印楝无性系当代种子农药品质性状的遗传变异及农药型印楝优株评价
2018-01-03彭兴民吴疆翀郑益兴赵保荣赵培仙张燕平
彭兴民 , 吴疆翀, 郑益兴, 罗 彰, 赵保荣, 赵培仙, 张燕平
(1.中国林业科学研究院资源昆虫研究所,云南 昆明 650224; 2.云南省元阳县林业局,云南 元阳 662400; 3.云南省元江县林业局,云南 元江 653300; 4.云南省元谋县林业局,云南 元谋 651300)
印楝无性系当代种子农药品质性状的遗传变异及农药型印楝优株评价
彭兴民1, 吴疆翀1, 郑益兴1, 罗 彰2, 赵保荣3, 赵培仙4, 张燕平1
(1.中国林业科学研究院资源昆虫研究所,云南 昆明 650224; 2.云南省元阳县林业局,云南 元阳 662400; 3.云南省元江县林业局,云南 元江 653300; 4.云南省元谋县林业局,云南 元谋 651300)
目的建立一套适合印楝药用优树无性系测定的分析评价方法,并进行农药型印楝优株评价。方法以印楝药用优树和平均木无性系为试验材料,通过不完全谱系设计建立无性系品比试验,采用重复力估算法在表型水平上对无性系当代种子农药品质性状进行遗传变异分析,提出“农药型印楝”的概念和标准,根据该标准进行农药型印楝优株评价。结果无性系种子印楝素组分含量随优树种子印楝素组分含量的变化而变化,但在无性系内和无性系间都存在不同程度变异:同一无性系各分株间的印楝素B变异(5.26%37.55%)较印楝素A变异 (6.06%15.80%)和印楝素AB变异(2.95%16.14%)大,印楝素B和印楝素AB几乎呈退化和/或趋中遗传倾向,而印楝素A则既有退化和/或趋中遗传倾向又有超母遗传倾向;无性系间种子印楝素组分含量性状具有异质性(P<0.01)。无性系印楝素A重复力(0.78)、印楝素B重复力(0.81)和印楝素AB重复力(0.83)均较高,表明无性系间印楝素组分含量的差异主要是由遗传效应和一般环境效应造成的,受特殊环境效应影响较小。按农药型印楝标准,评定LD0505、WW0401、AT0515三个无性系为“农药型印楝品种”, LD0505和WW0421是“农药型印楝亲本”。结论重复力估算法适用于印楝药用优树无性系测定的分析评价,优树种子农药品质性状的遗传传递力强,无性系能保持优树农药品质的优良特性;评定LD0505、WW0401、AT0515为农药型印楝品种,LD0505和WW0421还可作为农药型印楝亲本,其余优树无性系不宜选择。
农药型印楝;无性系;种子农药品质;方差分析;重复力
