海水胁迫对白子菜生长及生理特性的影响
2018-01-12覃和业徐微风王永壮符少萍郭建春段瑞军
王 蕾, 覃和业, 徐微风, 刘 姣, 王永壮, 符少萍, 郭建春, 段瑞军
(1.中国热带农业科学院/热带生物技术研究所,海南海口 571101;2.海南大学,海南海口 571101)
随着环境条件的恶化,土壤盐渍化逐渐成为一个全球性的生态问题。相关研究表明,盐渍化土壤占全球耕地面积的7%,其中约有20%的耕地正受到次生盐渍化的威胁[1-2]。土壤中过高的盐分易造成植物细胞结构的损伤,严重抑制植物的生长,甚至会导致植物大面积死亡,极大地影响了现代农业的生产及可持续发展[3]。随着全球性粮食、资源、生态环境问题的日趋严重,沿海滩涂植被显示出越来越大的生态及经济效益,海水蔬菜的开发及利用已经成为改良土壤盐渍化、维持生态平衡的重要途径,也是目前应对盐渍化危机的主要方法[4-5]。目前,北美海蓬子、碱蓬、番杏、中亚滨藜等耐盐植物已被报道用于盐渍土的改良[6]。
白子菜[Gynuradivaricata(L.) DC.]别称接骨丹、百子菜、茹童菜、鸡菜、大肥牛白背菜、散血姜、明月草等,属菊科三七属,为多年生草本植物,主要分布于我国华南地区、云南及越南北部等地区[7]。白子菜根系发达,繁殖能力强,可扦插,扦插5~7 d即可成活,适宜在含盐量高的沙地种植[8]。此外,白子菜抗逆性强、耐高温,适宜在海南地区种植。白子菜药用及营养价值较高,嫩茎可食用,味甘、淡,寒,根入药有清热泻火之效[9]。可用于清热解毒、活血通经、舒筋接骨、凉血止血,对支气管肺炎、小儿高热、百日咳、目赤肿痛、风湿关节痛、崩漏、跌打损伤、血瘀肿痛、骨折风湿骨痛、外伤出血痈疮肿毒、乳腺炎、疮疡疔肿、烧烫伤等具有明显的疗效[10]。当前对白子菜的研究多集中在栽培技术、药用价值等方面,关于其在盐胁迫下生长及生理响应的研究国内外尚未有报道。本试验通过海水长期处理白子菜,研究不同海水浓度处理对白子菜的生长及相关生理指标的影响,为进一步利用白子菜对滨海滩涂地、盐荒地、海水倒灌农田的改良和利用提供理论基础。
1 材料与方法
1.1 试验材料
白子菜采自海南省陵水黎族自治县分界洲岛,将嫩茎扦插于塑料花盆中进行育苗,插后4~7 d即能发生不定根,12~15 d后定植于透气的塑料花盆中。海水取自海南省海口市白沙门,盐浓度为3.0%。
1.2 试验方法
1.2.1 材料处理 本试验于2016年3月在中国热带农业科学院热带生物技术研究所内进行,设置6个海水浓度(0、10%、20%、30%、40%、50%),3次重复。当扦插的白子菜长出4~6张新叶后进行移栽,选取长势一致的幼苗定植于塑料花盆中,每盆3株,浇透水。试验时在06:00—7:00间土壤温度较低时进行海水处理,同时配以Hoagland 营养液,每隔5 d浇1次海水,每隔20 d浇1次Hoagland营养液,当表层沙土 1~2 cm发白时及时补浇自来水,使土壤含水量达到田间最大含水量的60%~70%。栽培基质选用筛过的细河沙。90 d后,测定其相关生理形态指标等。
