亚高山针叶林不同大小林窗植物多样性及其海拔动态
2017-12-11刘国华石松林
陈 力 ,刘国华 ,刘 丹 ,石松林
(1. 中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京100085;2. 中国科学院大学资源与环境学院,北京100049)
亚高山针叶林不同大小林窗植物多样性及其海拔动态
陈 力1,2,刘国华1,刘 丹1,2,石松林1,2
(1. 中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京100085;2. 中国科学院大学资源与环境学院,北京100049)
通过典型抽样法,分析了川西亚高山针叶林不同大小林窗中植物物种的更新组成及其沿海拔的动态变化,研究了林窗大小及林窗海拔对林窗内植物多样性的影响。结果表明:(1) 随林窗海拔的降低,乔灌层主要植物种垂直分布格局明显,林窗中植物种类组成逐渐增多,各物种更新数量逐渐增大,林窗越大,增加趋势越明显;(2) 在不同海拔、不同大小林窗中,岷江冷杉Abies faxoniana在乔木层物种重要值排序中始终位列第一,大叶金顶杜鹃Rhododendron faberisp、无柄杜鹃Rhododendron watsonii和华西箭竹Fargesia nitida重要值分别占据高、中、低海拔林窗中灌木层物种的首要位置,草本层主要物种排序有变化,但无明显规律;(3) 物种丰富度指数、Shannon-wiener物种多样性指数与林窗大小成极显著正相关,Simpson生态优势度与林窗大小呈极显著负相关,Pielou均匀度指数与林窗大小呈正相关但相关性不显著;(4) 低海拔林窗中植物多样性最高,其次为中海拔,高海拔林窗中的植物丰富度指数、Shannon-wiener物种多样性指数、Pielou均匀度指数和Simpson生态优势度指数与中低海拔之间存在显著差异。
林窗;大小;海拔;植物多样性;亚高山针叶林
林窗是森林更新的重要驱动力,是森林群落内植物更新的主要场所,对森林生物多样性的维持和生态系统的可持续发展具有重要影响[1-3]。林窗是森林生态系统中一种小尺度干扰[4],改善了林窗内部微生境,对森林内部资源进行了重新分配,为林窗中植物种子萌发和幼苗生长提供了可利用的能量和空间资源[5]。近年来,国内外学者研究发现,林窗大小对林窗中群落组成以及植物多样性具有重要影响。Adele等[6]在亚热带森林中研究发现,林窗大小对不同幼苗更新密度影响不一致,John等[7]阐述了温带森林中不同大小林窗对不同种植物存活率的影响,Zhang等[8]在中国北部半干旱地区研究发现,树高较高的沙地云杉Picea mongolica数量随林窗面积的增大而增多,而低矮的沙地云杉则相反,张建华等[9]对落叶松四种典型林分中进行择伐经营发现,一定的择伐强度能调整树种组成并进行近自然经营,崔宁洁等[10]以马尾松人工林为研究对象探讨了林窗大小对植物生活型和植物多样性影响以及季节性动态变化,认为植物多样指数在大体上表现为大林窗大于中小林窗,其中夏季植物多样性最高。关于不同大小林窗对植物更新和物种多样性影响的研究较多,但有关林窗中植物多样性沿海拔分布动态研究较少。
