厚壳贻贝(Mytilus coruscus)金属硫蛋白MT-10:cDNA克隆、结构分析及铜离子胁迫下的表达*
2017-12-14王昊盛宋鑫金董文强陈永霞卢玮筱祁鹏志郭宝英
王昊盛 宋鑫金 董文强 吴 斌 陈永霞 卢玮筱 祁鹏志 郭宝英
(浙江海洋大学 国家海洋设施养殖工程技术研究中心 舟山 316022)
厚壳贻贝(Mytilus coruscus)金属硫蛋白MT-10:cDNA克隆、结构分析及铜离子胁迫下的表达*
王昊盛 宋鑫金 董文强 吴 斌 陈永霞 卢玮筱 祁鹏志①郭宝英
(浙江海洋大学 国家海洋设施养殖工程技术研究中心 舟山 316022)
近年来我国沿海海域重金属污染日趋严重, 软体动物尤其是双壳贝类响应重金属污染胁迫的研究发展成为目前的研究热点。金属硫蛋白是一类广泛存在于生物体内富含半胱氨酸(Cys)的小分子蛋白, 具有结合金属离子的能力, 能有效调节生物体细胞内的金属离子平衡, 具有清除自由基,重金属解毒的功能, 关于厚壳贻贝MT蛋白的研究尚未见报道。本文首次利用RT-PCR和cDNA末端快速扩增技术克隆了厚壳贻贝 MT-10蛋白 cDNA全长(Mc-MT-10), 其包含 101bp的 5′ UTR区,108bp的3′UTR区以及222bp的ORF区, 编码73个氨基酸。经比对发现Mc-MT-10富含Cys, 且具有9个软体动物中常见的CXC结构以及CKCXXXCXCX结构, 系统进化树中同科或属中MT-10聚合成一个分支, 显示出高度保守性。采用Q-PCR技术分析了MT-10 mRNA在Cu2+胁迫下厚壳贻贝外套膜及消化腺中表达变化, 结果表明Mc-MT-10mRNA在Cu2+胁迫下两种组织中的表达量急剧升高, 说明Cu2+能够诱导Mc-MT-10mRNA的表达, 并且表现出诱导具有阈值性特点。研究结果可为深入研究厚壳贻贝MT蛋白的功能奠定基础, 为厚壳贻贝抗重金属污染新品种选育提供数据支持。
厚壳贻贝; 金属硫蛋白基因; 荧光定量PCR; 铜胁迫
厚壳贻贝(Mytilus coruscus), 属贻贝目(Mytiloida)、贻贝科(Mytilidae)、贻贝属(Mytilus), 是海洋双壳贝类中生长速度最快的种类之一。主要分布于我国黄渤海、东海、台湾等地, 在日本北海道、韩国济州岛等地也有分布。随着工业进程的不断深入,给沿海海域环境带来巨大压力, 工业废水、船舶油污、生活污水等造成的海水污染不断加剧, 许多重金属严重超标(郭远明等, 2012)。重金属不易降解, 在食物链中不断富集, 人类食用富含重金属的鱼虾贝等海产品会产生中毒现象, 对健康危害极大(Shuaiet al,2001)。双壳贝类对重金属具有较强的生物富集能力,且活动性低, 是天然的重金属污染指示物。其在重金属胁迫下的变化可间接反映在高浓度金属离子胁迫下生物机体内的氧化反应程度, 是指示环境污染的重要标志(Mooreet al, 2004)。贻贝作为重金属污染指示物在很多国家已经得到应用(Miller, 1999), 相关基础研究也逐渐得到重视。但现阶段国内外对贻贝的研究主要集中于翡翠贻贝与紫贻贝, 对厚壳贻贝的研究主要集中在人工育苗养殖(常抗美等, 2007), 遗传特异性(Yeet al, 2012)以及营养成分比对分析(Kimet al, 2012)等方面。本研究旨在填补贻贝属中厚壳贻贝的作为重金属污染指示物研究, 为今后对厚壳贻贝进行更加全面深入的科研提供基础。
金属硫蛋白(metallothioneins, MT)是一种富含半胱氨酸(Cys)及金属离子的小分子蛋白。通常含有CC、CXC等结构, 因为半胱氨酸(Cys)能与重金属离子结合因而取名金属硫蛋白, 在一定程度上起到重金属解毒和自由基清除等修复作用(刘维青等, 2006)。MT蛋白在生物体内的的表达水平通常也反映了该生物对重金属的应激及自我修复能力, 同时也能作为环境中重金属浓度的指示物。近年来国内外已经出现了许多关于生物体内MT的报道(Wanget al, 2011), 主要集中于各种鱼类(Olssonet al, 1998; Roevaet al,1999)和水生无脊椎动物(Roesijadiet al, 1991), 以及大量的软体动物(Isaniet al, 1997; Langstonet al, 1998)和甲壳动物(Roesijadiet al, 1997; Engelet al, 1993;Barka, 2000)。但厚壳贻贝 MT基因的研究尚未见报道。本研究通过 RACE技术克隆了厚壳贻贝 MT-10全长cDNA序列, 通过铜离子胁迫, 模拟当前海洋环境重金属污染现状, 通过对厚壳贻贝细胞内 MT-10 mRNA相对表达量的测定及分析, 论证厚壳贻贝遭遇重金属污染之后的应激行为及对重金属的自我修复能力, 为厚壳贻贝今后作为海洋环境中重金属浓度指示物提供理论基础。同时也为解决当前厚壳贻贝养殖业面对海洋重金属污染的困境提供一定建议。