印楝(AzadirachtaindicaA.Juss.)为楝科(Meliaceae)常绿乔木[1],因含有天然产物农药印楝素(azadirachtin)而被称为可以直接加工农药的植物即农药植物(pesticidal plant)[2-4]。印楝素有十余种异构体及其衍生物,其中印楝素A(azadirachtinA)和印楝素B(azadirachtinB)是印楝素的主要成分[5]。印楝素主要来源于印楝种子[6]。印楝种子品质依据研究或利用角度的不同,分为农药品质、外观品质、贮运品质、加工品质等,构成因素包括种子的农药活性物质、大小、形状、黄曲霉素、质地等。由印楝素A和印楝素B等因素组成的农药活性物质,不仅影响农药品质,而且是决定外观品质、贮运品质、加工品质的关键因子。因此,种子高印楝素A和印楝素B含量是印楝农药品质育种的目标[7-8]。进一步加强印楝引种资源评价及亲本利用研究,努力提升印楝农药原料林种子农药品质,协同拓宽印楝栽培品种的遗传基础,对于推动我国印楝农药原料生产的优质高效发展具有重要意义。
本课题通过印楝资源调查、表型选择,从233株药用候选优树中筛选出药用初选优树14株[9-10],并于2005年,在我国印楝引种栽培的中心分布区——云南省元阳县的红河流域干热河谷地区开展印楝药用优树无性系比较试验(或无性系测定)。对上述药用优良单株种子农药品质性状的遗传变异分析及农药型印楝优株评价,需要建立一套适宜的分析评价方法。果树等无性繁殖植物的育种,无性系能保持优树性状的比率和同一无性系各分株性状的稳定程度,不能用遗传力来度量,于是育种家们引入了重复力的概念[11-12]。重复力是一个最容易估计、十分重要且潜力很大实用性的遗传参数,重复力估算法在林木无性系育种上已有成功的尝试[13]。本研究在建立印楝药用优树无性系比较试验园的基础上,测定种子印楝素A、印楝素B及印楝素A和印楝素B总含量(印楝素AB)等3 个评价指标,采用重复力估算法研究印楝优树无性系当代种子农药品质性状的遗传变异特点,并进行农药型印楝优株评价,旨在为农药型印楝育种技术体系的创建及农药型印楝资源的利用保存提供基本资料。
1 材料与方法

1.1 试验材料
1.2 测定方法与指标
采用宗乾收等[14]提出的“印楝种仁中印楝素含量的快速液相色谱分析”法,分析种子印楝素A和印楝素B含量。计算印楝素A B总含量:印楝素A B= 印楝素A +印楝素B。
1.3 数据统计分析
重复力在生物学上的定义就是基因型方差与一般环境方差之和在表型方差中所占比例,而统计学上的定义则是同一生物个体或同一无性系在多次生产记录之间的组内相关系数[12]。单株小区无性系比较试验,无性系重复力的推算过程:首先将观测数据进行单因素随机区组设计方差分析,计算无性系间均方、机误(或环境)均方、无性系间方差或期望均方、机误(或环境)方差或期望均方;再根据重复力计算公式计算无性系重复力。
