光皮树不同无性系果期光合日变化研究
2017-12-11吴林瑛邓文剑张应中邓智曦张良波何祯祥王明怀
吴林瑛 ,邓文剑 ,张应中,林 军 ,邓智曦 ,张良波 ,何祯祥 ,王明怀
(1.广东省森林培育与保护利用重点实验室,广东 广州 510520;2. 广东省林业科学研究院,广东 广州510520;3.乐昌市龙山林场,广东 乐昌 512221;4.湖南省林业科学院,湖南 长沙 410004;5. 湖南省生物柴油工程技术研究中心,湖南 长沙 410004;6. 南京大学 生命科学学院,江苏 南京 210093)
光皮树不同无性系果期光合日变化研究
吴林瑛1,2,邓文剑3,张应中1,2,林 军3,邓智曦3,张良波4,5,何祯祥6,王明怀1,2
(1.广东省森林培育与保护利用重点实验室,广东 广州 510520;2. 广东省林业科学研究院,广东 广州510520;3.乐昌市龙山林场,广东 乐昌 512221;4.湖南省林业科学院,湖南 长沙 410004;5. 湖南省生物柴油工程技术研究中心,湖南 长沙 410004;6. 南京大学 生命科学学院,江苏 南京 210093)
为研究光皮树光合特征的日变异规律,采用美国Li-6400便携式光合测定仪,对光皮树8个无性系8月份果期叶片的光合日变化进行了测定和分析,结果表明:(1)参试光皮树无性系净光合速率(Pn)、气孔导度(Gs)日变化均呈单峰曲线,多数无性系最大值出现在9:00左右;蒸腾速率(Tr)日变化除JX13和GY8040外,均呈双峰曲线,第一个峰值高于第二个峰值;胞间CO2浓度(Ci)日变化曲线为扁平的“W”形;水分利用效率(WUE)日变化为不规则“V”形,谷底在15:00左右。(2)8个光皮树无性系净光合速率、蒸腾速率、气孔导度、胞间CO2浓度、水分利用效率日均值间差异不显著,其中GY8040、JX3、LC7、JX6的净光合速率(Pn)较高,均达到10.0 μmol·m-2s-1以上。(3)环境因子中,光照强度(PPAR)直接或通过大气温度(Ta)、大气湿度(HR)影响植物光合生理,大气CO2浓度(Ca)仅与胞间CO2浓度(Ci)呈显著正相关。净光合速率(Pn)与气孔导度(Gs)和蒸腾速率(Tr)呈极显著正相关,各因素对光皮树净光合速率(Pn)影响从大到小依次为气孔导度>蒸腾速率>光照强度>大气湿度>大气温度>水分利用效率>胞间CO2浓度>大气CO2浓度。
光皮树;无性系;光合日变化;环境因子;相关性
光皮树Cornus wilsoniana,又名光皮梾木,山茱萸科Cornaceae梾木属落叶乔木或灌木,广泛分布于黄河以南地区,适应性强,根系发达,生长速度较快。光皮树是高产优质的食用油料及生物质能源树种,大树产果量高(每株年产干果50 kg以上),干果含油率达33%~36%[1]。长期食用光皮树油能有效治疗高血脂,降低胆固醇[2]。作为一种优良的生物柴油,光皮树油的燃料特性和动力性能接近0号柴油[3-4]。近年来光皮树在油料成分分析及制取[5-6]、栽培及良种选育[7]等方面开展了重点研究。在光合生理方面,目前宋庆安等[8-9]、梁文斌等[10]针对湖南当地优良光皮树无性系进行了光响应、不同无性系开花期光合日变化、气孔及光合特性比较等研究,指出所选当地无性系光合特征值的日变异规律,并进行了无性系间光合参数日均值聚类以及差异性和相关性分析,为光皮树光合生理研究奠定了良好的基础。由于种源及分布地的不同,大部分树种的光合特性表现出很明显差异[11],故而本研究以光皮树分布区南部广东初选的乐昌、江西种源优良无性系为材料,开展光合日变化测定研究,旨在了解广东光皮树的光合日变化特征、生长环境与光合特性之间的互作,揭示本地光皮树光合作用的基本生理生态学规律,分析其生长的适宜生态条件,为光皮树优良无性系的选择、丰产栽培技术措施的制定提供科学依据。