不同大小林窗中植物更新和多样性组成本身具有差异性,随着海拔的变化,环境因子也随之变化,从而影响林窗中种子的萌发、幼苗的生长以及土壤养分的供给,进而影响林窗中植物的组成和物种多样性沿海拔垂直分布格局[11-14]。本文以亚高山针叶林为研究对象,对林线下方3 000~3 700 m处,不同海拔、不同大小林窗中植物多样性进行了调查,分析了不同大小林窗中植物多样性指数及沿海拔的动态变化,旨在更好地了解研究林窗海拔和大小对植物多样性的影响,丰富林窗对植物多样性影响的研究,也为亚高山针叶林生态系统的保护、合理开发以及可持续发展提供理论依据。
1 研究地区与研究方法
1.1 研究区概况
研究区位于四川省卧龙自然保护区邓生保护站(102°56′~ 102°57′E,30°49′~ 30°50′N),海拔3 000~3 700 m,卧龙自然保护区位于岷江上游,地貌以高山深谷为主,属于典型的亚热带内陆山地气候,该区降水丰富,年降水量在861 mm,年均相对湿度高为80.3%;冬寒夏凉,年平均温为8.4℃,其中1月平均温度在-1.7℃,7月平均温度在17.0℃[15-16]。沿海拔变化,乔木层和灌木层植物群落种类分布明显,主要乔木层物种为:岷江冷杉Abies faxoniana、五尖槭Acer、糙皮桦Betula utilis等;灌木层主要物种为:大叶金顶杜鹃Rhododendron faberisp、黄花杜鹃Rhododendron lutescens、 无 柄 杜 鹃Rhododendron watsonii、 凝毛杜鹃Rhododendron phaeochrysum、华西箭竹Fargesia nitida等;主要草本有高山露珠草Circaea alpina、三角叶蟹甲草Parasenecio deltophyllus和白鳞酢酱草Oxalis acetosella等。
1.2 研究方法
1.2.1 样地设置及调查方法
2015年8月—9月,根据前期样地调查,在实验区内选择海拔3 000~3 700 m的连续样地内,通过典型抽样法,在海拔每100 m选取形状相似、年龄大小相近的不同大小林窗3~4个,调查林窗共25个,使用GPS(Magellan GPS315)测定各林窗海拔和地理坐标;使用罗盘仪测定坡向、坡位、林窗长轴方向以及形成木倒向;按照Runkle[17]和臧润国等[18]的定义对林窗内边界木种类、株数、胸径、高度、冠幅,形成木树种、数量、直径、地径、死亡原因以及腐烂程度进行调查;参考“PKU-PSD计划”标准[19], 对林窗内高度未到达2/3林冠层高大乔木(h≥1.5 m)进行每木检尺,记录其种类、高度、冠幅、胸径、地径、枝下高等,对低矮幼小乔木(h<1.5 m)进行调查,统计其种类、数量、株高、基径、冠幅、年龄等;对灌木层和草本层进行植物种类、株高、冠幅、株数(丛数)及盖度等指标调查。仅针对本研究,将林窗分为小林窗(0~100 m2)、中林窗(100~200 m2)、大林窗(200~405 m2),将林窗海拔分为高海拔林窗(3 000~3 200 m)、中海拔林窗(3 200~3 500 m)、高海拔林窗(3 500~3 700 m)。
1.2.2 数据计算