方差分析线性模型为:xij=m+ai+bj+eij。根据这一模型,每一观察值xij含有总体平均值m、处理效应ai(=mi.-m)、区组效应bj(=mj-m)以及随机误差εij。方差分析列表如表1。

表1 单株小区无性系方差分析
注:n, 无性系数量;k, 同一无性系的个体数。
重复力的计算式[12]为:
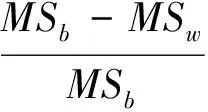

数据的描述统计由Microsoft Office Excel 2003软件完成;方差分析采用SAS 9.0 system for windows (SAS Institute Inc., Cary, NC, USA.)软件,由SAS的anova过程实现无协变量平衡随机区组设计试验方差分析,指定邓肯新复极差法(Duncan’s Multiple Range Test)作均值多重比较[15]。
2 结果与分析
2.1 优树无性系当代种子农药品质性状的遗传变异分析
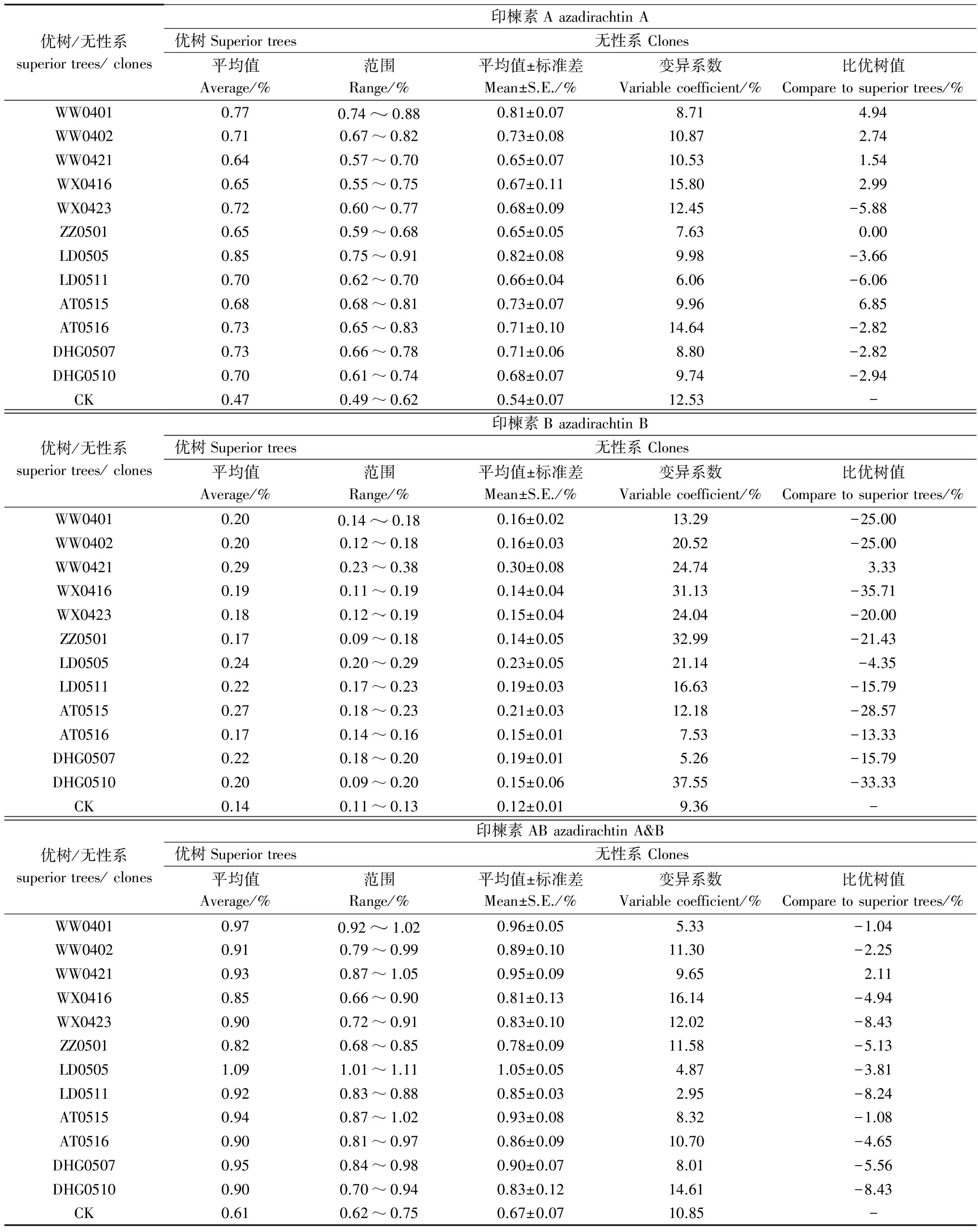
表2 印楝优树无性系种子印楝素组分含量的遗传倾向
无性系间种子印楝素组分含量的差异显著性分析结果(表3)表明,12个优树无性系种子印楝素组分含量均优于对照,表明无性系种子印楝素A、印楝素B和印楝素AB的优良特性是真实存在的(P<0.01),这为优良无性系的选择提供了可能。
2.2 优树无性系当代种子农药品质性状的稳定性分析
无性系种子农药品质性状的方差分析和重复力如表3。由表3可知,供试无性系种子印楝素A、印楝素B和印楝素AB的决定系数分别为0.754 2、0.727 1、0.779 5,可解释的模型效应均达70%以上,说明试验结果可靠。
重复力的大小表明性状受特殊环境影响的大小和其稳定性的高低,而且在一定的试验条件下,只要充分实现试验材料和试验地的一致性,尽量避免一般环境方差的影响,就可把重复力作为广义遗传力上限的估计值[13]。反映在重复力或广义遗传力上限的估计值上,印楝素A、印楝素B和印楝素AB的重复力分别为0.78、0.81和0.83或广义遗传力上限的估计值分别为78%、81%和83%,都属于高重复力性状,表明无性系能保持优树种子农药品质性状的比率较大,优树种子农药品质性状的遗传传递力强,种子印楝素组分含量性状由遗传效应和一般环境效应决定,受特殊环境效应影响较小,同一无性系各分株种子农药品质性状的稳定程度较高,无性系很好地保持优树种子农药品质的优良特性。

表3 印楝优树无性系品比试验因子效应方差分析及重复力