1.2.2 形态指标的测定 各处理分别取白子菜3株,用直尺测量其株高(cm)、节间长(cm),用游标卡尺测定其茎粗(cm)、叶片厚度(cm),用紫光平台扫描仪(LA2000)结合ImageJ软件测定叶面积[11]。
1.2.3 生理指标的测定 超氧化物歧化酶(简称SOD)活性的测定参照李合生等的氮蓝四唑光化还原法[12-13];过氧化物酶(简称POD)活性的测定参照朱广廉等的愈创木酚法[14];过氧化氢酶(简称CAT)活性的测定参照彭志英等的方法[15];脯氨酸含量的测定参照郑炳松的茚三酮法[16]并有改进;可溶性糖含量的测定参照郑炳松的蒽酮比色法[16];细胞膜透性参照刘宁等的方法[17-18]测定质膜相对透性;丙二醛(简称MDA)含量测定参照Heath等的硫代巴比妥酸(简称TBA)比色法[19];根系活力的测定参照张宪政的2,3,5-三苯基氯化四氮唑(TTC)法[20];叶绿素含量的测定采用张宪政的丙酮乙醇混合提取法[20]。
1.2.4 数据分析 采用Excel 2003整理试验数据、绘制图表;用SPSS 13.0进行方差分析和多重比较,进行α=0.05水平的统计显著性检验。
2 结果与分析
2.1 海水胁迫对白子菜生长的影响
为研究海水处理对白子菜生长的影响,本试验测定不同浓度海水处理下白子菜的形态指标(株高、茎粗、节间数、叶片厚度、叶面积、地上部生物量)。由表1可知,白子菜在0~30%海水浓度处理下,各生长指标相较于对照均未受到明显抑制,且在≤20%海水浓度处理下,大多数指标随海水浓度的提高有不同程度的增加;株高、节间数、叶面积在海水浓度≥40%时均显著减少;而地上部生物量在10%~20%海水浓度胁迫下有所增加,当海水浓度≥40%时,地上部生物量显著减少,说明当海水浓度在0~20%时在一定程度上促进了白子菜的生长,当海水浓度≥40%时,白子菜生长受海水抑制作用显著。
2.2 海水胁迫对白子菜细胞膜透性的影响
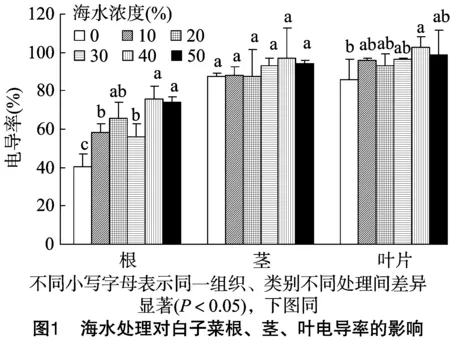
2.2.1 海水胁迫对白子菜细胞膜透性的影响 电导率是反映细胞膜透性的一个非常重要的指标。白子菜在未经海水处理下,其根、茎、叶中的细胞膜透性分别是40.25%、87.50%、85.64%。由图1可知,在海水胁迫下,白子菜根茎叶中膜透性整体随胁迫的增加而升高,但在茎中无显著性升高;在40%海水浓度胁迫下叶片中电导率显著增加,其余各浓度处理下无显著性变化;根系中的变化尤为明显,在≥10%海水处理下,电导率显著增加。说明海水胁迫对根系的损伤远远大于茎、叶。