1)林窗面积 根据林窗不同形状特征,采用相应的方法进行计算,本研究中林窗形状大多数为不规则近椭圆,极个别为近圆形,针对不同形状林窗采用椭圆法和圆面积法分别计算林窗面积(A)。
圆面积法:A2=πR2。
式中:L为椭圆长轴;W为椭圆短轴。
2)重要值 衡量物种在森林群落中的作用和地位的综合指标[20-21]。本研究分别计算了各林窗中乔灌草层植物的相对密度、相对频度和相对显著度,最后求得重要值。
相对频度=某一物种的频度/所有物种的频度总和×100%。
相对密度=某一物种的株数/所有物种的株数总和×100%。
灌木和草本:
重要值=(相对频度+相对密度+相对显著度)/3。
式中:Si为第i个物种的胸高或基径断面积;Cri为第i个物种的盖度。
3)植物多样性指数[9,22]
物种丰富度指数:R=(S-1)/lnN。
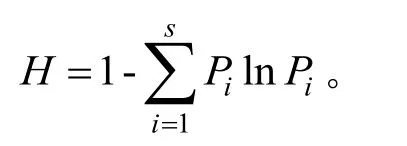
Simpson优势度指数:
式中:S为物种总数;N为林窗中个体数;Pi为第i个种占总个体数的比例。
1.2.3 数据处理与分析
通过野外调查数据,计算林窗中各物种重要值,统计各林窗中植物物种丰富度、Shannon-Wiener多样性指数、Pielou均匀度指数和Simpson优势度指数。采用Origin9.0软件作图。采用Excel2007和SPSS20.0进行数据统计分析,对不同大小不同海拔林窗间植物多样性采用单因素方差分析(One-way ANOVA)检验差异性,并进行LSD多重比较(α=0.05),采用Pearson相关性分析检验林窗大小、海拔以及环境因子与多样性指数的相关性。
2 结果与分析
2.1 不同大小林窗中物种更新及组成基本特征及沿海拔变化
野外调查数据显示,本研究区林窗中共有物种25科,44种,其中蔷薇科(7种)植物种类最多,之后杜鹃花科(4种)、百合科(4种)和菊科(3种)植物种类最多,然后为忍冬科(2种)、禾本科(2种)和荨麻科(2种)等(表1),分析发现:随林窗海拔的降低,乔木和灌木层物种组成变化明显,岷江冷杉为整个群落中乔木层的优势种,在中低海拔林窗中五尖槭和糙皮桦为共优种;中高海拔林窗中杜鹃花科植物为灌木层优势种,低海拔林窗中灌木层优势种为华西箭竹;草本层物种组成丰富,但除白鳞酢浆草、三角叶蟹甲草及高山露珠草等物种外其他种多为伴生种或随机种。总的来说,随林窗海拔的降低,林窗中植物种类组成逐渐增多,林窗中更新密度逐渐增大,且随林窗面积的增大,增加趋势越明显。
物种重要值反映了物种在群落中的优势地位以及对环境资源的利用能力,重要值越大,物种竞争力越强[9]。对林窗中乔灌草三层主要物种进行对比分析(表2),结果表明:在不同海拔、不同大小林窗内,岷江冷杉在乔木层主要物种的重要值排序中始终位列第一,沿林窗海拔降低岷江冷杉重要值呈下降趋势,说明对于林窗中乔木层其他物种来说,岷江冷杉具有较强的竞争力,而沿海拔的降低,林窗中的资源受到了其他物种的抢夺,林窗中岷江冷杉竞争压力增大;在不同海拔林窗中灌木层主要物种重要值排序不同,而相同海拔不同大小林窗内主要物种重要值排序不变,大叶金顶杜鹃、无柄杜鹃和华西箭竹重要值分别在高、中、低海拔林窗中位列第一,同一海拔灌木层物种重要值排序不随林窗大小变化而变化,说明海拔影响了灌木层物种的分布,与林窗海拔相对应的灌木层主要物种在同一海拔不同大小林窗中始终具有较强竞争力。不同海拔、不同大小林窗中草本层主要物种重要值排序均有所变化,但无明显规律,说明与乔灌层主要物种相比,草本层物种竞争力对林窗差异性环境条件响应更强烈。