各个无性系两两之间的差异显著性如表4。从表4不难看出,在α=0.01水平上,13个无性系种子的印楝素A含量大小排序为:LD0505>WW0401>AT0515、WW0402、HGD0507、AT0516>WX0423、DHG0510、WX0416>LD0511>WW0421、ZZ0501>CK;印楝素B含量大小排序为:WW0421>LD0505>WW0401、AT0515、WW0402、LD0511、HGD0507、AT0516、WX0423、DHG0510、WX0416、ZZ0501>CK;印楝素AB含量大小排序为:LD0505>WW0401、AT0515、WW0421、WW0402、HGD0507>AT0516、LD0511>WX0423、DHG0510、WX0416、ZZ0501>CK。

表4 印楝无性系均值的多重比较
从表4还可看出,12个优树无性系种子印楝素A、印楝素B和印楝素AB含量排名前三名的无性系,即LD0505、WW0401、AT0515,在α=0.05水平上,都与其它无性系差异显著。
2.3 农药型印楝优株的评价
各个无性系嫁接成效和发枝结果情况如表5。无性系间嫁接成活情况以及萌发枝条数、结果枝条数、结果枝开花情况等差异不大,但结实情况差异较明显。嫁接成活率均约大于80%,亲和性较好;13个无性系枝条平均结果数量由高到低排序:AT0515>DHG0510>LD0511>AT0516>DHG0507>LD0505>WW0421>WW0401>WW0402>WX0423>WX0416>ZZ0501>CK,优树无性系的结果数量均较对照大,产量性状优异的比对照高出101.16%,较好的也比对照高50%,产量性状优良,保持了优树高产的特性。
LD0505和WW0421是12个优树无性系中种子印楝素A、印楝素AB和印楝素B含量排名第一的无性系。LD0505的印楝素A和印楝素AB表现优异(印楝素A=0.82%和印楝素AB=1.05%),而WW0421的印楝素B表现优异(印楝素B=0.30%),将其称为“农药型印楝亲本”,可作为印楝农药品质杂交育种的杂交亲本。
3 讨论
3.1 关于印楝无性系当代种子农药品质性状的遗传变异问题
自然授粉状态下印楝异交率高于90%[16]。因此,无性系多系杂交后各无性系间种子印楝素组分含量具有异质性(P<0.01)。重复力反映了无性系优良性状受特殊环境影响的程度[12-13]。本研究种子印楝素组分含量约0.80的重复力值(即80%的广义遗传力上限估值),表明印楝素组分含量在无性系种子间的总体变异情况为,约4/5是由无性系间的遗传差异造成的,其余1/5是由无性系间的特殊环境效应造成的,遗传效应的影响大于特殊环境效应的影响。
本研究还发现,多系杂交,无性系当代种子印楝素组分含量性状存在超母遗传倾向、退化和/或趋中遗传倾向。综合印楝素AB遗传表现,无性系当代种子印楝素组分含量平均值较优树平均值低,呈退化遗传倾向。那么,印楝无性系当代种子印楝素组分含量性状普遍退化的遗传原因是什么呢?