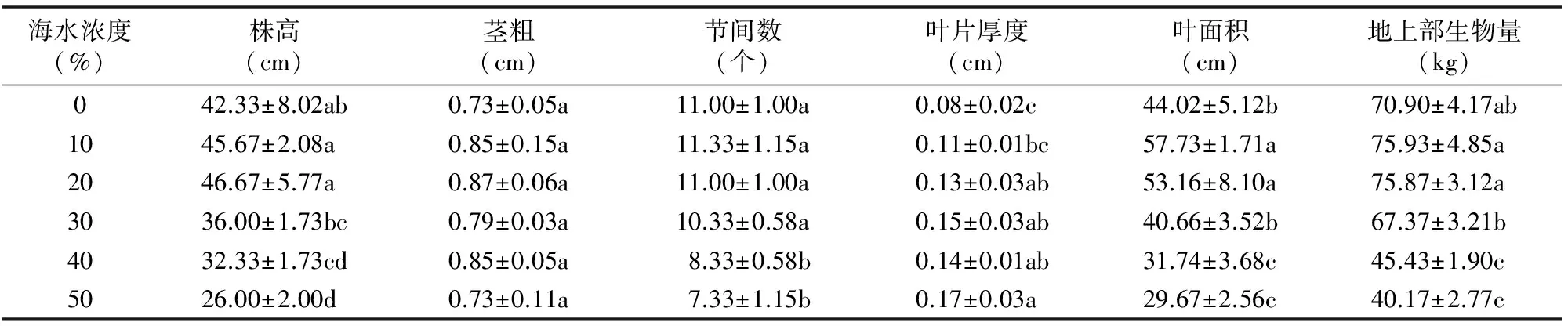
表1 海水处理对白子菜生长指标的影响
注:同列数据后不同小写字母表示差异显著(P<0.05)。
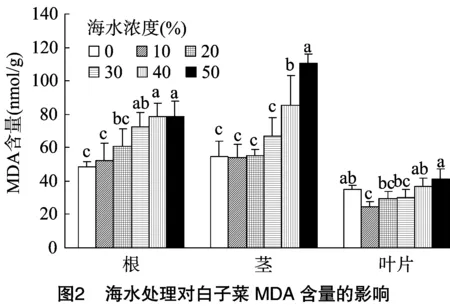
2.2.2 海水胁迫对白子菜MDA含量的影响 MDA是膜脂过氧化作用的主要产物之一,可间接表示细胞膜的受损程度,并兼有反馈的作用。在未经海水处理下,白子菜根、茎、叶片中MDA的含量分别为48.26、54.36、34.70 nmol/g。由图2可知,白子菜MDA含量在根、茎中随海水浓度的增加整体增加,分别在≥30%、≥40%海水处理下MDA含量显著高于对照。在叶片中,MDA含量呈先减少后增加的趋势,当海水处理浓度≤30%时,MDA含量较对照略有下降,但随着海水浓度的升高MDA含量逐步上升,最大值达到41.20 nmol/g,较对照增加了18.7%。说明在低浓度下海水对白子菜细胞膜的损伤较小,而在高浓度下细胞膜受到海水毒害作用明显增加。
心律失常患者选择射频导管消融术治疗能够有效的改善患者心功能,帮助患者缓解临床症状,促进患者健康的恢复。但在使用射频导管消融术治疗中,手术中可能对患者和术者产生的辐射危害是一种潜在的风险,有可能对患者和医生的身体造成不良影响,为了减轻治疗过程中的辐射危害,在手术治疗过程中合理采取防护措施帮助降低辐射十分关键[1]。为了具体分析心律失常射频导管消融中的放射防护措施及效果,本文选取2016年12月—2017年12月在我院接受射频导管的心律失常患者72例进行分组研究如下。
2.3 海水胁迫对白子菜渗透调节物质的影响
2.3.1 海水胁迫对白子菜脯氨酸含量的影响 植物细胞的脯氨酸含量增加,维持了细胞的正常膨压,同时还可以保护酶的活性和使细胞膜系统免受伤害。白子菜在未使用海水处理下,其根、茎、叶中的脯氨酸含量分别为2.32、5.28、2.04 μg/g。由图3可知,海水胁迫下白子菜根、茎、叶中脯氨酸含量的变化趋势整体基本相同,都是随着胁迫浓度的增加而增加,在低浓度胁迫下脯氨酸含量变化不显著。在≥30%海水处理下,根中的脯氨酸含量显著增加,海水浓度最高时其含量最高,是对照的15.37倍;在≥20%海水处理下,茎、叶片中的脯氨酸含量显著增加,浓度最高时含量最高,分别是对照的6.21、1.95倍。说明白子菜在低浓度的海水胁迫下对脯氨酸含量变化无显著性影响,而在高浓度海水胁迫下较为敏感。

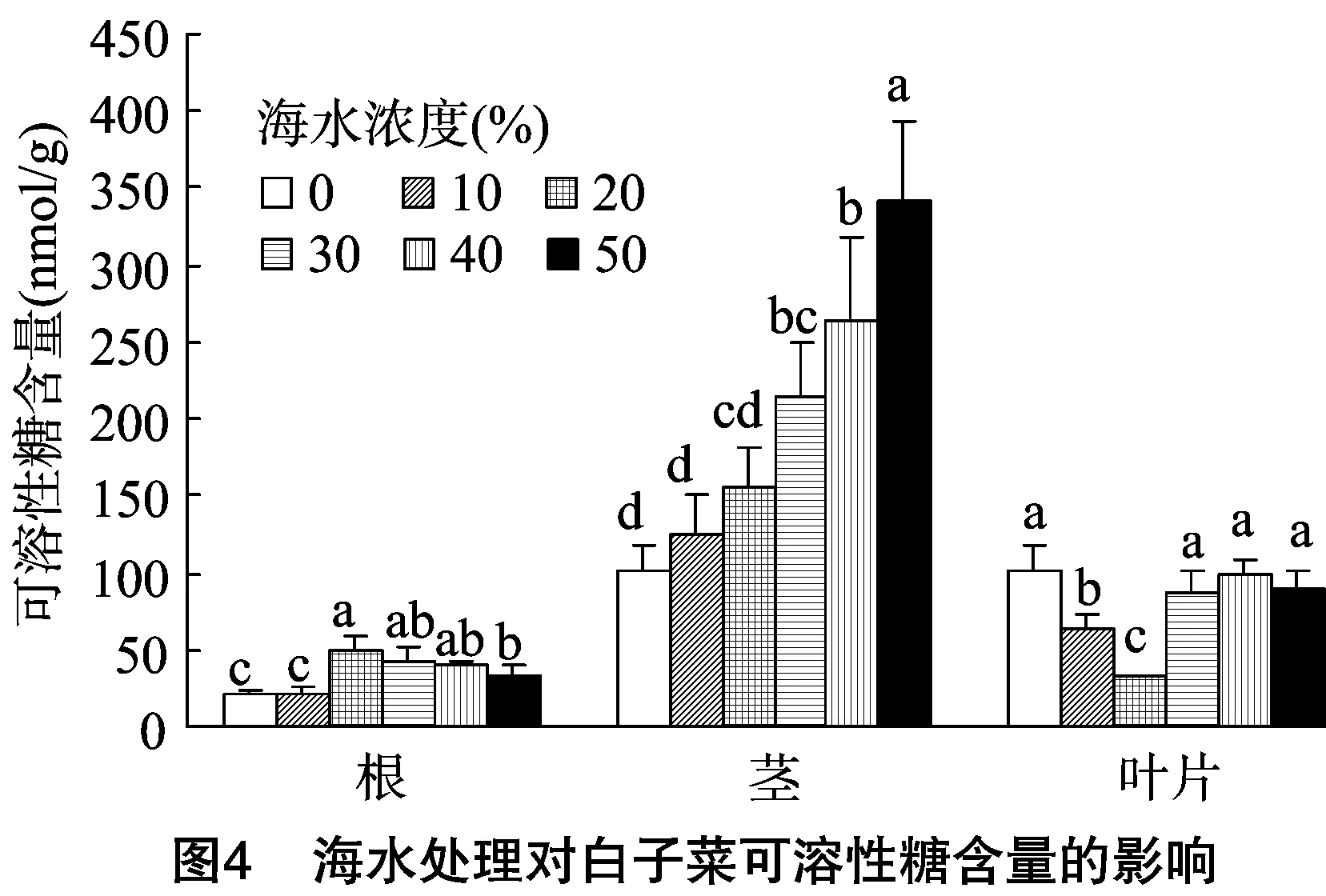