2.2 不同大小林窗中植物多样性指数及沿海拔的变化
2.2.1 不同大小林窗中植物多样性变化
林窗大小是林窗的基本特征之一,影响着林窗内部光照、温度等微环境,进而影响着林窗内部物种更新和生物多样性[5,23]。并分别对不同大小林窗更新层物种多样性进行统计分析(图1)。结果表明:随着林窗的增大,物种多样性指数、物种丰富度指数呈上升趋势;物种均匀度指数在中等大小林窗中最大;物种优势度变化为:小林窗>大林窗>中林窗。单因素方差分析结果表明:不同大小林窗中植物多样性各指数之间存在显著差异。多重比较显示:大林窗中植物多样性各指数分别和中、小林窗中植物多样性各指数存在显著差异。相关性分析表明(表3):林窗大小与物种多样性指数、物种丰富度呈显著正相关,与物种优势度指数之间呈显著负相关,林窗大小与物种均匀度指数呈正相关但相关性不显著。说明林窗越大,林窗内植物种类越丰富,各物种数量越多,而林窗中物种优势度越低。
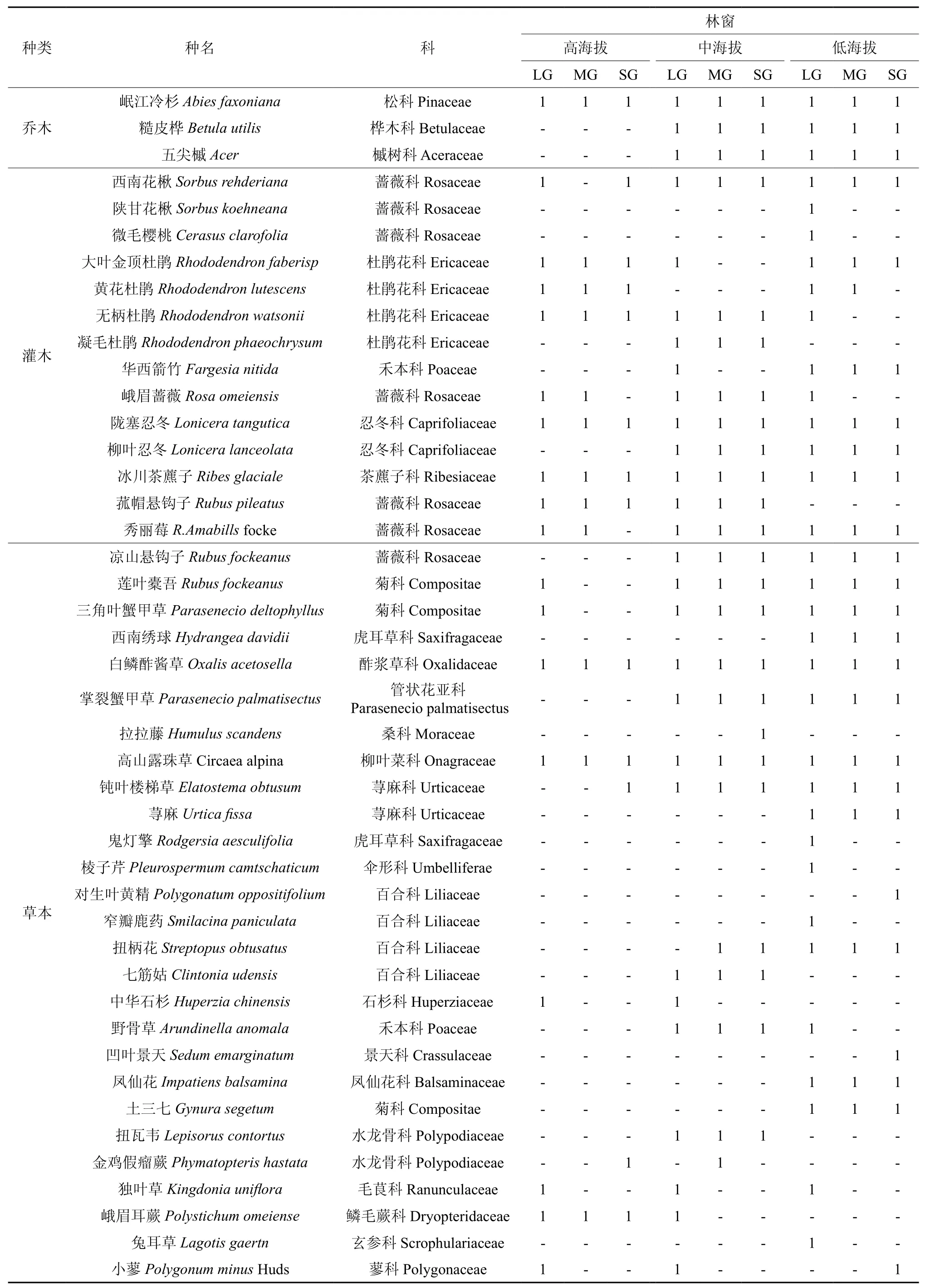
表1 亚高山针叶林不同大小林窗中物种沿海拔梯度分布†Table 1 The distribution of species in different gap size along the altitude in the subalpine coniferous forest
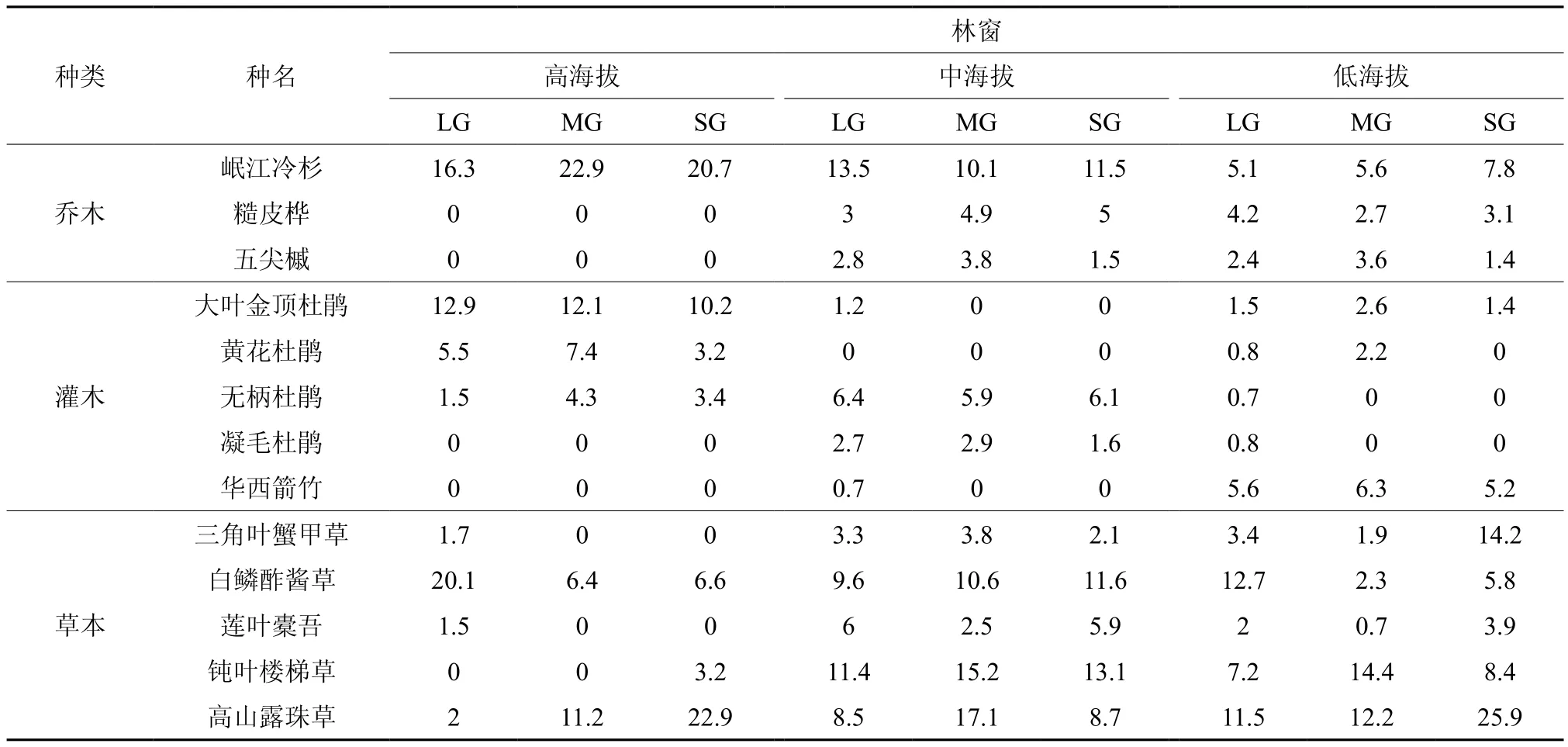
表2 不同大小林窗主要物种重要值沿海拔梯度的变化†Table 2 Importance value of main species in different size gap and altitude dynamic
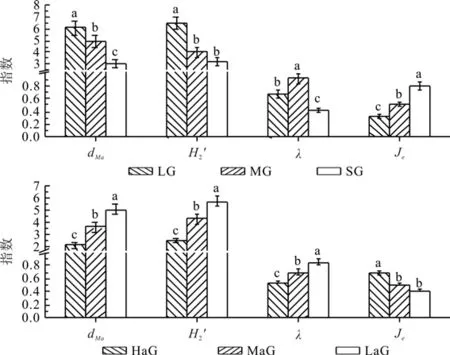
图1 不同大小、不同海拔林窗中植物多样性指数Fig.1 Plant diversity values in different size and altitude gaps
2.2.2 不同海拔林窗中植物多样性变化
林窗海拔是林窗最重要地形因子之一[24],随海拔的变化,林窗所处大环境发生显著变化,林窗内部生态因子也随之变化。对不同海拔林窗中植物多样性进行统计分析(图1)。结果表明:中、低海拔林窗中物种多样性指数、物种丰富度和物种均匀度指数大于高海拔林窗,但高海拔林窗中物种优势度指数要比中低海拔林窗中高。单因素方差分析表明,不同海拔林窗中植物多样性指数之间存在显著性差异。多重比较显示,海拔林窗中的物种多样性指数、物种丰富度、物种均匀度指数和物种优势度与中低海拔具有显著差异。相关性分析表明(表3):林窗海拔与物种多样性指数、物种丰富度和物种均匀度指数呈显著负相关,与物种均匀度指数呈显著正相关。说明低海拔林窗更适合维持物种多样性,林窗海拔对林窗内物种组成和物种数量具有显著影响。