1 材料与方法
1.1 实验材料
本次实验所用的厚壳贻贝(Mytilus coruscus)于2016年10月采集于舟山东极青浜岛附近海域, 外壳平均长度为(9.25±0.43)cm, 壳宽(4.51±0.3)cm, 壳高(3.05±0.20)cm, 湿重(60.18±6.45)g。清污后置于蓝色养殖桶内暂养, 每天换取经过过滤的海水, 10d之后选择生命特征明显的个体进行对照实验。
1.2 方法
1.2.1 铜离子胁迫 将选择后的厚壳贻贝随机分为两组, 每组 40只。实验组海水中加入硫酸铜作为铜离子源, 使 Cu2+终浓度为 20μg/L。每天换水一次,更新一半并在实验组中添加硫酸铜维持Cu2+浓度。
1.2.2 RNA提取及逆转录 处理后2d、5d、10d、15d、21d、28d进行取样。对照组和实验组随机取出4个的厚壳贻贝, 解剖得到外套膜及消化腺存放于–80°C冰箱内暂存。采集厚壳贻贝腮、性腺、消化腺、血细胞、肾脏、肌肉及外套膜组织, 存放于–80°C冰箱内暂存。待样品全部采集完成之后带回实验室提取总RNA。RNA提取采用常规TRIzol RNA提取方法进行, RNA提取完成后立即进行逆转录, 逆转录步骤参考RevertAidTMFirst StrandcDNA Synthesis Kit说明书。
1.2.3 引物设计及Mc-MT-10全长cDNA克隆 通过NCBI查找并下载已知的双壳贝类MT蛋白氨基酸序列并进行序列比对, 在保守区设计一对特异引物MT-F: ATGSCTGCACCTTGTAACTGYATY, MT-R:TCAYTTGCAGGARCANCCAGRTKC, 克隆Mc-MT-10基因部分 cDNA片段, 并送武汉转导生物科技有限公司进行Race测序。
1.2.4Mc-MT-10荧光定量分析 逆转录后的cDNA为模板稀释后作为Q-PCR的模板, 以厚壳贻贝β-actin为内参, MT-F、MT-R为引物进行Q-PCR实验, 分析Mc-MT-10组织表达谱及铜离子胁迫下的表达变化。
1.2.5 数据处理及分析 运用最小二乘法 2–ΔΔCt法(Livaket al, 2001)处理Q-PCR数据, 以溶解曲线判定扩增产物的特异性, 利用SPSS 13.0软件对数据进行分析, 采用单因子方差分析显著性差异。通过Oligin软件对数据进行处理并导出柱状图。
2 结果与分析
2.1 Mc-MT-10基因分析
经Race测序获得Mc-MT-10基因cDNA全长序列(图 1), 注册序列号为 KX987251。该序列全长为431bp, 包含 101bp的 5′-UTR 区, 108bp的 3′-UTR区,ORF区全长为 222bp, 推导其编码 73个氨基酸。预测其分子量为72.81kDa, 等电点为7.24。其分子式为C274H460N86O101S22, 含 943个原子。预测分子量大小和等电点分别为76.11kDa和9.027, 原子总数10747,分子式 C3365H5400N950O1005S27, 不稳定系数 44.34。用Clustal W 软件将Mc-MT-10氨基酸序列与其他物种MT-10序列进行比对(图2), 发现19个保守的Cys, 其中9对Cys与其相邻的氨基酸构成9个CXC结构。另外, CKCXXXCXCX结构也是软体动物金属硫蛋白常见的保守结构, 在Mc-MT-10蛋白序列中也发现存在一个类似结构域。

图1 厚壳贻贝MT-10基因cDNA及氨基酸序列Fig.1 The whole sequence of cDNA and deduced amino acid sequence of Mc-MT-10

图2 MT-10蛋白氨基酸多序列比对Fig.2 The multiple alignments of amino acid sequences of MT-10s
构建了基于Neighbor-joining的系统进化树, boot值为5000(图3)。在进化树中,Mc-MT-10首先与同属物种M.galloprovincialis的MT-10聚合在一起, 通过单序列比对,Mc-MT-10与Mg-MT-10核苷酸相似度为93.15%。而后与同属或科中的同源物聚合成一个大的分支, 与脊椎动物的相距较远。厚壳贻贝MT-10的进化地位与厚壳贻贝的生物学分类地位基本一致, 显示了较强的保守性。
2.2 Mc-MT-10基因组织表达分析
用qRT-PCR技术检测了Mc-MT-10基因在厚壳贻贝腮、肌肉、消化腺、外套膜、血细胞、性腺和肾脏等组织中的表达情况, 结果表明Mc-MT-10基因在各组织中均有表达, 在消化腺中的表达量最高, 其次为腮、血细胞和肾脏, 在肌肉、外套膜和性腺中的表达量最低(图4)。