2.3.2 海水胁迫对可溶性糖含量的影响 可溶性糖作为植物重要的渗透调节物质,在逆境环境中其含量的增加有利于增强植物对环境的适应能力。由图4可知,白子菜在未经海水胁迫处理下,根、茎、叶中可溶性糖含量分别为20.98、101.79、100.66 nmol/g。在叶片中,可溶性糖含量随着海水浓度的升高呈先下降后上升的趋势,在20%海水浓度处理下达到最低值,较对照减少了14.5%;在30%~50%海水处理下,其含量略有提高。在根中,可溶性糖含量随着海水浓度的升高呈先增加后略有下降的趋势,在20%海水浓度处理下含量最高,为对照的2.38倍;在30%~50%海水浓度处理下略有下降,分别为对照的2.00、1.95、1.61倍。在茎中可溶性糖含量呈稳定增长趋势,在50%海水处理下达到最大值,为342.60 nmol/g,是对照的 3.37 倍,表明海水胁迫促进了可溶性糖在白子菜根茎中的积累。
2.4 海水胁迫对白子菜不同组织抗氧化酶系统的影响



2.4.2 海水胁迫对白子菜POD活性的影响 由图6可知,在未经海水胁迫处理下,白子菜根、茎、叶中POD活性分别为208.14、476.85、220.93 U/g。白子菜POD活性对海水胁迫较敏感,根、茎中分别在10%、20%海水胁迫下,POD活性较对照组显著增加,分别达到526.8、729.4 U/g,分别较对照增加了153.3%、50.8%,且随海水浓度的增加POD 活性增加,根中POD活性在50%海水浓度处理下达到最大值。叶中POD活性呈先增加后减小的变化趋势,除20%海水胁迫下显著增加,其余各个浓度与对照组相比没有显著性变化。POD活性的增加,有助于清除体内的H2O2,从而增强白子菜的耐盐性。

2.4.3 海水胁迫对白子菜CAT活性的影响 由图7可知,在未经海水处理下,白子菜根、茎、叶片中CAT活性分别为86.93、28.60、72.21 U/g。白子菜根中CAT活性随处理浓度的升高而升高,在10%~20%的海水处理下,其活性无明显增加,在30%~50%浓度下,活性明显增强,分别比对照增加了26.4%、26.2%、41.6%。茎、叶片中的变化趋势相似,在10%~30%浓度处理下,CAT活性显著增加,且30%处理浓度时达到最大值,分别是对照的1.66、1.93倍。在高浓度的海水胁迫下,茎、叶片中的CAT活性下降,但仍然略高于对照,说明白子菜中CAT活性能对海水胁迫作出响应。
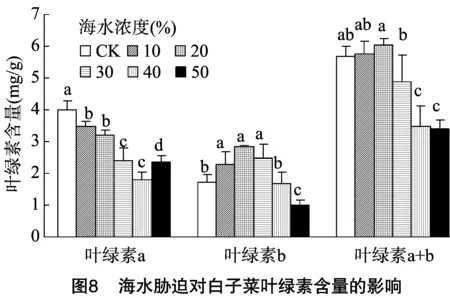
2.5 海水胁迫对白子菜叶绿素含量的影响

2.6 海水胁迫对白子菜根系活力的影响
根系活力是衡量植物根系吸收、合成、抵抗逆境能力大小的重要生理指标[23]。由图9可知,在未经海水处理下,白子菜根系活力为350.251 μg/(g·h)。在海水胁迫≤30%浓度处理下,根系活力保持在较高水平,且在10%处理浓度下,根系活力较对照显著增加了18.76%,在40%、50%浓度处理下,根系活力与对照相比下降,分别比对照减少了21.41%、12.03%。所以,低浓度的海水胁迫提高了白子菜的根系活力,且能忍受较高浓度的海水胁迫。

