油松天然群体同工酶的变异性研究
2018-01-17王建义贺义才朱松林马佳琳梁书平马彦广
王建义,贺义才,朱松林,马佳琳,梁书平,马彦广
(1.山西省林业科学研究院,山西 太原 030012;2.山西省吕梁山国有林管理局上庄油松种子园,山西 隰县 041300)
油松(Pinustabulaeformis)具有抗寒、耐旱、耐瘠薄等特点,是我国北方重要的乡土树种和造林绿化树种。在油松天然林分中,经过长期的自然选择形成不同的自然类型,个体间差异明显,具有很大的遗传改良潜力,国外已经对油松11种酶系统作过研究[1],从酶谱种类、等位基因频率及杂合率来看,在酶水平上是一个变异丰富的树种[2~4]。为此,研究和评价油松遗传多样性,对于油松育种理论和实践都具有重要意义[5,6]。上世纪90年代,我国开始利用同工酶分析油松群体的遗传结构和变化趋势,直接在基因位点上分析油松天然群体的遗传变异,评价和探讨良种的遗传多样性的[7~9]。本文采用形态结合同工酶技术,分析山西省太岳山国有林管理局灵空山林场的5个天然群体的遗传多样性、等位基因酶的遗传变异以及群体变异与地理因素之间的关系,为山西省油松种质资源的开发利用研究提供科学依据[10,11]。
1 材料和方法
1.1 试验材料
灵空山林场位于山西省太岳山腹地,介于东经110°01′46″~112°08′12″,北纬36°33′52″~36°40′之间,海拔1 200~1 961 m之间,土壤为黄土质棕壤和山地褐土。灵空山区属于暖温带大陆性季风气候,冬季少雪干旱,春季多风干燥,夏季雨量集中,年均气温6.2 ℃,年均降雨量662 mm,相对湿度69.8%~74.3%。灵空山区主要树种有油松、辽东栎、白桦、山杨等,其中油松面积约2 493 hm2。试验材料采自灵空山林场牛槽沟、五龙川、麻刺背、刁王坪、陈家沟等5块生长良好,林相整齐,无病虫害的油松林,在每个林分中选择30株油松样木,株距约30 m。油松林分情况见表1。