1 材料与方法
1.1 试验地概况
试验苗木位于广东省乐昌市龙山林场良种苗木基地。地理位置为 25°11′N ,113°30′E,属中亚热带季风气候,年降水量为1 550~1 750 mm,全年平均气温为19. 6 ℃,极端最低气温为-4.6 ℃,极端最高气温为38.4 ℃,无霜期304 d 以上。
1.2 试验材料
试验材料为6年生光皮树无性系幼树,共计8个,分别为来自江西的JX2、JX3、JX6、JX7、JX13和来自广东乐昌的LC3、LC7、GY8040。每个无性系选取株高、地径接近的5株苗木进行测定。平均株高为3.5 m,胸径为4.5 cm,冠高为3.0 m,冠幅为2 m×2 m。
1.3 试验方法
采用美国Li-6400便携式光合测定仪,于2015年8月中旬,晴朗天气选取生长良好的中上部成熟叶片进行瞬时光合速率测定。光合日变化分别在 7:30、9:00、10:30、12:00、13:30、15:00、16:30、18:00各测1次,先测定自然光照强度,然后将红蓝光源设定与自然光相同的光强,同一时间段光强相同,测定时用缓冲瓶控制CO2浓度。光合仪可以自动记录每一时刻测定的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)以及环境因子如光照强度(PPAR)、大气温度(Ta)、大气湿度(HR)、大气CO2浓度(Ca)等参数。每个无性系选择5个样株,每个样株测定2片叶子,每片叶子记录5次,每个无性系1天仅能测2样株,共测定3日。分析数据时取平均值。
1.4 数据处理
本研究数据整理和分析采用Excel 2007和SPSS17.0。叶片水分利用效率(WUE,EWUE)由公式EWUE=Pn/Tr计算获得。
2 结果与分析
2.1 光皮树参试无性系光合参数日变化规律及环境因子的日变化
由图1可知,在测定范围内,参试的8个光皮树无性系净光合速率(Pn)日变化均呈单峰曲线,不存在“光合午休”现象。多数无性系Pn值在9:00左右达到最大,随后缓慢下降, 只有LC3的最大值出现在12:00左右,13:30之后各无性系净光合速率急速下降,18:00为观测范围内的最低值。各个无性系整个净光合速率日变化过程中,早晨的Pn值高出傍晚约4.5倍。
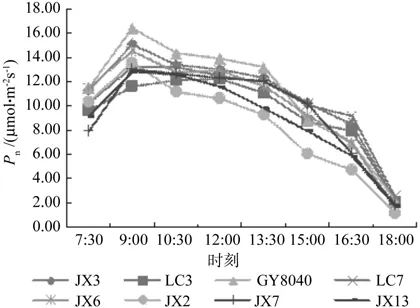
图1 光皮树无性系净光合速率(Pn)日变化Fig.1 The diurnal variation of net photosynthetic rate (Pn)of Cornus wilsoniana clones
由图2(左)可以看出,光皮树不同无性系气孔导度(Gs)日变化与净光合速率有相似的变化趋势,呈单峰曲线,早晨的Gs平均值高出傍晚约5.8倍。在观测范围内,JX6、JX2峰值在7:30左右,其他峰值主要出现在7:30—10:30;GY8040、JX3、JX7、LC3、LC7峰值在9:00左右,JX13峰值在10:30左右;之后整体缓慢下降,13:30之后急速下降。总体上来看,光皮树各无性系间气孔导度日变化趋势相对分散。
