲线,7月上旬含量达到最高值;成熟叶叶绿素含量呈双峰曲线变化,6月下旬达到最大值,8月下旬又出现第二高峰。
参考文献:
[1]曹淑燕,李映珍.喜树生物学特性与栽培技术[J].广州林业科技, 2000,23(1) :118~120.
[2]中国科学院中国植物编辑委员会.中国植物志[M].北京:科学出 版社,1983 :144~147.
[3]Wall M E,Wani M C,Cook C E,Palmer K H,McPhail A T,Sim G A. Plant antitumor agents.I.Theisolation and structure of camptothecin,a novel alkaloidalleukemia and tumor inhibitor from Camptothecaacuminata[J].J Amer. Chem. Soc., 1966, 88(16):3888-3890.
[4] Hisang Y H, Herizberg R, Hecht S, Liu L F.Camptothecin induces protein-linked DNA Breaks via mammalian DNA topoisomerase I[J].J bioal Chen,1985, (27):4873~14878.
[5]张彦波,张晓婕,肖 磊,等.产生喜树碱及提高其含量的研究现状 与展望[J].河北农业科学,2015,19(5):63~67.
[6] Sriram D, Yogeeswari P, Thirumurugan R, Bal T R.Camptothecin and its analogues:a review on theirchemotherapeutic potential[J].Nat. Prod. Res.,2005,19(4):393~412.
[7]朱 丹.喜树的特征特性及主要繁殖技术[J].上海学业科技, 2015,(3):95~96,78.
[8]王 洋,戴绍军,阎秀峰.光强对喜树幼苗叶片次生代谢产物喜树 碱的影响[J].生态学报,2004,24(6):1118~1122.
[9]王玲丽,刘文哲.不同种源喜树幼枝中喜树碱的含量[J].植物学通 报,2005,22(5):584-589.
[10]史伟国,祖元刚,赵春建,等.匀浆法提取喜树果和喜树叶中喜树碱的研究[J].林产化学与工业,2009,29(1):79~82.
[11]黄瑞松,叶云锋,梁启成,等.HPLC法测定不同产地喜树果中喜树碱[J].中草药,2009,40(1):137~140.
[12]周 涛,江维克,梅 旋,等.贵州喜树的分布特征及群落结构[J].贵州农业科学,2010,38(10):157~159.
[13]刘丽杰,于景华.当年生喜树不同发育阶段叶中喜树碱和10 -羟基喜树碱含量变化[J].齐齐哈尔大学学报,2009(4):63~67.
[14]史伟国,祖元刚,杨磊,等.聚酰胺分离纯化喜树果中10 -羟基喜树碱和喜果苷的研究[J].中国中药杂志,2008,33(21);2486~2489.
[15]向极钎,江喜敏,李红英,等.二年生喜树各器官中喜树碱含量动态分析[J].湖北农业科学,2009,48(12):3141~3142.
[16]冯建灿,张玉洁.喜树光合速率日变化及影响因子的研究[J].林业科学,2007,38(7):34-39.
[17]杨学荣.植物生理学[M].北京:人民教育出版社,1981.
[18]王沙生,.植物生理学(第二版)[M].长春:东北师范大学出版社,1991.
[19]杜占池,杨宗贵.十种草原植物光合速率与光照关系[J].生态学报,1988,8(4):319~323.
[20]苗德全,李淑美,刘 欣,等.田间茌梨叶片光合速率的日变化[J].植物行政管理学,通讯,1991,27(4):273~275.
[21]陶 俊,陈 鹏,佘旭东.银杏光合特性的研究[J].园艺学报,1999,26(3):157~160.
[22]吴大付,张胜利,李东方,等.金银花净光合速率日变化及其对光合有效辐射影响的数学模型[J].安徽农业科学,2009,37 (15):6977~6978.
[23]唐 微,朱名安,刘 俊.C3、C4及CAM植物的光合速率日变化及叶绿素含量的比较[J].湖北农业科学,2002,14:39~40.
[24]彭 立,杨振乾,刘敏敏,等.大气污染物与绿化植物光合速率的关系研究[J].生态环境学报,2015(7):1166~1170.
[25]郑丕尧.作物生理学导论[M].北京:北京农业大学出版社,1992.
