特呈岛红海榄林分结构特征分析
2018-01-12周元满谢嘉莉黄剑坚刘志美
周元满,谢嘉莉,黄剑坚,刘志美
(广东海洋大学农学院,广东 湛江 524088)
红树林生长于陆地与海洋交界带的滩涂,是陆地向海洋过度的特殊生态系统,也是热带亚热带海岸的生态关键区,对开展全球保护生物多样性,调节全球气候和发展生态旅游都有极其重要的作用。红海榄作为红树林家族中的重要成员,为灌木或小乔木,支柱根发达,滞潮消浪作用显著,是海滩良好的防护树种。然而,目前我国的红树林保护面临严峻形势,围垦滥伐、化学污染、白色污染、捕杀林鸟等都对我国各地区红树林造成重大的破坏。如何有效地阻止对红树林生态系统的破坏以及恢复红树林生态系统的各种生态功能,已成为急需解决的重大问题。林分结构特征在很大程度上决定了林分的稳定性和经营空间大小[1],同时,林分结构是森林发展过程,如更新方式竞争、自然稀疏和经历的干扰活动的综合反映,是森林经营和研究分析中的一个重要因子[2]。目前有关红树林空间结构的研究主要是从林分的混交度、 大小比数和角尺度等林分空间结构参数以及空间结构均质性入手分析其空间分布特征[3,4], 然而,林木个体之间的空间距离和相互连接程度决定了林木之间的竞争势及其空间生态位,对林木的生长和繁育有重要作用。本文以湛江特呈岛红树林生态系统为对象,通过重要值指数、最小距离指数和连接度指数等方面对天然红海榄的林分结构进行研究,更好地揭示红海榄种群的生态位特征及分布格局,并探求其分布规律的原因,以期为研究红树林群落的演替趋势以及红树林生态系统的恢复与空间优化模型的建立提供理论依据。
1 研究区概况
特呈岛位于湛江市霞山区东南面的湛江港海域。地处110°25′—110°27′ E,21°09′—21°10′ N,全岛南北宽1.4 km,东西长2.7 km,海岸线长7.4 km,岛上陆地面积约3.6 km2[3]。特呈岛属北热带海洋性季风气候,长年温暖湿润。年平均气温22.3 ℃,历年1月平均气温最低,7月最高,全年无冬季。年均降雨量1 800~2 000 mm,夏秋盛吹偏南风,常有台风侵袭,并夹带暴雨,风力最大达12级以上。特呈岛四周波浪浪高全年以3级(0.5~1.4 m)为主,潮间带为半日潮,潮高变化-27~469 cm[5]。特呈岛的红树林以白骨壤单优群落及白骨壤和红海榄混交群落为主,群落类型主要有白骨壤群落、白骨壤+红海榄混交群落和红海榄群落等[6]。本文选取的群落类型为:红海榄纯林(Ⅰ),中偏高潮带;红海榄+木榄+白骨壤混交林(Ⅱ),中潮带;红海榄+白骨壤混交林(Ⅲ),中潮带,土壤质地为沙质土。
2 材料与方法
2.1 样地设置与材料收集
本研究于2015年7月至2016年5月在特呈岛的红海榄纯林、红海榄+木榄+白骨壤混交林以及红海榄+白骨壤混交林中采用随机抽样的方法,分别设置10 m×10 m的样地,3次重复。然后在样方内进行每木检尺。记录样方内植株所在的样本号、种名、数量、树高、枝下高、地径、冠幅以及红海榄相邻植株之间的距离。
2.2 研究方法
2.2.1 重要值指数
重要值指数Ⅳ(%)=(相对多度+相对频度+相对优势度)/3
相对多度A(%)=100×某个种的株数/所有种的株数;