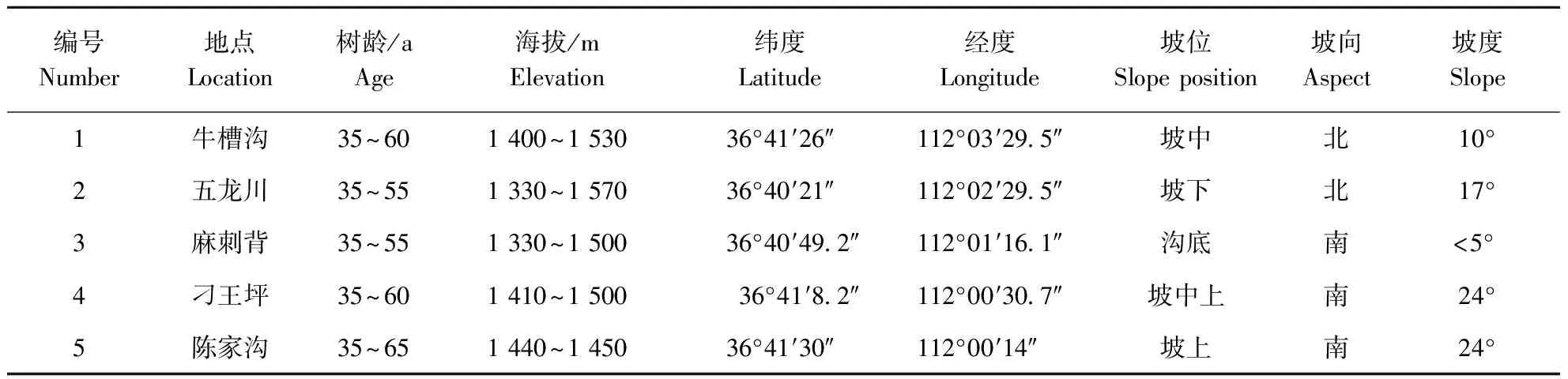
表1 油松林分情况Table 1 Distribution of Pinus tabulaeformis forest
1.2 试验方法
1.2.1 形态特征
针叶采自树冠上部1/3处,每株取针叶33束,测量其长度值。球果采自树冠中上部,在东南西北四个方向各采5个球果,每株20个球果,测量球果长、宽和种子百粒重。
1.2.2 同工酶试验
采用垂直板聚丙烯酰胺凝胶电泳法。分离胶浓度为10%,浓缩胶浓度为4%。电泳3 h后,待溴酚兰指示剂移至液面1 cm 时,停止电泳,剥胶染色。
同工酶位点的确定是由不同的位点由连字符后的数字代表,从离正极最近的位点依次用1,2,3,… 表示。同一位点等位基因根据谱带的迁移率,依次用A,B,C…表示。SA表示无活性或活性极弱的谱带[12,13]。
1.2.3 数据处理
利用软件 Pop Gen 32和 SPSS 22.0进行数据分析,统计出多态位点百分率、等位基因平均数、等位基因频率、期望杂合度、基因分化系数、和遗传距离。
a.多态位点百分率(P)多态位点百分率是同工酶测定多态的位点占总位点数的百分比。
b.等位基因平均数(A)
式中:ai为第i个位点的等位基因数;n为同功酶测定的位点总数。
c.等位基因频率(fij)
式中:Nij为基因的个数;N为基因的总数。