表3 林窗大小、海拔与植物多样性指数的相关关系†Table 3 Correlation analysis of plant diversity indices and gap size and altitude
3 结论与讨论
在3 000~3 700 m处亚高山针叶林中不同大小林窗内,乔、灌木种类组成变化不大,但随林窗海拔的变化分布格局明显,而草本层物种在林窗中组成变化较大且无明显规律;大林窗中物种丰富度和物种多样性指数均高于中、小林窗,中林窗中均匀度指数最大,小林窗中优势度指数最大;林窗内植物多样性与物种丰富度随着海拔的降低而升高,因此亚高山针叶林林窗中植物的生长更新以及植物多样性除了受林窗大小的影响外,还与林窗海拔有关。
3.1 亚高山岷江冷杉原始林林窗物种组成及比较
研究区林窗中共发现植物24科,44种。大林窗中植物种类组成比小林窗更为丰富,说明大林窗更新层植物的生长和发展在群落组成和整个森林更新演替过程中占据了重要位置。随着海拔梯度的下降,乔木和灌木种类及数量发生了明显的变化,中低海拔林窗中乔木层植物种类比高海拔林窗种类多,优势种岷江冷杉分布于各海拔林窗中,共优种植物五尖槭和糙皮桦主要分布在中低海拔各林窗中,结合本研究区域海拔高度,这与大多数对木本植物种数沿海拔的分布格局的研究结果相似[25-27];沿海拔的变化,林窗中灌木层出现分层分布现象,高海拔林窗中以大叶金顶杜鹃和黄花杜鹃为主,中海拔林窗主要以无柄杜鹃和凝毛杜鹃,低海拔林窗中为华西箭竹,可能原因是研究区地处于较高海拔,具有较强的生境隔离度,从而引起了物种分布的破碎化[28-29],本研究中这种破碎化在乔木层主要物种分布上有体现,而在灌木层主要物种沿海拔梯度分布中表现最明显;沿海拔梯度的变化草本层主要植物种类和数量也发生相应的变化,但变化比较复杂,可能受林窗中其他环境因子以及种内种间竞争等因素的综合影响[29]。
随着海拔的变化,不同大小林窗中主要植物种的重要值也不同。乔木层植物中岷江冷杉重要值位列第一,在相同海拔不同大小林窗中岷江冷杉竞争力与资源利用能力变化不明显,随着海拔的降低岷江冷杉重要值变小,结合之前对物种沿海拔分布统计结果可知:岷江冷杉的生存竞争压力随海拔的而降低不断增大。灌木层主要物种重要值排序随林窗海拔的变化而变化,主要与灌木层植物种沿海拔梯度的分布格局有关。草本层主要物种重要值排序有变化,但无明显规律,这可能与草本层植物对林窗中多因素综合影响的环境适应能力有关。
3.2 林窗大小对植物多样性的影响
林窗的产生使得林内水、热等资源进行了再分配[30-31],林窗大小是林窗最重要的特征因子之一[32],对林窗内微环境及资源的分配状况有重要影响[33-34],与林窗内植物多样性维持具有密切的联系[5]。研究结果显示:同一海拔,大林窗中物种丰富度和物种多样性高于中、小林窗。林窗形成后,一方面大林窗为物种入侵提供了更多机会[35],另一方面林窗越大,其内部光照越充沛,地表温度随之上升,从而促进了土壤中种子的萌发和土壤有机质分解[36-38],这说明了大林窗为种子的萌发和生长提供了更为有利的环境条件和资源保障。不同大小林窗中植物均匀度指数大致表现为:中林窗>大林窗>小林窗,这主要与林窗中物种种类、数目以及分布有关,大林窗中物种种类和数量均较多,喜阳性物种和生长需光较多植物生态优势凸显,导致不同生物学特性物种个体数目的分配不均匀,而小林窗中物种种类和数量均相对较少,优势种和其他种个体数目分配也极不均匀,因此大林窗和小林窗内物种均匀度均低于中等大小林窗。