相对频度F(%) = 100×某个种在样方中出现的次数/所有种出现的次数;
相对显著度T(%)=100×某个种的优势度/所有种的优势度之和(本文研究树种的显著度指标用基部盖度来表示)[7]。
2.2.2 距离指数 用林分内植株之间的距离来构造的指数称为距离指数。距离指数有两种作用:第一是用来分析群落中植株分布是否服从随机分布,二是用来定量描述植株之间的连接度或隔离度[8]。距离指数分为两种:一种是最小距离指数,另一种是连接度指数。
最小距离指数用来检验群落里一个物种的个体是否服从随机分布。
NNI=MNND/ENND
(1)
式中:NNI为最小距离指数;MNND为每一植株与其最近相邻植株间的平均最小距离;ENND为在假定随机分布前提条件下MNND的期望值[9]。
MNND和ENND计算如下:
(2)

(3)
式中:NNDi为植株i与其最近相邻植株间的最小距离;d为群落中给定植株类型的密度;N为给定植株类型的植株数;NNDi为植株i中心到其最近相邻植株中心的距离。
密度d计算如下:
d=N/A
(4)
式中:A为样地总面积。
若NNI的取值为0,则格局完全团聚分布;若NNI的取值为1,则格局为随机分布;若NNI取其最大值2.149,则格局为完全规则分布[8,9]。
连接度表示群落中同种植株在生态功能与生态过程上相互联系的程度或潜力[10]。连接度指数用来描述种群里同类植株联系程度的指标。连接度指数是最近相邻植株距离的反函数,它使用植株占地面积作加权:
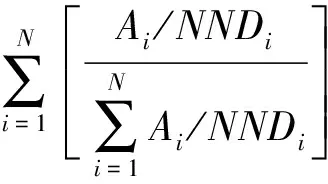
(5)
式中:PX为连接度指数;Ai为植株i的占地面积;NNDi为植株i到其相邻植株的最小距离。
PX取值从0到1。PX取值接近1时,则表明格局是群聚分布;取值接近0时,表明格局是离散分布[9,10]。
3 结果与分析
3.1 不同红海榄群落类型中树种的重要值分析
外业调查结果(表1)表明,在红海榄+木榄+白骨壤混交林(Ⅱ)中,白骨壤在乔木层的重要值指数最大,为46.5%,红海榄在灌木层的重要值指数最大,为66.7%。在红海榄+白骨壤混交林(Ⅲ)中,白骨壤在乔木层的重要值指数最大,为100%,红海榄在灌木层的重要值指数最大,为70.2%。可见,在红海榄的混交林群落类型中,白骨壤在乔木层处于优势地位,而红海榄在灌木层处于优势地位。

表1 不同群落类型中物种重要值指标 %
树种的重要值反映了一个树种在群落中的地位。在不同群落类型灌木层中红海榄的重要值指数均为最大,表明红海榄在灌木层均处于优势地位;而白骨壤在样地Ⅱ和样地Ⅲ中的重要值指数为46.5%和100%,均比木榄和红海榄的重要值指数大。在乔木层中,白骨壤先锋树种主要占据了乔木层,其优势和主导地位主要体现在它的基部盖度(地径大小)。由于乡土树种中能够发展到乔木层的红海榄和木榄数量不多,而绝大部分的个体还是集中分布于灌木层。白骨壤在所调查的红海榄群落中数量不多,呈孤立木状态分布。白骨壤作为特呈岛红树林的先锋树种,其纯林带主要分布在红树林的低潮位外缘地带。由于潮水的涨退,其种子更容易进入红海榄群落林带进行自然更新和树种的迁移。因此,一段时期内先锋树种在乔木层的优势地位都不会改变。在灌木层中,白骨壤的优势地位不再突出,红海榄数量多,密度大,是整个林分灌木层的优势种群,其优势和主导地位主要体现在它的相对多度。红海榄林分分布在特呈岛红树林中潮带,总体位于红树林中心地带,外围有高大的白骨壤作为天然的防护屏障,很大程度上能够削减风浪对红树林内部的侵蚀和破坏。因此,红海榄在中心区域中更新及生长情况良好,一段时期内在相对稳定的生境中优势地位不会被取代。