4)王利溥[16]在研究地形对气温和降水的影响时指出:大气水汽含量随海拔增加而减少、地形起伏造成的局地环流能促进上升运动和地形对气流的动力抬升作用对降水影响显著;地形对气温的影响主要体现在地形对辐射分布和对气流方向、速度的改变。关于山地地形风的分布特点,傅抱璞[17]指出,在起伏地形中,由于受局地环流和地形障碍的影响,即使在非常邻近的地方,其风向也可以很不相同。因此,为了能够进一步探讨地形对气温、降水和风速等要素的影响,更加精细、更多样的地形特征信息的应用显得愈发重要,这将在今后的研究中做深入探讨。
d.平均期望杂合度(He)
式中:hei为第i个位点期望杂合度;h为同功酶测定的位点总数。
e.基因分化系数(GST)
式中:Ht为总群体的基因多样度;Hs为群体内基因多样度。
GST反映了群体间变异占总变异的比值大小,是度量群体分化的理想指标。
f.遗传距离(D)

式中:JX为在X群体随机选出的基因相同概率的平均数;JY为在Y群体随机选出的基因相同概率的平均数;JXY为在X,Y两个群体随机选出的两个基因相同概率的平均数;fXij=X群体第i个位点第j个等位基因的频率;fYij=Y群体第i个位点第j个等位基因的频率。
2 结果与分析
2.1 群体间形态特征值变异分析
油松群体、群体内单株及各性状群体特征值列于表2和表3。不同群体间针叶长变幅为7.5~13.6 cm;球果长变幅4.2~7.6 cm;球果宽变幅2.8~4.8 cm;百粒重变幅3.43~6.4 g。百粒重的变异系数明显大于针叶长、球果长和球果宽的变异系数。因此,我们推测百粒重可能受立地条件影响较大。各性状方差分析结果表明,群体间和群体内均未达显著水平[14]。
表2 5个油松林分各性状特征值
Table2 Characteristic values of characters in 5 pine forests
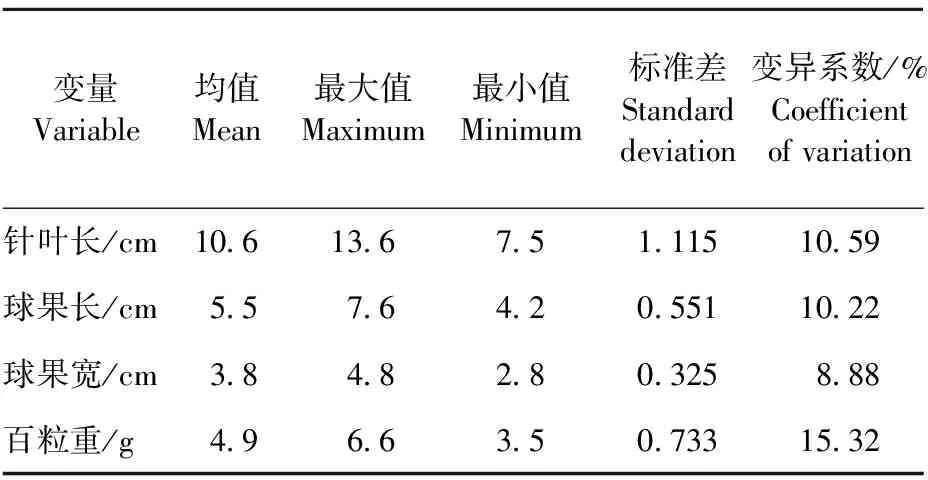