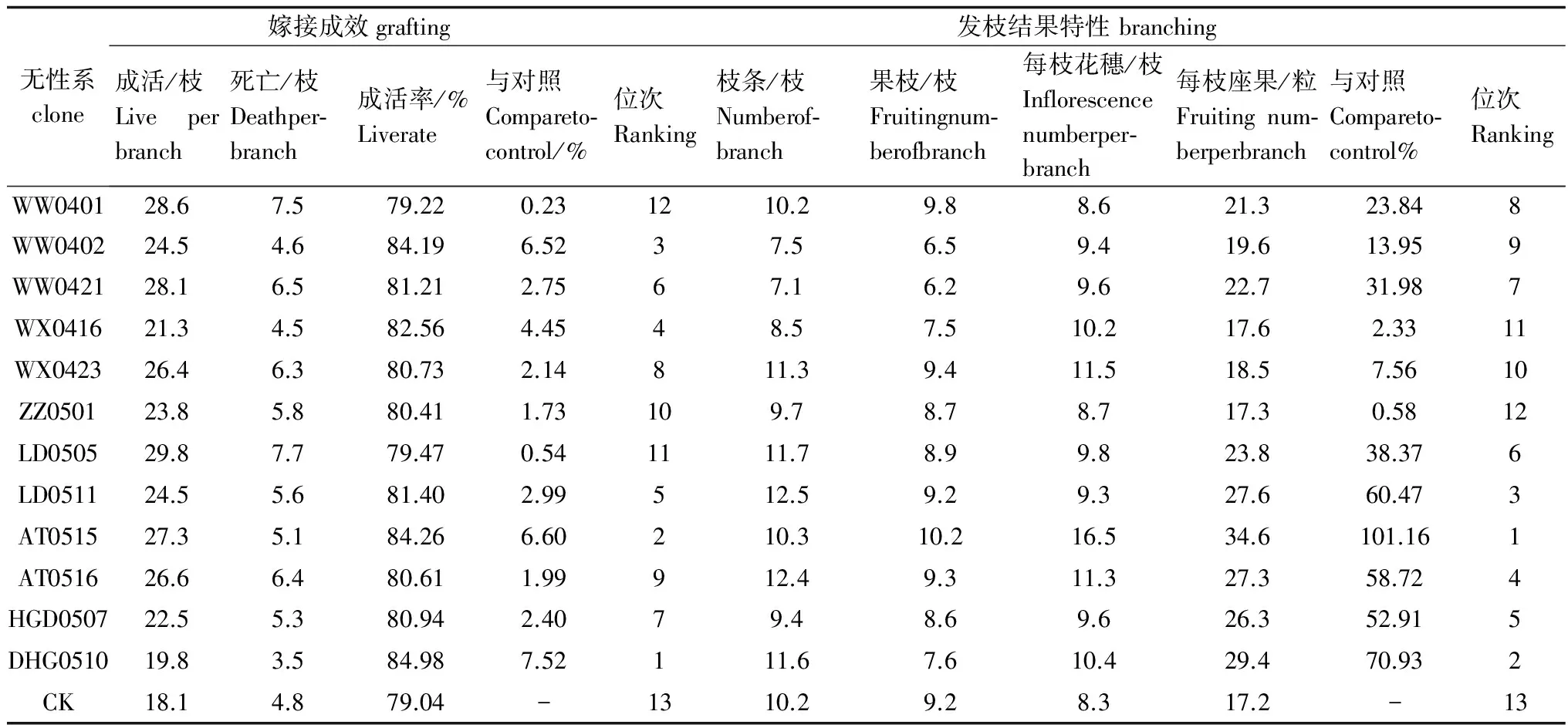
表5 印楝无性系嫁接成效及发枝结果特性
注:嫁接成效的“与对照±%”和“位次”是成活率(%);发枝结果特性的“与对照±%”和“位次”是每枝座果(粒)。
3.2 关于印楝无性系当代种子农药品质性状的遗传稳定性问题
重复力的大小还表明性状稳定性的高低[13]。本研究一方面种子印楝素组分含量性状0.80左右的重复力或80%左右的广义遗传力上限的估计值,表明无性系能保持优树种子农药品质性状的比率较大,同一无性系各分株种子农药品质性状的稳定程度较高。
另一方面种子印楝素组分含量性状普遍退化。果树等无性繁殖植物育种的遗传研究结果证实:无性系杂交或进行有性繁殖,无性系遗传值中的非加性效应部分通过基因重组而解体,不能固定遗传。这是果树等无性繁殖植物在有性繁殖时一系列经济性状衰退在遗传上的真正原因[17]。据此不难对本研究结果作以下解释或推论:印楝优树无性系杂交或进行有性繁殖,无性系遗传值中的非加性效应部分通过基因重组而解体,不能固定遗传,导致杂交当代种子印楝素组分含量性状普遍退化。这就是本研究优树无性系在有性繁殖时当代种子农药品质性状衰退遗传的真正原因。印楝素B衰退程度较印楝素A大,表明印楝素B的非加性效应较印楝素A大,是非加性效应比重较大的基因型。
3.3 关于农药型印楝及其育种技术问题
印楝良种化是生物农药品质改良的重要方式之一。世界范围内印楝种子印楝素A含量平均为0.36%[18],其中国内栽培印楝种子印楝素A平均含量为0.55%,但变异幅度较大,达到农药原料林标准的植株仅有约7%[7,10]。本文认为“农药型印楝”应满足下列 3 个条件:①种子印楝素A含量2倍于世界平均含量,即印楝素A≥0.72%;②经区域和品比试验结实量高于平均值,具有丰产特性;③具有种源特有的遗传多样性。
资源亲本利用也是印楝种子农药品质改良的重要方式之一。杂交育种中,亲本选择与选配是决定品质改良成效大小的关键,优良性状功能基因挖掘是亲本利用的基础。LD0505和WW0421是12个优树无性系中种子印楝素A、印楝素AB和印楝素B含量排名第一的无性系。LD0505的印楝素A和印楝素AB表现优异,而WW0421的印楝素B表现优异,将其称为“农药型印楝亲本”。
4 结论
重复力估算法适用于印楝药用优树无性系测定的分析评价,优树种子农药品质性状的遗传传递力强,无性系能保持优树农药品质的优良特性;评定LD0505、WW0401、AT0515为农药型印楝品种,LD0505和WW0421还可作为农药型印楝亲本,其余优树无性系不宜选择。