3.2 红海榄径级结构和高度级结构特征分析
3.2.1 红海榄径级结构特征分析 由表2可知,红海榄、木榄和白骨壤3个树种的树高、冠幅和枝下高相当,而白骨壤的地径平均值远远大于红海榄和木榄。可见,红海榄、木榄和白骨壤生态位有重叠性,其中红海榄和木榄的重叠度高,种间竞争激烈。
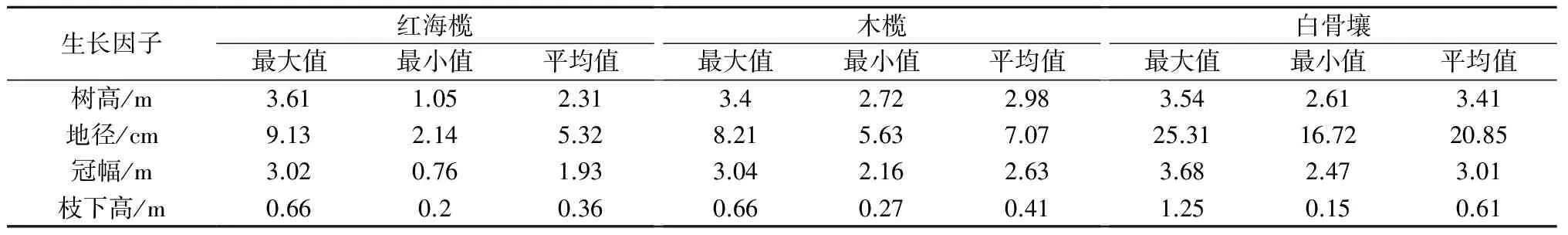
表2 主要树种林木生长特征
将红海榄的地径(DBH)划分为3个等级:DBH<4 cm, 4≤DBH<6 cm,DBH≥6 cm。然后分别统计各个级别的数量、比例和平均值(见表3)。

表3 不同群落类型中红海榄的地径分布结构
表3显示,在红海榄纯林中,红海榄地径大小集中分布在4≤DBH<6 cm与DBH≥6 cm,在红海榄+木榄+白骨壤混交林中,红海榄地径大小集中分布在4 cm≤DBH<6 cm之间,占60.5%,地径平均值比纯林小,表明红海榄在该混交林中,种间竞争对红海榄的径级分布影响较大。在红海榄+白骨壤混交林中,红海榄地径大小分布在4 cm≤DBH<6 cm之间的林木占57.1%。
3.2.2 红海榄树高级结构特征分析 将红海榄的树高划分为两级:灌木层(H<3 m),乔木层(H≥3 m)。由不同群落类型红海榄的树高分布(见表4)可知,红海榄在不同群落类型中灌木层的数量最多,分别占60.9%,97.4%,100.0%。在乔木层中,红海榄纯林的比例最大,为39.1%,平均树高也最大,为2.87 m;红海榄+木榄+白骨壤混交林的比例最小。可见红海榄在与其他红树植物的混交林中其树高生长不及红海榄纯林,表明种间竞争对红海榄树高的生长有着明显影响。

表4 不同群落类型红海榄的树高分布
3.3 红海榄地径与树高的相关性分析
在分析不同群落类型中红海榄径阶结构的基础上,将3种群落类型的调查数据作为一个整体,进一步分析红海榄的径阶结构与树高的相关关系。由表5可见,红海榄林分的地径分布呈现常规的正态分布趋势,依据树高随地径分布的变化趋势,运用符合散点分布趋势的若干相关模型进行模拟,得出各模型的参数及复相关指数(R2)如表6所示。
由表6可见,各拟合方程的复相关指数(R2)均大于0.96,拟合效果较好,其中以二次多项式方程H=a+bD+cD2拟合效果最好,复相关指数(R2)达0.996 6。经回归检验计算得出:F=244>F0.01(1,87)=6.96,表明模型拟合效果极显著。

表5 红海榄各径阶的特征值
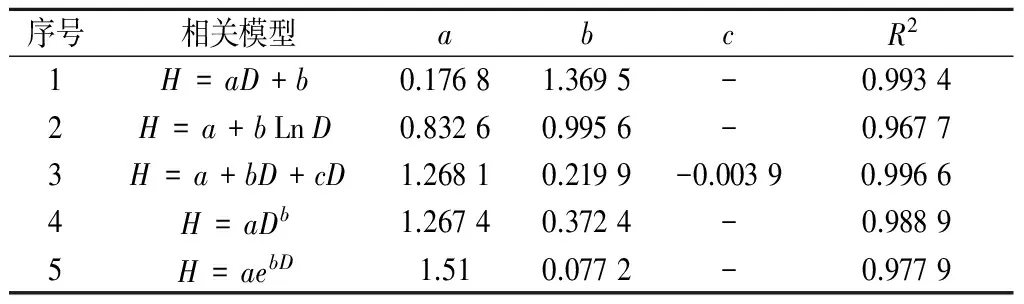
表6 红海榄树高与地径的拟合模型
为进一步验证上述模型的适应性,用参加回归的89株红海榄的地径与树高数据,计算最佳回归方程的系统误差(Se)和各径阶系统偏差(表7),树高与地径的回归方程系统误差,即各径阶系统偏差之和为0.64%,远小于±5%,表明红海榄模型拟合精度极高。