变量Variable均值Mean最大值Maximum最小值Minimum标准差Standarddeviation变异系数/%Coefficientofvariation针叶长/cm10 613 67 51 11510 59球果长/cm5 57 64 20 55110 22球果宽/cm3 84 82 80 3258 88百粒重/g4 96 63 50 73315 32

表3 各性状方差分析表Table 3 Analysis of variance of each character
2.2 同工酶分析
2.2.1 酶谱分析
GOT在油松单株中均有GOT-1和GOT-2两个染色区。GOT-1有慢带和快带2种酶带形式,GOT-2有3种酶谱形式。GOT重复性好,在其位点上存在差异;ADH和SDH在油松单株中均有1个染色区,均受1个位点控制,有3种谱带形式。MDH在油松单株中有2个染色区,受2个位点控制,MDH-1 和MDH-2分别有3种和4种谱带形式。ME在油松单株中均显示1条清晰的谱带,有1个染色区,受1个位点控制,属单态位点。
2.2.2 群体基因变异水平

图1 GOT酶谱Fig.1 GOT zymogram
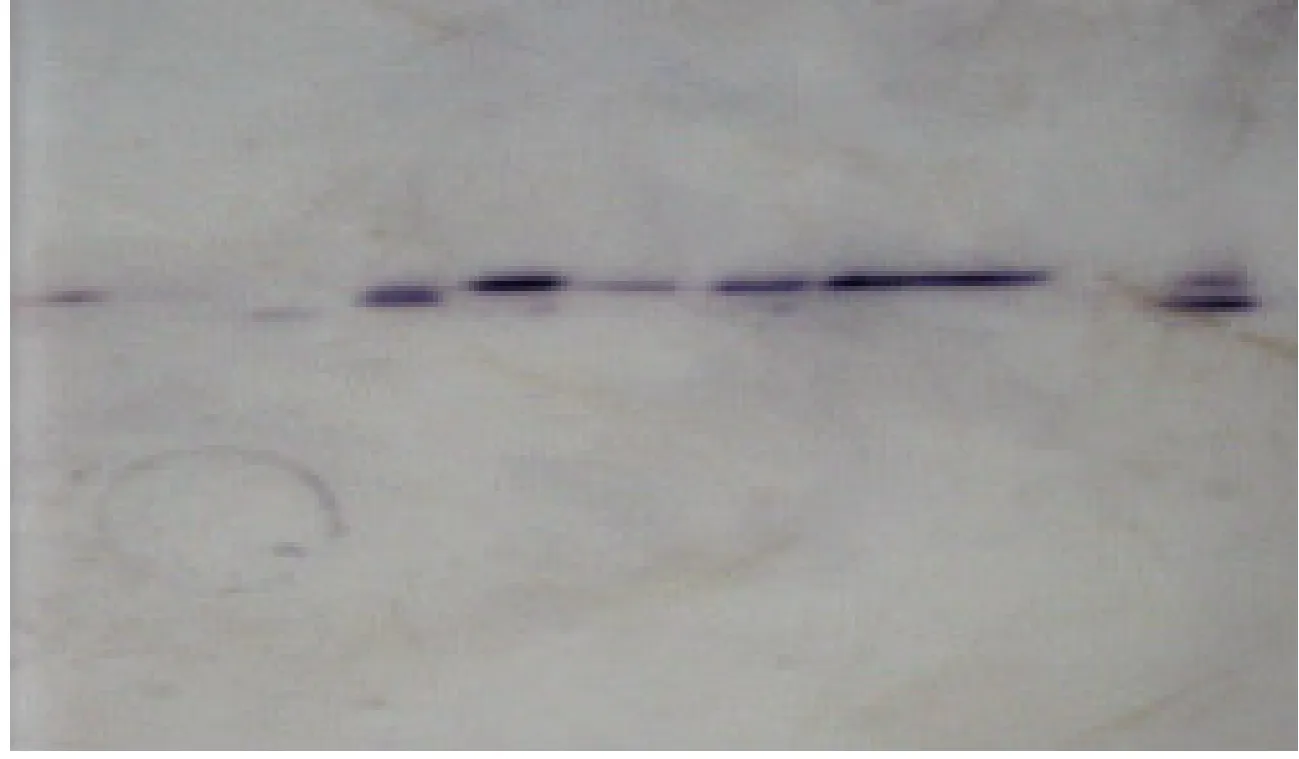
图2 ADH酶谱Fig.2 ADH zymogram

图3 SDH酶谱Fig.3 SDH zymogram
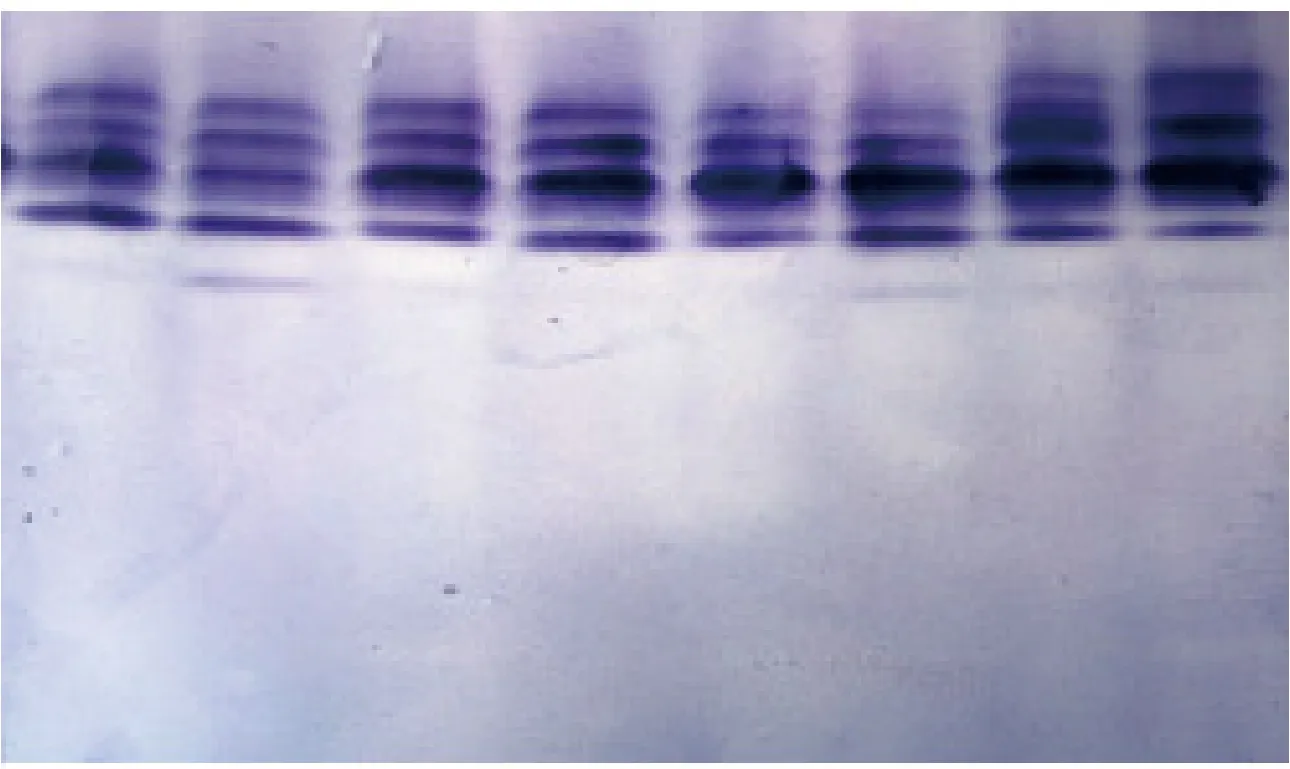
图4 MDH酶谱Fig.4 MDH zymogram
由表4可见,所有油松群体中有Got-1、Got-2、Adh-1、Sdh-1、Mdh-1、Mdh-2等6个位点是多态的。只有Me-1位点是单态的。各位点在所有油松林分中的He值按大小依次为Sdh-1(0.401)、Adh-1(0.243)、Mdh-1(0.236)、Mdh-2(0.232)、Got-1(0.196)、Got-2(0.125)和Me-1。由于He平均值在位点间变化较大,推测不同位点间有较大的异质性[15]。