2.3 铜离子胁迫下Mc-MT-10基因表达分析
用qRT-PCR技术检测了厚壳贻贝外套膜和消化腺Mc-MT-10基因在Cu2+胁迫下的表达情况(图5, 图6)。实验结果表明: 铜离子胁迫下, 外套膜和消化腺Mc-MT-10基因呈现类似的表达模式。处理 2d时,Mc-MT-10基因表达量显著升高, 5d时, 表达量达到峰值, 而后逐渐下降, 处理15d时, 消化腺Mc-MT-10基因表达量已回落至正常水平, 实验结束时(28d),外套膜Mc-MT-10基因表达量回落至接近正常水平。但两种组织Mc-MT-10基因表达量变化并不完全相同,外套膜中Mc-MT-10基因表达量变化显著高于消化腺。
3 讨论
金属硫蛋白一类广泛存在于生物界的低分子量蛋白, 富含半胱氨酸, 由于Cys巯基能共价结合金属离子, 是金属结合位点, 因此其对多种重金属, 如铜、镉及锌具有高度亲和性, 可参与生物体中对金属元素的解毒过程(Coyleet al, 2002)。本研究中, 厚壳贻贝MT-10蛋白含有19个Cys残基, 含量达26.0%,显示其发达的金属结合能力, 这与在其他软体动物的报道相一致(Engelkenet al, 1999; 吕达等, 2012)。CXC结构是软体动物MT蛋白的显著特征, 结果发现厚壳贻贝MT-10蛋白含有9个CXC结构, 经序列比对, 发现此类 CXC结构在软体动物 MT蛋白中的高度保守。有研究认为, 软体动物MT蛋白还存在保守的CKCXXXCXCX结构(Imagawaet al, 1990), 本研究发现厚壳贻贝 MT-10蛋白也存在一个这样的结构域, 同大部分软体动物的发现相一致, 但在海湾扇贝(Argopecten irradians)MT蛋白中却存在两个类似结构, 而且其包含145个氨基酸, Cys含量高达27.6, 显示了更强的重金属结合能力, 可能是两物种进化过程所处环境差异造成的,A.irradians在进化过程中所面对的重金属环境压力更大(刘维青等, 2006)。基于Neighbor-joining的系统进化树表明厚壳贻贝 MT-10的进化地位与厚壳贻贝的生物学分类地位基本一致,显示了较强的保守性。

图3 基于Neighbor Joining的系统进化树Fig.3 The phylogenetic tree constructed based on Neighbor Joining method
厚壳贻贝MT-10的组织表达谱表明, MT-10在各组织中是广泛存在的, 显示其广泛参与各类生命活动。而在消化腺以及腮、血细胞和肾脏中的表达量较高, 在软体动物中此类组织均为重要的免疫组织, 参与对外界刺激的应急、防御及免疫调控等, 进一步表明MT-10蛋白在厚壳贻贝中发挥重要的防御作用。
一些学者认为, 生物体内MT蛋白响应重金属胁迫呈现倒“U”型模式(Hermeszet al, 2001; Chanet al,2004; Wuet al, 2006), MT蛋白适应重金属胁迫有一个剂量阈值, 即当重金属达到某一浓度时, 金属硫蛋白会随重金属剂量增加而上升, 一旦超出该浓度阈值, 重金属就会对生物体产生不可逆的毒性, 金属硫蛋白水平就会随重金属水平增加而降低。本研究发现,随着处理时间的不断延长, MT-10蛋白的表达量逐渐升高, 到达峰值后逐步回落。在组织细胞吸收的铜离子剂量不断增加的情况下, 确实出现了典型的倒“U”型模式。研究结果表明, 在20μg/L铜离子胁迫下, 厚壳贻贝细胞内铜离子浓度阈值出现的时间在 5d, 5d后MT-10蛋白的表达量就逐渐下降, 15d后其表达量回落至正常水平。
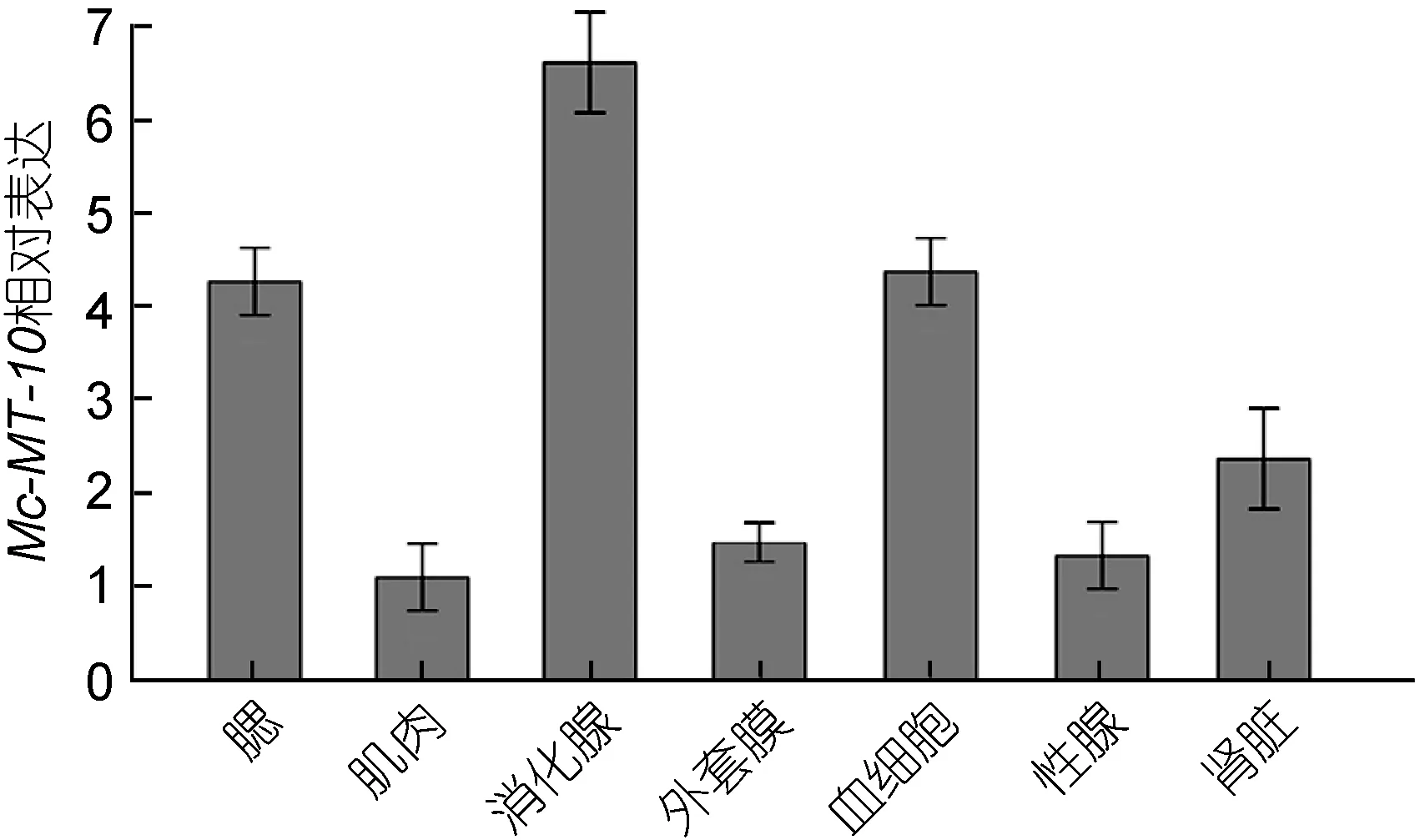
图4 厚壳贻贝MT-10 mRNA组织表达谱Fig.4 The tissues distributions of MT-10 mRNA in M.coruscus

图5 铜离子胁迫下厚壳贻贝外套膜组织MT-10 mRNA表达变化Fig.5 The expression analysis of MT-10 mRNA in mantle of M.coruscus under Cu2+ stress

图6 铜离子胁迫下厚壳贻贝消化腺组织MT-10 mRNA表达变化Fig.6 The expression analysis of MT-10 mRNA in digestive glands of M.coruscus under Cu2+ stress
4 结论
厚壳贻贝作为一种具有重要经济价值的海洋双壳贝类, 其作为环境指示生物的作用近年来也逐渐引起人们重视。本研究首次克隆了厚壳贻贝 MT-10蛋白全长cDNA序列, 分析其结构特征, 并对其响应重金属胁迫的 mRNA表达变化进行了初步研究。研究结果可为深入研究厚壳贻贝 MT蛋白的功能奠定基础, 为厚壳贻贝抗重金属污染新品种选育提供数据支持, 为降低贻贝养殖业风险提供了重要依据。