表7 各径阶系统误差
3.4 红海榄林分空间连接与分布规律
3.4.1 最小距离指数分析 根据上述红海榄地径和树高的相关分析表明,红海榄的地径大小集中在4 cm≤DBH<6 cm。因此,以地径为依据,进一步分析不同群落类型中DBH<4 cm,4 cm≤DBH<6 cm,DBH≥6 cm3个级别的红海榄空间连接度。利用外业调查数据计算出林分中植株的最小距离指数。结果见表8。

表8 最小距离指数各参数表
注:A,4 cm≤DBH<6 cm红海榄纯林;B,DBH≥6 cm红海榄纯林;C,DBH<4 cm红海榄+木榄+白骨壤混交林;D,4 cm≤DBH<6 cm红海榄+木榄+白骨壤混交林;E,DBH≥6 cm红海榄+木榄+白骨壤混交林;F,DBH<4 cm红海榄+白骨壤混交林;G,4 cm≤DBH<6 cm红海榄+白骨壤混交林;H,DBH≥6 cm红海榄+白骨壤混交林
由表8可知,除DBH≥6 cm红海榄纯林以外,其他各类型红海榄的最小距离指数均接近于1,而且无论在纯林还是混交林,在4 cm 3.4.2 林分空间连接度分析 在对不同群落红海榄的最小距离指数分析的基础上,根据连接度指数公式计算出各类型林分的连接度指数(图1)。由图1可知,各群落类型中红海榄林分的连接度指数均接近于0,表明红海榄分布趋于离散。其中,类型C,即DBH<4 cm红海榄+木榄+白骨壤混交林红海榄植株群落的连接度指数最小为0.089,分布格局显示为最离散;类型E,即DBH≥6 cm红海榄+木榄+白骨壤混交林红海榄植株群落连接度指数最大为0.378。 在红海榄纯林(Ⅰ)中,连接度指数大小顺序为:0.219(4 cm≤DBH<6 cm)<0.294(DBH≥6 cm)。 图1 红海榄在不同群落类型中空间连接度分布 在红海榄+木榄+白骨壤混交林(Ⅱ)中,连接度指数大小顺序为0.089(DBH<4 cm)<0.158(4 cm≤DBH<6 cm)<0.378(DBH≥6 cm);在红海榄+白骨壤混交林(Ⅲ)中,连接度指数大小顺序为:0.099(DBH≥6 cm)<0.133(4 cm≤DBH<6 cm)<0.137(DBH<4 cm)。分布频数最大的红海榄(4 cm≤DBH<6 cm)在3种群落类型中的连接度指数顺序为:类型Ⅲ(0.133)<类型Ⅱ(0.158)<类型Ⅰ(0.219);DBH≥6 cm红海榄群落,在3类样地的连接度指数顺序为:类型Ⅲ(0.099)<类型Ⅰ(0.294)<类型Ⅱ(0.378);DBH<4 cm红海榄群落,在两类样地的连接度指数顺序为:类型Ⅲ(0.137)<类型Ⅱ(0.089)。可见,在同一群落类型中,地径大小及其连接度指数大小既有正相关也有负相关;而不同群落类型中,地径大小及其连接度指数关系不明显。而林分的组成结构对红海榄空间连接度亦有一定影响,从整体上看,红海榄在木榄和白骨壤混交林中其连接度指数较小,分布格局更趋于离散。这是因为该林分受人为干扰较大,造成红海榄种群分布破碎化程度高,林分空间连接度亦较小。 [1] 惠刚盈,胡艳波.混交林树种空间隔离程度表达方式的研究[J].林业科学研究,2001,14(1):177-181 [2] 雷相东,唐守正.林分结构多样性指标研究综述[J].