油松群体中多态位点百分率相同,平均期望杂合度变化范围为0.154~0.267,平均为0.210。等位基因平均数变化范围为2.02~2.48,平均为2.20(表5)。说明群体内等位基因的丰富度一般,群体间变异小,遗传多态参数的均值小,群体在酶位点上变异水平低[16]。
2.2.3 群体间的遗传分化
过对各个天然群体变异进行基因多样度分析,得出其同工酶位点上的变异总量,以及在群体间和群体内的分布情况,由表6可见,Gst值在位点间变化范围为0.006 4~0.071 4,变化幅度较大。7个位点的Gst平均值为0.045 7,表明群体间的变异量平均占总群体的4.57%,95.43%的变异发生在群体内。
2.2.4 群体间遗传距离
分析各个群体间的Nei’s遗传距离D(表7),刁王坪与陈家沟遗传距离为0.984 7,牛槽沟分别与刁王坪、陈家沟的遗传距离是0.986 2、0.994 6。表明群体间遗传距离变动范围非常接近,群体间分化程度并不大。
2.2.5 同工酶变异与海拔的关系
结合各群体的遗传变异水平和海拔高度可知,刁王坪、陈家沟及牛槽沟海拔高度相近,变化幅度小,群体变异相似。五龙川与麻刺背的群体变异相似。表明油松群体的同工酶变异水平与海拔高度变化有关,即海拔高度变化大,群体变异就大;海拔高度变化小,群体变异就小。
3 结论与讨论
3.1 油松群体间的分化
从5个油松优良林分表现性状和同功酶分析看出,油松天然群体的遗传距离与海拔间无明显的相关关系。群体间差异很小,是相对稳定的,具有可遗传稳定性。由此也可推断出灵空山优良种源的性状表现也具有可遗传稳定性[17~19]。从遗传距离分化看,林分间的变异量平均占总群体的的4.50%,而大多数的变异(95.50%)存在于林分内的个体。说明个体差异还是非常明显的。