3 结论与讨论
植物在逆境胁迫下细胞内自由基代谢平衡遭到破坏,导致膜脂过氧化,使细胞膜系统严重受损。MDA是膜脂过氧化产物,能与细胞内各种成分发生反应,进而引起膜的损伤,其含量的高低可间接表现植物受逆境胁迫的强弱[24-25]。前人研究发现,在中低盐度胁迫下细胞膜透性增加不明显,而在高盐度胁迫下迅速增加,本研究也获得了相似结果,白子菜在≤30%海水胁迫下,MDA含量较对照略有升高,增加不明显,表明在此浓度范围内,白子菜对海水胁迫具有一定的适应性,而在≥40%海水胁迫下,MDA含量在根、茎中显著增加,其电导率的变化与MDA趋势基本相同,说明高盐胁迫下白子菜的细胞膜结构受到损伤,细胞内外的稳定性遭到破坏,造成植物细胞内的电解质渗漏增强,并随胁迫浓度的增加而增加。
脯氨酸和可溶性糖是植物体内重要的渗透调节物,有研究者将其作为植物抗盐的重要生理指标[26-27]。谭大海等发现,芦苇受盐胁迫时,能够大量迅速地累积脯氨酸来降低细胞渗透势,使植物体对盐分具有一定的耐受性[28]。本研究中白子菜体内的脯氨酸含量随盐浓度的升高大量积累,并且浓度越高积累越迅速,根茎中的积累量大于叶片中的,表明游离脯氨酸的积累是白子菜适应盐胁迫的重要途径之一,根的适应性高于叶片,这是植物为了对抗盐胁迫而采取的一种保护性渗透调节反应。可溶性糖为有机物质的合成提供碳架和能量来源,能稳定细胞膜,是重要的渗透调节物质[29]。在本研究中,白子菜根茎中的可溶性糖含量随海水处理浓度的升高整体呈上升的趋势,说明植物可通过可溶性糖的积累来增强渗透调节能力,而在叶片中表现为先减小后增加的趋势,可能是由于低浓度的海水胁迫使白子菜的生长维持在较高的水平,并且海水中一些矿物元素促进了植物的吸收,而在高浓度(≥30%)胁迫下,其含量显著增加。高盐胁迫下,可溶性糖含量增加也有可能是由于高浓度盐胁迫下植物生长受抑制,导致糖利用减少,可溶性糖含量增加[30]。白子菜中脯氨酸与可溶性糖含量的增加,有助于维持细胞正常的膨压,有利于细胞生理生化过程的正常进行;保持细胞持续生长,以减轻逆境对细胞生长的抑制;对于保持原生质体与环境的平衡和膜结构的完整性具有重要的作用,此外,还可以维持生物膜的稳定性和某些酶的活性。
植物在逆境胁迫下,会产生大量对植物有强烈毒害作用的活性氧,破坏植物体的动态平衡,使膜透性增加,代谢紊乱,此时植物会启动细胞内的保护酶(SOD、CAT、POD等)系统,从而及时清除体内由于氧化胁迫产生的大量自由基来维持正常的代谢[31-32]。SOD、POD和CAT是酶保护系统中的重要组成部分,可以清除过氧化物、过氧化氢和氧自由基,避免或减轻植物受到伤害,使植物正常生长代谢[33]。本研究中,在低浓度海水(≤30%)处理下,3种酶活性都有升高,表明植物对海水胁迫作出了反应。高浓度海水(≥40%)处理的白子菜根中SOD、POD、CAT活性维持在较高水平,而茎、叶中SOD活性较对照无明显变化,说明白子菜各组分因海水的胁迫产生了大量的活性氧,抑制了活性氧清除酶活性,SOD、POD、CAT协同作用下降,活性氧的产生与清除失去平衡,保护酶活性的变化与MDA含量变化相一致。