刘维青, 倪多娇, 宋林生等, 2006.海湾扇贝(Argopecten irradians)金属硫蛋白基因的克隆与分析.海洋与湖沼,37(5): 444—449
吕 达, 罗凯娅, 潘宝平等, 2012.青蛤(Cyclina sinensis)金属硫蛋白及硫氧还蛋白基因的克隆与表达分析.海洋与湖沼, 43(1): 47—51
郭远明, 刘 琴, 顾 捷等, 2012.4种海洋贝类对海水中铜(Cu)的富集能力.水产学报, 36(5): 708—713
常抗美, 吴剑锋, 2007.厚壳贻贝人工繁殖技术的研究.南方水产, 3(3): 26—30
Barka S, 2000.Processus de détoxication et localisation tissulaire des métaux traces (cuivre, zinc, nickel, cadmium, argent et mercure) chez un crustacé marinTigriopus brevicornis(Müller).Etude du biomarqueur “protéines type métallothionéines”, de la bioaccumulation des métaux et des conséquences sur le transfert trophique.Paris: Université de Paris
Chan P C, Shiu C K M, Wong F W Yet al, 2004.Common carp metallothionein-1 gene: cDNA cloning, gene structure and expression studies.Biochimica et Biophysica Acta-Gene Structure and Expression, 1676(2): 162—171
Coyle P, Philcox J C, Carey L Cet al.2002.Metallothionein: The multipurpose protein.Cellular and Molecular Life Sciences,59(4): 627—647
Engel D W, Brouwer M, 1993.Crustaceans as models for metal metabolism: I.Effects of the molt cycle on blue crab metal metabolism and metallothionein.Marine Environmental Research, 35(1—2): 1—5
Engelken J, Hildebrandt A, 1999.cDNA cloning and cadmium-induced expression of metallothionein mRNA in the zebra musselDreissena polymorpha.Biochemistry and Cell Biology, 77(3): 237—241
Hermesz E, ábrahám M, Nemcsók J, 2001.Tissue-specific expression of two metallothionein genes in common carp during cadmium exposure and temperature shock.Comparative Biochemistry and Physiology Part C:Toxicology & Pharmacology, 128(3): 457—465
Imagawa M, Onozawa T, Okumura Ket al, 1990.Characterization of metallothionein cDNAs induced by cadmium in the nematodeCaenorhabditis elegans.Biochemical Journal, 268(1): 237—240
Isani G, Serra R, Cattani Oet al, 1997.Adenylate energy charge and metallothionein as stress indices inMytilus galloprovincialisexposed to cadmium and anoxia.Journal of the Marine Biological Association of the United Kingdom,77(4): 1187—1197