3.2 油松群体间遗传距离
对群体的遗传一致度(I) 和遗传距离(D) 进行分析发现,遗传一致度(I) 较高,刁王坪与陈家沟遗传距离为0.984 7,牛槽沟分别与刁王坪、陈家沟的遗传距离是0.986 2、0.994 6,这表明群体间遗传距离非常接近(图5)。由此可见,群体间的遗传距离较近与它们分布区之间的地理距离较近有一定关系,油松是高寿命的风媒受粉树木,较近的地理距离使它们之间的基因交流成为可能,再经过多世代的遗传选择使群体间的遗传趋于一致[20~22]。

表4 各群体等位基因频率及期望杂合度Table 4 allele frequency and expected heterozygosity of each population
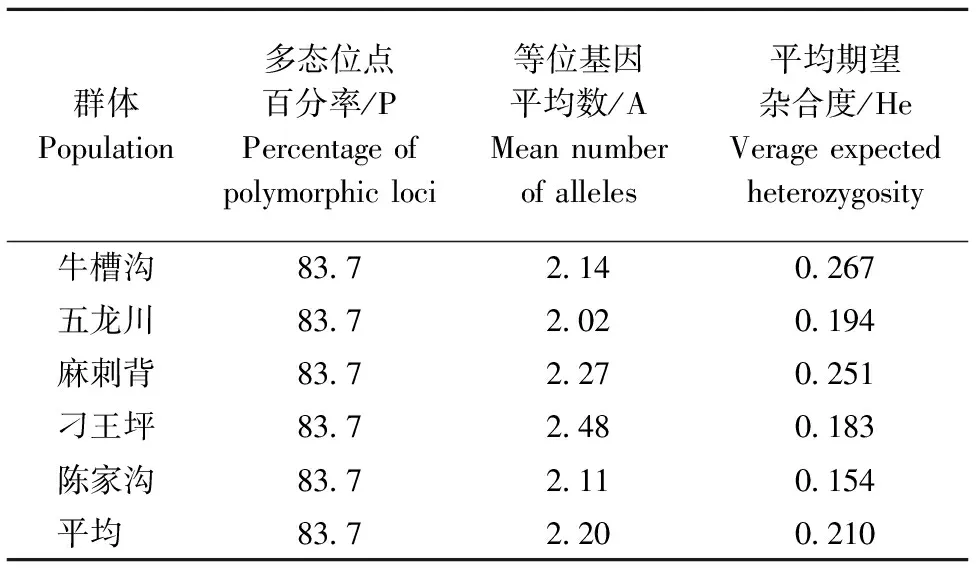
表5 各群体的遗传参数Table 5 Genetic parameters of each population

表6 遗传分化参数值表Table 6 Genetic differentiation parameter values

表7 各群体间的遗传距离Table 7 Genetic distances among populations
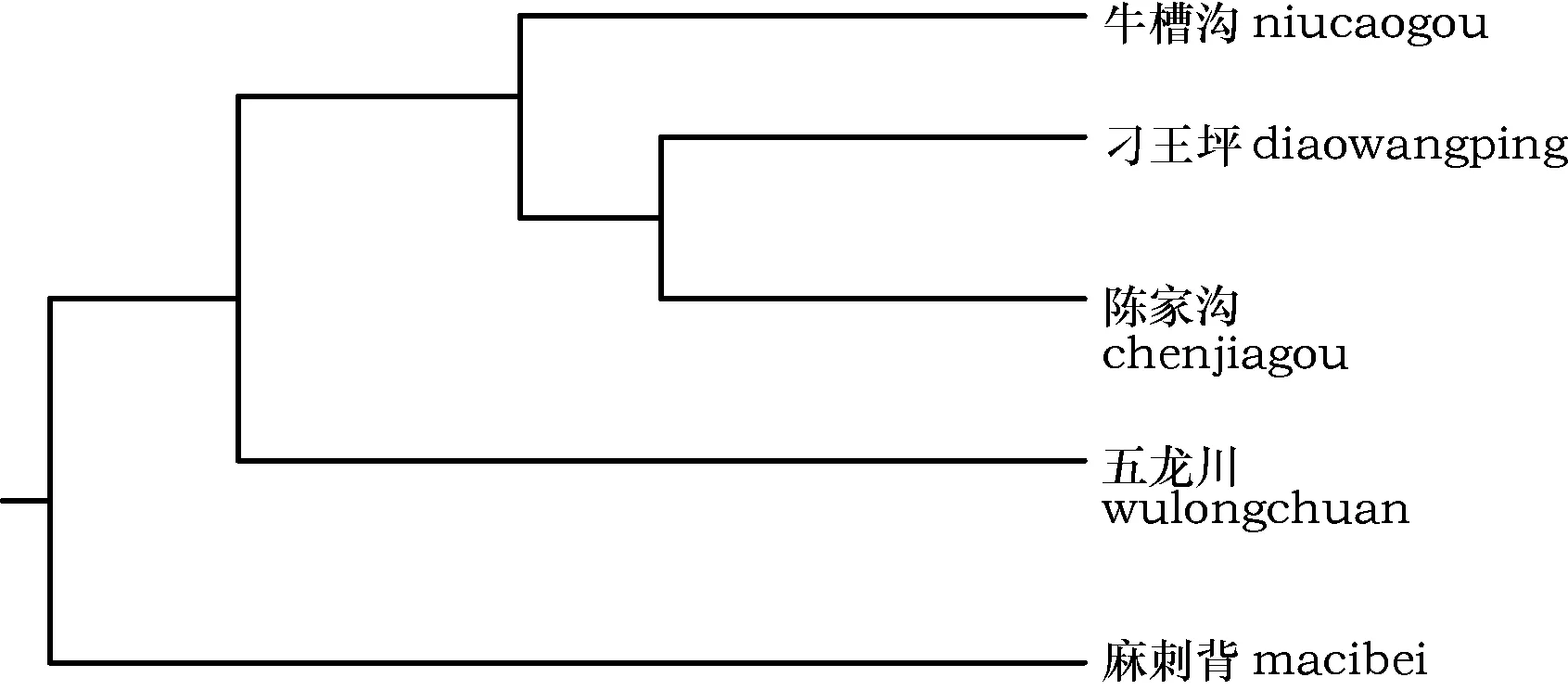
图5 各群体间遗传距离系统图Fig.5 Genetic distance map between populations
[1] 王中仁.植物等位酶分析[M].北京:科学出版社,1996:1-6.