由图2(右)可以看出,不同光皮树无性系胞间CO2浓度(Ci)日变化曲线呈偏平“W”形:10:30时出现第一个低谷,Ci整体均值为242 μmol·mol-1;随后逐渐升高,13:30—15:00出现峰值,16:30 出现第二个低谷,Ci均值为 240 μmol·mol-1,比第一个谷值略低,之后回升。傍晚各无性系间Ci均值比早晨低 18.7 μmol·mol-1。

图2 光皮树无性系气孔导度Gs(左)和胞间CO2浓度Ci(右)日变化Fig.2 The diurnal variation of stomatal conductance(Gs) and intercellular concentration (Ci) of Cornus wilsoniana clones
由图3(左)可以看出,蒸腾速率(Tr)日变化方面,JX13和GY8040为单峰曲线,峰值出现在12:00。其他无性系均呈双峰曲线:上午蒸腾随着光照不断增加,9:00达到第一个峰值,10:30出现第一个低谷;随后在12:00达到第二个峰值,其中JX2第二个峰值在13:30。整体来看,第二个峰值大于第一个峰值。13:30以后,参试8个光皮树无性系的蒸腾速率均出现迅速下降的趋势。8个无性系的蒸腾速率最大值均出现在12:00—13:30之间,最高值为 5.0 mmol·m-2s-1。
水分利用效率表示每损失单位数量的水所固定的CO2的数量。由图3(右)可知,各无性系的水分利用效率在早晨较高,9:00之后逐渐降低,15:00 左右达到谷底,均值为 2.3 mmol·mol-1。在测定范围内,JX3、GY8040、LC7、JX13、JX2、JX7的水分利用效率值18:00升至最高,JX6、LC3两个无性系在16:30出现峰值,之后降低。早晨水分利用效率均值为3.6 mmol·mol-1,略高出傍晚,二者相差4.2%。
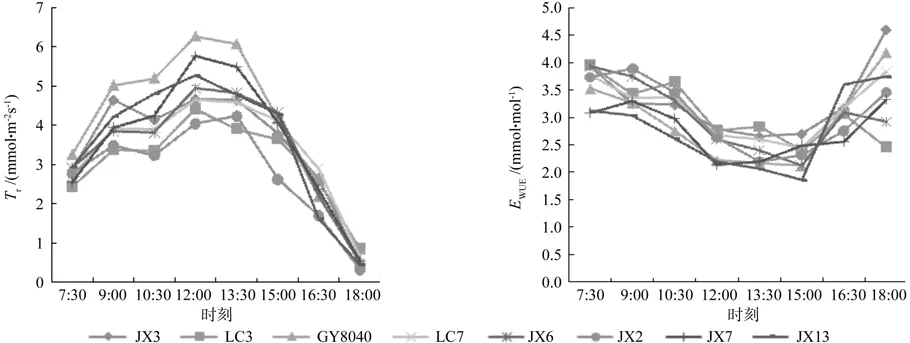
图3 光皮树无性系蒸腾速率(左)和水分利用效率(右)日变化Fig.3 The diurnal variation of transpiration rate (Tr) and water use efficiency (WUE) of Cornus wilsoniana clones
对光皮树光合速率日变化测定的同时,记录了相应时刻环境因子的变化情况(见图4)。光照强度从早晨开始增加,由486 μmol·m-2s-1猛增至9:00的 1 276 μmol·m-2s-1,至 10:30 增幅放缓,之后 12:00左右达到最大值,为 1 877 μmol·m-2s-1,15:00 之后急速下降,18:00 仅为 100 μmol·m-2s-1。大气 CO2浓度(Ca)9:00之后急速下降,之后略有波动,整体表现出“两边高,中间低”的变化趋势。大气温度(Ta)清晨时最低,中午达到高峰35℃左右,傍晚回落,温差4℃左右。大气湿度从清晨开始逐渐下降,12:00达到最低值,之后缓慢回升。

图4 环境因子日变化Fig.4 The diurnal variation of environment factors
2.2 光皮树不同无性系间光合特性差异