不同大小林窗中物种优势度指数大致表现为:小林窗>大林窗>中林窗,这主要原因可能是小林窗内物种种类少,林窗资源环境对物种生存发展具有较强的选择性,导致小林窗内优势种占据林窗生境中的大部分资源,生态优势地位凸显,故小林窗中物种优势度较高;中林窗中物种优势度指数最低,从另一方面也反映了中林窗中物种均匀度最高;中林窗内物种均匀度最高,因此,物种优势度指数最小,这一点与吴昊对森林群落多样性指数对比研究的部分结果相吻合[39]。根据Haghverdi等[40]在伊朗北部的山毛榉森林中的研究发现,植物多样性指数与林窗面积呈线性变化,但本研究发现,物种丰富度、物种多样性和物种均匀度与林窗面积之间存在正相关,但并不是简单的线性相关,这一点和崔宁洁等在马尾松人工林中的研究相似。
3.3 林窗海拔对植物多样性的影响
在本研究区内,不同海拔林窗中物种多样性、物种丰富度、物种均匀度和生态优势度均存在显著差异,低、中海拔林窗中植物多样性明显高于高海拔。林窗中高大树木的缺失,使得林内资源进行了再配置,从而促进了林窗中植物种子的萌芽和较为幼小、低矮幼苗的生长发展[41],亚高山地区岷江冷杉树高随海拔的升高而变矮[16],林内郁闭度随海拔的升高而降低,而高海拔地区积雪时间长,生长季相对较短[42],因而高海拔地区林窗的出现不但没有很好地改善林下资源,增加种子的萌发率,反而增大了林窗内幼苗受外界干扰的几率及降低了幼苗幼树的存活率。中低海拔林内郁闭度高,林窗形成后促进了林窗内种子的萌发以及幼苗的生长,再加上中低海拔林窗中比高海拔林窗中更具适宜的温度、光照辐射等,为植物的繁殖和生长提供了良好、稳定的条件,因此中低海拔林窗中具有较高的物种多样性及丰富度,而高海拔林窗中均匀度指数较低而优势度指数最高,可能和高海拔林窗中优势种数量多而其他种数量较少有关。
本文从林窗大小以及海拔两种因素探讨了林窗对林窗内生物多样性的影响,为今后开展林窗与生物多样性之间相关关系的研究提供了一定的借鉴,然而林窗对植物更新和多样性维持的影响是一个十分复杂的生态学过程,沿海拔的变化不同大小林窗对植物更新和多样性的影响来自多方面,因此相关研究还需不断深入。另外,林窗中植物的更新是一个长期的动态过程,有关该区域林窗中植物更新以及多样性的时间变化动态还需进一步关注,因此本研究还存在一定的局限性。林窗中植物多样性除受林窗大小和海拔等林窗自身特征因子影响外,还有可能与其他地形因子(坡度、坡向等)、环境因子(温度、光照、辐射和土壤养分等)和生物因子(植物本身的生长特性、种内种间竞争、植物的化感作用以及动物的取食)有关,这有待于今后进一步的研究。
[1]Verma R, Kapoor K, Rawat R,et al.Analysis of plant diversity in degraded andplantation forests in Kunihar forest division of Himachal Pradesh[J]. IndiaJournal of Forestry, 2005,28(1):11-16.