[2] 黄启强,王莲辉.马尾松天然群体同工酶遗传变异[J].遗传学报,1995(2):142-151.
[3] 郭军战,张懿藻,张存旭,等.油松天然群体遗传结构的研究[J].西北林学院学报,1996(4):14-18.
[4] 张春晓,李悦,陈雪梅.油松同工酶位点选择研究[J].北京林业大学学报,1999,21(1):11-16.
[5] Wang X R,Shen X H,Szmidt A E.The Choice of Allozyme Markers for Studies in Conifer Seed Orchards:The Case ofPinustabulaeformisCarr.In:S Fineschi,M E Malvolti,F Cannata and H.H.Hattemer.Biochemical Markers in the Population Genetics of Forest Trees[J].Academic Publishing,The Hague,Netherlands,1991:173-181.
[6] Wang X R,Szmidt A E.Hybridization and chloroplast DNA variation in a Pinus species complex from Asia[J].Evolution,1994,48(4):1020-1031.
[7] 罗建勋,顾万春.云杉表型与同工酶遗传多样性研究进展[J].林业科学研究,2004,17(2):255-262.
[8] 邹春静,盛晓峰,韩文卿,等.同工酶分析技术及其在植物研究中的应用[J].生态学杂志,2003,22(6):63-69.
[9] 张春晓,李悦,沈熙环.林木同工酶遗传多样性研究进展[J].北京林业大学学报,1998,20(3):58-66.
[10] 王意龙,李毳,柴宝峰.山西高原天然油松群体过氧化物酶和多酚氧化酶分析[J].生态环境,2007,16(2):530-532.
[11] 张冬梅,张华新,沈熙环,等.油松种子园交配系统的时空变化研究[J].林业科学,2004,40(1):70-77.
[12] 丁坤善,郑彩霞,包仁艳.油松雌性不育系的POD同工酶和蛋白质多肽分析[J].西北植物学报,2004,24(1):17-20.
[13] 柴国锋,郑勇奇,王良桂,等.枫香同工酶遗传多样性分析[J].林业科学研究,2013,26(1):15-20.
[14] 毕春侠,郭军战,张懿藻.油松天然群体酯酶同工酶的变异分析[J].西北林学院学报,1998,13(4):39-43.
[15] 许玉兰,蔡年辉,陈诗,等.云南松天然群体遗传变异与生态因子的相关性[J].生态学杂志,2016,35(7):1767-1775.
[16] 许玉兰.云南松天然群体遗传变异研究[D].北京:北京林业大学,2015.
[17] 刁松锋,邵文豪,姜景民,等.基于种实性状的无患子天然群体表型多样性研究[J].生态学报,2014,34(6):1451-1460.
[18] 李帅锋,苏建荣,刘万德,等.思茅松天然群体种实表型变异[J].植物生态学报,2013,37(11):998-1009.
[19] 赵罕,郑勇奇,李斌,等.白皮松天然群体遗传结构的地理变异分析[J].植物遗传资源学报,2013,14(3):395-401.
[20] Gonçalves A C,Pommerening A.Spatial dynamics of cone production in Mediterranean climates:A case study of Pinus pinea L.in Portugal[J].Forest Ecology and Management,2012,266:83-93.
[21] Owens J N,Kittirat T,Mahalovich M F.Whitebark pine (PinusalbicaulisEngelm.) seed production in natural stands[J].Forest Ecology and Management,2008,255(3-4):803-809.
[22] Xu Y,Woeste K,Cai N,et al.Variation in needle and cone traits in natural populations of Pinus yunnanensis[J].Journal of forestry research,2016,27(1):41-49.