叶绿素参与植物的光合作用,是光合作用的重要物质,逆境胁迫会抑制植物叶绿素合成的相关酶类,导致叶绿素生成受阻、含量下降,叶绿素功能结构受损、氧化分解增强。本研究中,叶绿素a与叶绿素b的变化不一致,说明二者对盐害的适应性不同,在海水浓度≤30%时,叶绿素b含量增加,叶绿素a含量减少,而叶绿素总量无显著减少,表现出较强的忍耐能力。这可能是由于低钠盐可以增加植物体内的叶绿素含量,而高浓度海水胁迫破坏了植物叶绿体结构,使体内叶绿素含量下降,引起植株光合能力减弱[34-37]。
植物的根系活力能够客观地反映根系生命活动力,可以衡量植物根系的吸收和合成能力也是判断植物根系抵御逆境胁迫能力的重要生理指标,能够反映植物抗盐能力[38-40]。本研究中,在≤30%海水浓度处理下,白子菜根系能够保持较高的活力,但随着海水胁迫浓度(≥40%)的增高,根系受到损伤,根系活力较对照下降,但下降不明显。说明白子菜具有较强的耐盐性,这与张润花等对黄瓜[41]、陈炳东等对油葵的研究结果[42]相似。
综上,白子菜在海水浓度为0~30%时各项生理活动能正常进行,白子菜的生长未受到明显抑制,能正常生长;海水中丰富的矿质营养使得在≤20%海水浓度胁迫下生物量等指标增加。通过对各项生理指标的综合分析发现,白子菜在海水浓度≤30%时表现正常,其原因为(1)白子菜抗氧化酶(SOD、POD、CAT)的高活性,能最大限度地清除因海水胁迫产生的大量活性氧,减轻活性氧对白子菜细胞的损伤。(2)脯氨酸和可溶性糖积累,维持了白子菜的细胞渗透压。(3)根系活力处于较高水平有助于营养和水分的吸收,保障了地上部的营养供应。(4)叶绿素含量无明显减少,因而植物能进行正常的光合作用,维持正常的生理活动。当海水浓度≥40%时,白子菜叶绿素含量、根系活力降低抑制了白子菜的光合作用与营养物质的运输,膜系统受到的毒害作用增加,MDA大量积累,保护酶协同清除活性氧作用下降,白子菜生长受到显著抑制。
[1]Aroca R,Ruiz-Lozano J M,ZamarreoM,et al. Arbuscular mycorrhizal symbiosis influences strigolactone production under salinity and alleviates salt stress in lettuce plants[J]. Journal of Plant Physiology,2013,170(1):47-55.