[2]Dupuy J, Chazdon R. Interacting effects of canopy gap,understory vegetation and leaf litter on tree seedling recruitment and composition in tropical secondary forests[J]. Forest Ecology and Management, 2008, 255(11): 3716-3725.
[3]Elisa R, Dias E. Gap dynamics and regeneration strategies in Juniperus-Laurus forests of the Azores Islands[J]. Plant Ecology,2009, 200(2): 179-189.
[4]臧润国,徐化成. 林隙(GAP)干扰研究进展[J]. 林业科学,1998, 34(1): 90-98.
[5]李兵兵,秦 琰,刘亚茜,等. 燕山山地油松人工林林隙大小对更新的影响[J].林业科学, 2012, 48(6): 147-151.
[6]Adele M, Maria S, Silvio B,et al. Gap size effects on above- and below-ground processes in a silver fir stand[J]. European Journal of Forest Research, 2010,129(3): 355-365.
[7]John L, Willis, Michael B,et al. Scarification and gap size have interacting effects on northern temperate seedling establishment[J]. Forest Ecology and Management, 2015, 347:237-247.
[8]Zhang C, Zou C, Peltola H. The effects of gap size and age on natural regeneration of Picea mongolica in the semi-arid region of Northern China[J]. New Forests, 2013, 44(2): 297-310.
[9]张建华,张 宁,郭宾良,等. 木兰围场落叶松四种典型林分的经营模式初探[J]. 中南林业科技大学学报,2016,36(2):12-18.
[10] 崔宁洁,刘 洋,张 健,等. 林窗对马尾松人工林植物多样性的影响[J]. 应用与环境生物学报, 2014, 20(1): 8-14.
[11] Yan Q, Zhu J, Zhang J,et al.Spatial distribution pattern of soil seed bank incanopy gaps of various sizes in temperrate secondary forests, Northeast China[J]. Plant and Soil, 2010, 329(1-2): 469-480.
[12] 费世民,何亚平,何 飞,等. 关于森林林窗中几个问题的综述和展望[J]. 四川林业科技, 2009, 30(4): 28-37.
[13] 宋新章,肖文发. 林隙微生境及更新研究进展[J]. 林业科学,2006, 42(5):114-119.
[14] James G, Mathew L, Mark S,et al.Growth response by big-leaf mahogany(Swietenia macrophylla) advance seedling regeneration to overhead canopy release in southeast Pará, Brazil[J]. Forest Ecology and Management. 2005,204(2): 399-412.
[15] 郑绍伟,黎燕琼,何 飞,等. 卧龙森林生态站林内外主要气象因子对比分析[J]. 成都大学学报(自然科学版), 2006,25(3): 210-214.
[16] 何 飞,冯秋红,潘红丽,等. 四川卧龙岷江冷杉林分布规律及种群特征[J]. 四川林业科技. 2015,36(2):10-14.
[17] Runkle J. Gap regeneration in some old-growth forests of eastern United States[J]. Ecology, 1981, 62: 1041-1051.
[18] 臧润国,徐化成. 蛟河阔叶红松林林冠干扰及林隙更新研究[J]. 植物研究, 1999, 19(2): 112-120.
[19] 方精云. 探索中国山地植物多样性分布规律[J]. 生物多样性,2004, 12(1):1-4.
[20] Curtis J, Mcintosh R. An Upland Forest Continuum in the Prairie-Forest Border Region of Wisconsin[J]. Ecology, 1951,32(3): 476-496.
[21] 王育松,上官铁梁. 关于重要值计算方法的若干问题[J]. 山西大学学报(自然科学版), 2010, 33(2): 312-316.
[22] 朱教君,刘世荣. 森林干扰生态研究[M]. 北京: 中国林业出版社, 2007.
[23] 刘文杰,李庆军,张光明,等. 西双版纳望天树林干热季不同林窗间的小气候差异[J]. 生态学报, 2000,20(6):749-755.
[24] 张志国,马遵平,刘何铭,等. 天童常绿阔叶林林窗的地形分布格局[J]. 应用生态学报, 2013,24(3):621-625.
[25] 陈廷贵,张金屯. 山西关帝山神尾沟植物群落物种多样性与环境关系的研究I.丰富度、均匀度和物种多样性指数[J]. 应用与环境生物学报, 2000,6(5):406-411.
[26] 郝占庆,于德永,杨晓明,等. 长白山北坡植物群落α多样性及其随海拔梯度的变化[J]. 应用生态学报, 2002,13(7):786-789.
[27] 岳 明,张林静,党高弟,等. 佛坪自然保护区植物群落物种多样性与海拔梯度的关系[J]. 地理科学, 2002,22(3):349-354.