林业科学,2002,38(3):140-146 [3] 李际平,张宏伟,李建军,等.不同潮带红树林空间结构比较研究[J].中南林业科技大学学报,2010,30(1):8-11 [4] 李建军,李际平,刘素青,等.红树林空间结构均质性指数.林业科学,2010,46(6):6-13 [5] 韩维栋,高秀梅,吴钿. 特呈岛种子植物区系研究[J]. 湖南林业科技,2006,33(6): 23-26 [6] 韩维栋,高秀梅.特呈岛白骨壤古树群的调查分析[J].防护林科技,2008(1):6-8 [7] 李洪旺,王旭,余雪标,等.海南东寨港秋茄+桐花天然红树林群落林分结构特征研究[J].热带林业,2008,36(1):30-33 [8] 李哈滨,伍业刚.景观生态学的数量研究方法[M].北京:中国科学技术出版社,1992:209-234 [9] 邬建国.景观生态学概念与理论[J].生态学杂志,2000,19(1):42-52 [10] 张芸香,郭晋平. 森林景观斑块密度及边缘密度动态研究[J]. 生态学杂志,2001,20(1):18-21
4 结论与讨论
4.1 不同红海榄群落类型中树种的重要值表明红海榄在群落灌木层处于优势地位,并与木榄处于长期竞争状态,白骨壤在乔木层处于优势地位。
4.2 红海榄地径与树高的最佳拟合模型为H=1.268 1+0.219 9 D-0.003 9 D2,复相关指数(R2)为0.996 6,拟合效果理想。对于快速准确地估测特呈岛红海榄树高、林分平均高及评价立地质量等提供理论依据。
4.3 由最小距离指数及空间连接度分析结果表明,特呈岛红海榄种群呈随机分布格局。从整体上来看,红海榄纯林的最小距离指数低于混交林,其中红海榄+白骨壤混交林中最小距离指数最高,但仍接近随机分布,这与种间竞争和红海榄这种以母树为中心散播种子的生物特性有关。可见,特呈岛不同群落中红海榄种群可能趋于近熟林,林分结构趋于稳定。
4.4 群落中每个物种都占据相应的生态位,种内竞争使种群的生态位宽度变宽,而种间竞争则使其生态位变窄。特呈岛红树林生态系统中红海榄和木榄主要分布在群落的灌木层,两者生态位宽度值接近,生态位重叠高,种间竞争可能导致其分布格局不断变化;在红海榄纯林中,种内竞争而发生的自疏现象是影响其分布格局的重要因素。红海榄纯林长势较好,平均株高与平均地径均大于其他2种混交林(表3,表4), 随着植物种群发育及年龄的增大,个体对环境条件的要求也相应提高,在阳光、土壤、养分等资源不充足时种内竞争导致种群发生自疏现象,种群的个体数量递减,从而减弱种群分布的聚集程度,这有利于提高种群利用资源的能力,扩大其生态位宽度,从而导致种群的分布格局趋向于随机分布型。
4.5 红海榄空间连接程度低,混交程度越高,分布破碎化程度越高。红海榄种群的空间分布格局除了与生境异质性、自身生物学特性、种内和种间竞争等方面有关,还可能与人为的干扰和动物的啃食有关。目前,特呈岛红树林保护区生态环境日益恶化,环境污染不断加剧,生物多样性也逐年递减,许多植株生长畸形,呈现白化或枯萎状态。在红树林生态系统经营中应尽量避免因围海养殖、乱砍滥伐、港口建设、水质及固体废弃物污染等造成红树林群落分布的破碎离散,促进生态系统的恢复。