[1] 彭兴民,吴疆翀,郑益兴,等. 印楝属(AzadirachtaA. Juss. ) 植物分类及分布的研究现状[J]. 植物遗传资源学报,2012,13( 4):583-588.
[2] Tomilin C D. The Pesticide Manual.12nd ed[M]. UK: Surrey, British Crop Protection Council, 2000.
[3] International Neem Network. Establishment of the Network[EB/OL]. (2007-01-16)[2015-07-21] http://www.fao.org/forestry/neem/5309/zh/
[4] Krishnan N M, Pattnaik1 S, Jain P.etal. A draft of the genome and four transcriptomes of a medicinal and pesticidal angiosperm Azadirachtaindica[J]. BMC Genomics, 2012,13(1): 464.
[5] 樊会丹,张从海,严胜骄,等. 印楝素的合成、结构修饰及生物活性研究进展[J]. 有机化学,2009,29(1): 20-33.
[6] 谭卫红. 印楝种仁成分的化学及活性成分研究[D]. 北京: 中国林业科学研究院, 2007.
[7] 彭兴民,吴疆翀,王有琼,等.印楝种子品质性状的遗传多样性及稳定性分析[J]. 林业科学研究,2016,28(6):767-774.
[8] 张志祥,程东美,徐汉虹,等. 印楝素A 和印楝素B 的生物活性及增效作用[J]. 华中农业大学学报,2004,23(5):515-518.
[9] 彭兴民,吴疆翀,王有琼,等. 药用印楝表型选择的因子分析及综合评价[J]. 林业科学研究,2015,28(4):464-472.
[10] 彭兴民,吴疆翀,张燕平,等.印楝农药原料林优树选择方法与标准[J]. 福建林学院学报,2010,30(3):265-269.
[11] 吴 裕,毛常丽. 树木育种学中遗传力、重复力和遗传增益的概念及思考[J]. 热带农业科技,2012,35 (1):47-50.
[12] 续九如. 重复力及其在树木育种中的应用[J]. 北京林业大学学报,1988,10(4):97-102.
[13] 王 琦. 林木无性系育种若干遗传参数估算的研究进展[J]. 林业科学, 1995,31(2):169-176.