由表1可知,各无性系间光合参数P值均大 于 0.05, 表示 JX2、JX3、JX6、JX7、JX13、LC3、LC7、GY8040 8个光皮树无性系之间在净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)和水分利用效率日均值方面数据一致性较强,无显著差异。
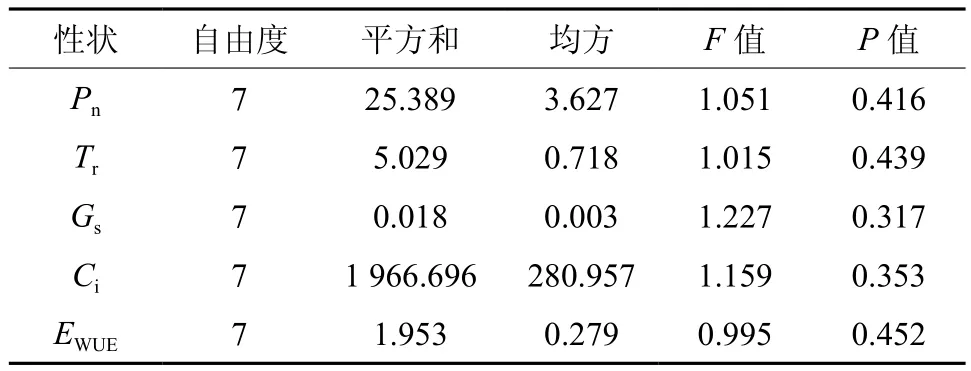
表1 光皮树8个无性系间光合参数日均值的方差分析†Table 1 Variance analysis of photosynthetic parameter on 8 Cornus wilsoniana clones
由图5可以看出,光皮树净光合速率(Pn)日均值较高的无性系有GY8040、JX3、LC7、JX6,均达到 10.0 μmol·m-2s-1以上;LC3、JX7 均为 9.5 μmol·m-2s-1,处中等水平;JX2、JX13略低于 9.0 μmol·m-2s-1。总体上8个光皮树无性系间净光合速率日均值相差不大,极差为 2.5 μmol·m-2s-1。
由图5可知,光皮树气孔导度(Gs)日均值较高的无性系是GY8040,可达到0.21 mol·m-2s-1;LC7、JX3、JX7、JX13的Gs值相同,均为0.17 mol·m-2s-1,JX6、LC3 略小,为 0.16、0.15 mol·m-2s-1,气孔导度均属中等;JX2较低,为0.13 mol·m-2s-1。
图5反映的是光皮树胞间CO2浓度(Ci)日均值大小比较,不同无性系间差异较大,极差为22 μmol·mol-1。其中 JX7、JX13、GY8040 的Ci值较高,在 270 μmol·mol-1左右,JX6、JX2、JX3、LC7 中等,LC3 的Ci值最小,为 249 μmol·mol-1。
图5蒸腾速率(Tr)的日均值比较来看,8个无性系间差异不大,GY8040蒸腾速率最高,Tr值为 4.09 mmol·m-2s-1,LC3、JX2最低,分别为3.07、2.80 mmol·m-2s-1。其他无性系处于中等水平,Tr值在 3.50 mmol·m-2s-1左右。
由图5可知,参试的8个光皮树无性系的水分利用效率日均值较为一致,大小在3.00 mmol·mol-1左右,最高为 3.29 mmol·mol-1,最低为2.76 mmol·mol-1,极差仅为 0.53 mmol·mol-1。
2.3 光皮树各光合参数及环境因子间的相关性分析

图5 光皮树8个无性系间各光合参数日均值的比较Fig.5 The comparison of daily average values of photosynthetic parameter in Cornus wilsoniana clones
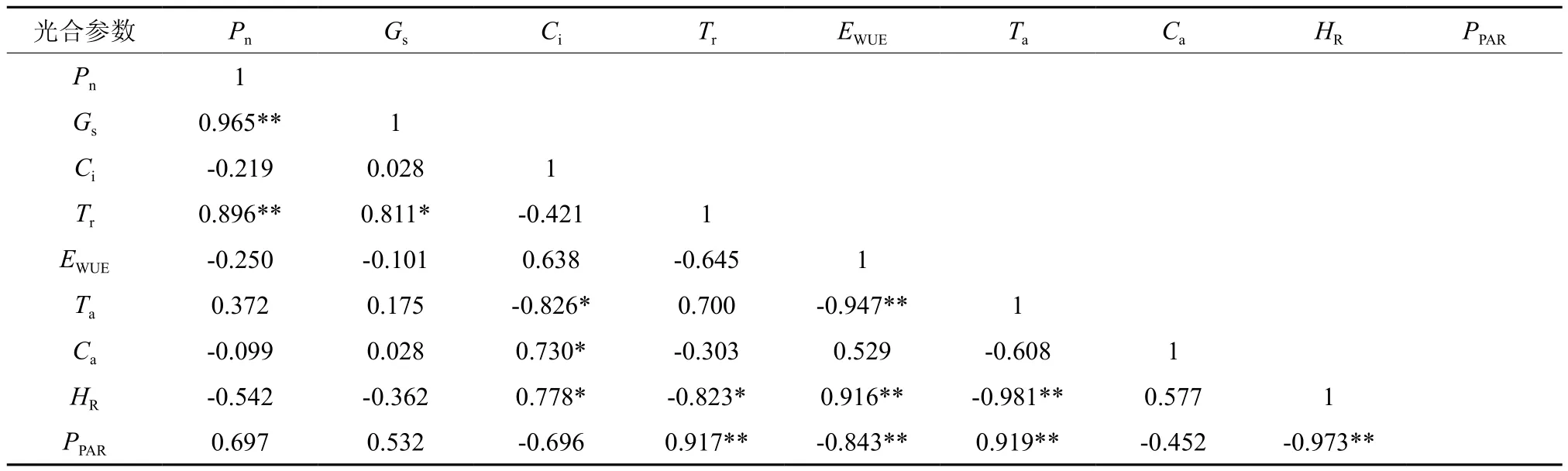
表2 光皮树光合参数与环境因子的相关分析†Table 2 Correlation analysis of environment factors and photosynthetic parameter in Cornus wilsoniana clones