[2]Li J G,Pu L J,Han M F,et al. Soil salinization research in China:advances and prospects[J]. Journal of Geographical Sciences,2014,24(5):943-960.
[3]Estrada B,Aroca R,Barea J M,et al. Native arbuscularmycorrhizal fungi isolated from a saline habitat improved maize antioxidant systems and plant tolerance to salinity[J]. Plant Science,2013,201(1):42-51.
[4]Lieth H. Concepts for different uses of halophytes[M]// Mangroves and halophytes:restoration and utilisation. Berlin:Springer Netherlands,2008:3-5.
[5]杨成龙,段瑞军,李瑞梅,等. 盐生植物海马齿耐盐的生理特性[J]. 生态学报,2010,30(17):4617-4627.
[6]王 蕾,吴朝波,徐微风,等. 海水胁迫对番杏生长、渗透调节物质和细胞膜透性的影响[J]. 江苏农业科学,2016,44(7):248-251.
[7]昊 霞,张钱钱,王忠震,等. 白子菜叶片中酸性多糖的降血糖作用及其对相关指标的影响[J]. 植物资源与环境学报,2015,24(2):115-117.
[8]杨 暹,刘厚诚. 白子菜的特征特性及栽培技术[J]. 广东农业科学,1998(5):16-17.
[9]关佩聪,吴琰芬. 8 种野菜的特征、生境、食法及保健作用[J]. 长江蔬菜,1999(11):28-30.
[10]李小军,覃 欢,刘 欢. 菊三七属植物的民族药用和食用价值[J]. 中南民族大学学报(自然科学版),2015,34(4):62-67.
[11]戴志聪,杜道林,司春灿,等. 用扫描仪及 Image J 软件精确测量叶片形态数量特征的方法[J]. 广西植物,2009,29(3):342-347.
[12]李合生,孙 群,赵世杰,等. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000.
[13]郝建军,康宗利,于 洋. 植物生理学实验技术[M]. 北京:化学工业出版社,2007.
[14]朱广廉,钟海文,张爱琴. 植物生理学实验[M]. 北京:北京大学出版社,1990:37-40.
[15]彭志英,蒋 黎. 紫外速率直接法测定过氧化氢酶活性[J]. 华西医学,1995,10(1):4-7.
[16]郑炳松. 现代植物生理生化研究技术[M]. 北京:气象出版社,2006:3-120.
[17]刘 宁,高玉葆. 渗透胁迫下多花黑麦草叶内过氧化物酶活性和脯氨酸含量以及质膜相对透性的变化J]. 植物生理学通讯,2000,36(1):11-14.
[18]陈建勋,王晓峰. 植物生理学实验指导[M]. 2版. 广州:华南理工大学出版社,2006:64-66.
[19]Heath R L,Parke L. Photoperitation in isolated diloroplastskinstics and stoichimertry fatty acid peroxidation [J]. Archbiophys,1968,75:189-198.
[20]张宪政. 作物生理研究法[M]. 北京:农业出版社,1992.