[28] Kessler M. The elevational gradient of Andean plant endemism:varying influences of taxon-specific traits and topography at different taxonomic levels[J].Journalof biogeography, 2002, 29:1159-1165.
[29] 唐志尧,方精云. 植物物种多样性的垂直分布格局[J]. 生物多样性, 2004, 12(1):20-28.
[30] 喻阳华,李 飒,严令斌,等. 赤水河上游次生林林窗特征及森林更新[J]. 森林与环境学报, 2015, 35(3):265-271.
[31] 吴庆贵,吴福忠,谭 波,等. 高山森林林窗对凋落叶分解的影响[J]. 生态学报, 2016,36(12):1-9.
[32] 胡理乐,李俊生,吴晓莆,等. 林窗几何特征的测定方法[J].生态学报, 2010, 30(7): 1911-1919.
[33] 王金铃,段文标,陈立新,等. 云冷杉林风倒区林隙和掘根微立地微气候变化[J]. 林业科学研究, 2015, 28(2): 173-182.
[34] 李苏闽,游巍斌,肖石红,等. 天宝岩长苞铁杉林林窗的微环境特征[J]. 森林与环境学报, 2015, 35(4): 343-350.
[35] Yan Q, Zhu J, Yu L. Seed regeneration potential of canopy gaps at earlyformation stage in temperate secondary forests, Northeast China[J]. Plos One, 2012, 7(6): 1495.
[36] 段文标,王丽霞,陈立新,等. 红松阔叶混交林林隙大小及光照对草本植物的影响[J]. 应用生态学报,2013,24(3):614-620.
[37] Whitmore T. Canopy gaps and the two major groups of forest trees[J]. Ecology, 1989, 70(3): 536-538.
[38] 张 敏,朱教君,闫巧玲. 光对种子萌发的影响机理研究进展[J]. 植物生态学报, 2012, 36(8): 899-908.
[39] 吴 昊. 不同类型群落物种多样性指数的比较研究[J]. 中南林业科技大学学报, 2015, 35(5): 84-89.
[40] Haghverdi K, Kiadaliri H, Sagheb-Talebi,et al.Variability of Plant Diversity and Soil Features Following Gap Creation in Caspian Beech Forests of Iran[J]. Annalsof Biological Research,2012, 3(9): 4622-4635.
[41] 韩文娟,袁晓青,张文辉. 油松人工林林窗对幼苗天然更新的影响[J]. 应用生态学报, 2012,23(11): 2940-2948.
[42] 程 伟,吴 宁,罗 鹏. 岷江上游林线附近岷江冷杉种群的生存分析[J]. 植物生态学报, 2005,29(3):349-353.
[本文编校:吴 彬]
Plant diversity and elevation dynamics in forest gaps of varying sizes in subalpine coniferous forest
CHEN Li1,2, LIU Guohua1, LIU Dan1,2, SHI Songling1,2
(1. State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-environmental Sciences, Chinese Academy of Sciences,Beijing 100085, China; 2. School of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100049, China)
Through typical sampling method, this paper analyzed the composition and updated dynamic changes along altitude gradients,studied the effects of gap size and elevation to plant species in gaps. The results indicated: (1) with the decrease of forest gap elevation,the pattern of main plants in trees layer and shrubs layer in forest gaps distributed obvious, plant species composition and update quantity in forest gaps increased gradually. What’s more, the greater the gaps, increasing trend more obvious; (2) In different size gaps along altitude gradients,Abies faxonianaalways ranked first in tree layer species significant value sort, significant value ofRhododendron faberisp,Rhododendron watsoniiandFargesia nitidarespectively occupied the first place at high, middle and low altitude in forest gap shrub species, the significant value of main species ordering in herb layer is mutative, but without obvious regularity; (3) There is a very significant positive correlation between forest gap size and species richness index and Shannon wiener diversity index, there is a very significant negative correlation between forest gap size and Simpson ecological dominance index, there is a positively related between forest gap size and Pielou evenness index, but had no significant correlation; (4) The highest plant diversity in forest gap at the low altitude, the medium altitude followed, there is significant differences between forest gaps at high altitude and medium altitude and low altitude of plant richness index, Shannon-wiener diversity index, Pielou index and Simpson ecological dominance index.
Forest gap; Size; Elevation; Plant diversity; subalpine coniferous forest
S718.54 文献标志码:A 文章编号:1673-923X(2017)10-0090-08
10.14067/j.cnki.1673-923x.2017.10.015
http: //qks.csuft.edu.cn
2016-06-02
国家重点研发计划专项(2016YFC0502102)
陈 力,硕士研究生
刘国华,研究员,博士生导师;E-mail:ghliu@rcees.ac.cn
陈 力,刘国华,刘 丹,等. 亚高山针叶林不同大小林窗植物多样性及其海拔动态[J].中南林业科技大学学报,2017, 37(10):90-97.