Kim E K, Joung H J, Kim Y Set al, 2012.Purification of a novel anticancer peptide from enzymatic hydrolysate ofMytilus coruscus.Journal of Microbiology and Biotechnology,22(10): 1381—1387
Langston W J, Bebianno M J, Burt G R, 1998.Metal handling strategies in molluscs.In: Langston W J, Bebianno M J eds.Metal Metabolism in Aquatic Environments.Dordrecht:Springer, 219—283
Livak K J, Schmittgen T D, 2001.Analysis of relative gene expression data using Real-time quantitative PCR and the 2-ΔΔCtmethod.Methods, 25(4): 402—408
Miller B S, 1999.Mussels as biomonitors of point and diffuse sources of trace metals in the clyde sea area, Scotland.Water Science and Technology, 39(12): 233—240
Moore M N, Depledge M H, Readman J Wet al, 2004.An integrated biomarker-based strategy for ecotoxicological evaluation of risk in environmental management.Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis, 552(1—2): 247—268
Olsson P E, Kling P, Hogstrand C, 1998.Mechanisms of heavy metal accumulation and toxicity in fish.In: Langston W J,Bebianno M J eds.Metal Metabolism in Aquatic Environments.Dordrecht: Springer, 321—350
Roesijadi G, Fowler B A, 1991.Purification of invertebrate metallothioneins.Methods in Enzymology, 205: 263—273
Roesijadi G, Brubacher L L, Unger M Eet al, 1997.Metallothionein mRNA induction and generation of reactive oxygen species in molluscan hemocytes exposed to cadmium in vitro.Comparative Biochemistry and Physiology Part C:Pharmacology, Toxicology and Endocrinology, 118(2):171—176
Roeva N N, Sidorov A V, Yurovitskii Y G, 1999.Metallothioneins,proteins binding heavy metals in fish.Biology Bulletin, 26(6):617—622
Shuai J S, Wang L, 2001.Discussion about the health impact of heavy metal and the countermeasure.Environment and Exploitation, 16(4): 62
Wang Q, Ning X X, Chen L Let al, 2011.Responses of thioredoxin1 and thioredoxin-related protein 14 mRNAs to cadmium and copper stresses inVenerupis philippinarum.Comparative Biochemistry and Physiology Part C:Toxicology & Pharmacology, 154(3): 154—160