[21]陈少裕. 膜脂过氧化对植物细胞的伤害[J]. 植物生理学通讯,1991,27(2):84-90.
[22]张玉霞,李志刚,李美娟,等. 四种草地早熟禾抗盐碱生理生化特性的研究[J]. 中国农学通报,2004,20(5):209-213.
[23]Yanagawa H A,Watanabe K,Nakamura M. Application of the feed ingredients for livestock to an artificial diet by using polyphagous strain of the silkworm[J]. The Journal of Sericultural Science of Japan,1989,58(5):401-406.
[24]华 春,周泉澄,王小平,等. 外源GA3对盐胁迫下北美海蓬子种子萌发及幼苗生长的影响[J]. 南京师大学报(自然科学版),2007,30(1):82-87.
[25]孙海菁,王树凤,陈益泰. 盐胁迫对6个树种的生长及生理指标的影响[J]. 林业科学研究,2009,22(3):315-324.
[26]刘占彬,袁庆华,景启美. 9个高羊茅品种苗期耐盐性研究[J]. 中国草地学报,2007,29(6):81-85.
[27]马宗仁,郭 博. 短芒披碱草和老芒麦在水分胁迫下游离脯氨酸积累的研究——Ⅰ. 植物的抗旱性与脯氨酸积累能力关系的标准[J]. 中国草地,1991,4:12-16.
[28]谭大海,沙 伟,张莹莹. 芦苇盐胁迫下渗透调节物质含量变化研究[J]. 齐齐哈尔大学学报(自然科学版),2006,22(2):84-86.
[29]李圆圆,郭建荣,杨明峰,等. KCl和NaCl处理对盐生植物碱蓬幼苗生长和水分代谢的影响[J]. 植物生理与分子生物学学报,2003,29(6):576-580.
[30]Nixon P J,Mullineaux C W.Regulation of photosynthetic electron transport[M]//Regulation of Photosynthesis. Springer Verlag:Springer Netherlands,2001:533-555.
[31]Ouariti O,Boussama N,Zarrouk M,et al. Cadmium-and copper-induced changes in tomato membrane lipids[J]. Phytochemistry,1997,45(7):1343-50.
[32]Kanazawa S,Sano S,Koshiba T,et al. Changes in antioxidative enzymes in cucumber cotyledons during natural senescence:comparison with those during dark-induced senescence[J]. Physiologia Plantarum,2001,109(2):211-216.
[33]刘爱荣,张远兵,陈登科. 盐胁迫对盐芥(Thellungiellahalophila)生长和抗氧化酶活性的影响[J]. 植物研究,2006,26(2):216-221.
[34]Munns R,Termaat A. Whole-plant responses to salinity[J]. Functional Plant Biology,1986,13(1):143-160.
[35]Matoh T,Murata S. Sodium stimulates growth of Panicum coloratum through enhanced photosynthesis[J]. Plant Physiology,1990,92(4):1169-1173.
[36]汪贵斌,曹福亮. 盐分和水分胁迫对落羽杉幼苗的生长量及营养元素含量的影响[J]. 林业科学,2004,40(6):56-62.
[37]吴永波,薛建辉. 盐胁迫对3种白蜡树幼苗生长与光合作用的影响[J]. 南京林业大学学报(自然科学版),2002,26(3):19-22.
[38]王素平,郭世荣,胡晓辉,等. 盐胁迫对黄瓜幼苗叶片光合色素含量的影响[J]. 江西农业大学学报,2006,28(1):32-38.
[39]宋海星,王学立. 玉米根系活力及吸收面积的空间分布变化[J]. 西北农业学报,2005,14(1):137-141.
[40]高福元,张吉立,刘振平,等. 盐胁迫对树锦鸡儿叶绿素含量和根系活力的影响[J]. 贵州农业科学,2010,38(5):46-48.
[41]张润花,郭世荣,李 娟. 盐胁迫对黄瓜根系活力、叶绿素含量的影响[J]. 长江蔬菜,2006(2):47-49.
[42]陈炳东,黄高宝,陈玉梁,等. 盐胁迫对油葵根系活力和幼苗生长的影响[J]. 中国油料作物学报,2008,30(3):327-330.