[14] 宗乾收,林 军,武永昆,等. 印楝种仁中印楝素含量的快速液相色谱分析[J]. 农药, 2003,42(4):23-24.
[15] 阮 敬. SAS统计分析从入门到精通[M]. 北京:人民邮电出版社出版. 2009.
[16] 吴疆翀. 印楝繁育系统的研究[D]. 北京:中国林业科学研究院,2007.
[17] 景士西,吴录平,李宝江. 果树遗传变异的特点初探[J]. 遗传, 1995,17(1):40-44.
[18] Ermel K. #Azadirachtin content of neem seed kernels from different regions of the world[C] //Schmutterer H. The Neem Tree: Source of Unique Natural Products for Integrated Pest Management, Medicine, Industry and Other Purposes.Weinheim,Germany: VCH Verlagsgesellschaft,1995:89-92.
GeneticVariationofAzadirachtinQualityinSeedsofNeemClonesandEvaluationofSuperiorNeemTreesforPesticides
PENGXing-min1,WUJiang-chong1,ZHENGYi-xing1,LUOZhang2,ZHAOBao-rong3,ZHAOPei-xian4,ZHANGYan-ping1
(1. Research Institute of Resource Insects, Chinese Academy of Forestry, Kunming 650224, Yunnan, China; 2.Forestry Bureau of Yuanyang County,Yuanyang 662400, Yunnan, China; 3.Forestry Bureau of Yuanjiang County, Yuanjiang 653300, Yunnan, China; 4.Forestry Bureau of Yuanmou County,Yuanmou 651300, Yunnan, China)
ObjectiveTo evaluate superior trees for pesticides and provide practical methods for assessment of pesticidal neem clones.MethodWith azadirachta content in seeds tested, the variety comparative analysis of azadirachta-related phenotypic traits between superior pesticidal neem trees and average control group was performed by incomplete pedigree design, and then the repeat force was calculated to establish the standards of the potential cultivar type “pesticidal neem”. Then the superior trees for pesticides were evaluated.ResultThe azadirachta content in seeds of clones varied with the azadirachta content in seeds of superior trees. The azadirachta content varied both within and among clones. The variation of azadirachta content in same clone ranged as: azadirachta B (5.26%37.55%) > azadirachta A (6.06%15.80%) > azadirachta A+B (2.95%16.14%). Both the contents of azadirachta B and azadirachta A+B showed degradation and neutrality of genetic predispositions, but the content of azadirachta A showed all genetic predispositions including degradation, neutrality and development. The azadirachta content in seeds of various clones was significantly different (P<0.01) .For stability of azadirachta content among superior trees and clones, the coefficient of azadirachta A, B, and A+B was 0.78, 0.81, and 0.83, respectively, indicating that the variance of azadirachta content among clones was mainly influenced by genetic effects and normal environmental effects than some particular environmental effects. According to comprehensive performance of superior tree clones, three clones (LD0505, WW0401, and AT0515) were selected as the “variety of pesticidal neem”. LD0505 and WW0421 were “parentage of pesticidal neem”.ConclusionThe repeatability of seed azadirachta content could be used to evaluate the quality of superior neem clones, to select variety and parentage of pesticidal neem trees. Analysis results indicates that superior pesticidal features could remain in same clone by genetic heritability. LD0505, WW0401 and AT0515 could be further developed as the varieties of pesticidal neem. Besides, LD0505 and WW0421 could be further developed as parentage of pesticidal neem. Other superior tree clones are unfavorable choice.
pesticidal neem; clones; seed azadirachtin quality; ANOVA; repeatability
10.13275/j.cnki.lykxyj.2017.06.006
2017-04-15
国家自然科学基金(31500515、31270710)和国家“十二·五”科技支撑计划项目(2012BAD21B04)
彭兴民(1962—),高级工程师,主要研究方向:植物引种、种质创新与新品种选育。E-mail:penggong007@21cn.com
S722.5
A
1001-1498(2017)06-0921-08
张 玲)