Wu S M, Jong K J, Lee Y J, 2006.Relationships among metallothionein, cadmium accumulation, and cadmium tolerance in three species of fish.Bulletin of Environmental Contamination and Toxicology, 76(4): 595—600
Ye Y Y, Li J, Wu C Wet al, 2012.Genetic analysis of mussel(Mytilus coruscus) populations on the coast of East China Sea revealed by ISSR-PCR markers.Biochemical Systematics and Ecology, 45: 1—6
THE CDNA CLONING AND CHARACTERIZATION OFMYTILUS CORUSCUSMETALLOTHIONEIN 10 AS WELL AS ITS MRNA EXPRESSION UNDER THE CU2+STRESS
WANG Hao-Sheng, SONG Xin-Jin, DONG Wen-Qiang, WU Bin,CHEN Yong-Xia, LU Wei-Xiao, QI Peng-Zhi, GUO Bao-Ying
(Zhejiang Ocean University,National Engineering Research Center of Marine Facilities Aquaculture,Zhoushan316022,China)
In this paper, a 413bp full-length cDNA sequence of metallothionein 10 from thick shell mussel was obtained with RT-PCR and rapid amplification of cDNA ends (RACE) technique (so called ‘Mc-MT-10’ thereafter).It consists of a 101bp 5′untranslated region (UTR), an 222bp open reading frame (ORF) and a 108bp 3′UTR.The translated protein is composed of 73 amino acids.The multiple alignments showed the abundant Cys residues, 9 conserved CXC domains and one CKCXXXCXCX domain, in addition, theMc-MT-10cluster into one branch with the counterparts from congeneric species in the phylogenetic tree, suggesting the high evolutionary conservation.The Q-PCR assays were employed to assess the relative expressive changes ofMc-MT-10mRNA in mantle and digestive gland under the stress of copper ions.The results showed that the expression levels ofMc-MT-10mRNA rose sharply under the stress of Cu2+,implying that theMc-MT-10could involve in the regulation of thick hell mussel against Cu2+stress.
Mytilus coruscus; metallothionein; Real-time fluorescence quantitative PCR; Cu2+stress
Q789; Q955
10.11693/hyhz20170300056
*浙江省公益项目, 2017C32009号; 浙江海洋大学科研启动资金项目, 21105013615号。王昊盛, E-mail: wanghaosheng1234@163.com
① 通讯作者: 祁鹏志, 博士, 助理研究员, E-mail: qpz2004@sina.com
2017-03-15, 收修改稿日期: 2017-04-03