以各无性系不同时刻光合参数平均值与相应即时环境因子进行相关性分析,结果见表2。光皮树净光合速率与气孔导度和蒸腾速率呈极显著正相关(P<0.01),相关系数分别高达0.965、0.896;净光合速率与光照强度和外界大气温度呈正相关,相关系数为0.697、0.372,未达到显著水平;净光合速率与胞间CO2浓度、水分利用效率、空气中CO2浓度、空气相对湿度呈现一定的负相关;各因素对光皮树Pn影响从大到小依次为:气孔导度>蒸腾速率>光照强度>大气湿度>大气温度>水分利用效率>胞间CO2浓度>大气CO2浓度。光皮树叶片气孔导度与蒸腾速率呈显著正相关;胞间CO2浓度与大气CO2浓度呈显著正相关,而与大气温度呈显著负相关。光照强度对光皮树光合的影响中表现出与大气温度、蒸腾速率呈极显著正相关,相关系数分别为0.919、0.917;与相对湿度、水分利用效率呈极显著负相关,相关系数分别为-0.973、-0.843;空气相对湿度与水分利用效率、胞间CO2浓度呈极显著正相关,相关系数分别为0.916和0.778,与大气温度、蒸腾速率呈极显著负相关,相关系数依次为-0.981、-0.823;大气温度与水分利用效率极显著负相关,相关系数达0.947,与胞间CO2浓度显著相关;大气CO2浓度仅与胞间CO2浓度呈显著相关,即空气中CO2浓度显著影响叶片胞间CO2浓度。
3 结论与讨论
通常用净光合速率反映植物光合能力的大小,自然条件下,植物光合作用的日变化曲线有单峰、双峰和不规则3种类型[12]。本研究中,8个参试光皮树无性系净光合速率日变化于8月份果期所测均呈单峰曲线,多数无性系最大值出现在9:00左右,之后逐渐下降。这与宋庆安等[9]以湖南光皮树无性系为材料于5月份测定的结果较为相近:12个无性系中8个呈单峰曲线,且多数无性系一天之内的峰值出现在上午较早时候,比如9:00。本研究中无性系净光合速率日均值在10 μmol·m-2s-1左右,整体高于湖南光皮树无性系。蒸腾作用是植物吸收和运输水分的主要驱动力[13]。本研究中不同光皮树无性系蒸腾速率一天内呈现不同的变化模式,蒸腾速率日均值(3.5 mmol·m-2s-1)也略高于湖南光皮树无性系(3.0 mmol·m-2s-1),可能是由测定时节、环境因子、生长阶段及不同无性系遗传背景差异所致。气孔是植物叶片与外界进行气体交换的通道,气孔导度是气体及水分进出难易程度的指标。胞间CO2浓度是指植物气孔下腔或叶肉细胞间隙CO2浓度,胞间CO2浓度的变化方向是确定光合速率变化的主要原因是否为气孔因素的必不可少的判断依据[14-15]。梁文斌等[10]对光皮树不同无性系苗木气孔及光合特性比较结果表明,光皮树不同无性系之间净光合速率、蒸腾速率、气孔导度存在显著差异,而无性系之间的胞间CO2浓度和水分利用效率的差异不显著。在本研究中所选8个光皮树无性系之间净光合速率、蒸腾速率、气孔导度、胞间CO2浓度和水分利用效率差异均不显著,可能因为所选无性系之间遗传背景较为相近,光合生理特性相差不大,从而表现较为整齐一致的光合日变化规律。植物光合生理指标的变化与自身因素和环境因子密切相关[16]。 净光合速率与蒸腾速率、气孔导度呈极显著正相关,与梁文斌等[10]的结论一致,较高的蒸腾速率和气孔导度有利于CO2和水分的供应,通过保持较高的气孔导度来保证较高的碳同化速率,提高光合效率。
本研究对8个光皮树无性系8月份果期叶片的光合日变化测定分析可以得出的结论有:参试光皮树无性系净光合速率、气孔导度日变化均呈单峰曲线,多数无性系最大值出现在9:00左右;蒸腾速率日变化除JX13和GY8040外,均呈双峰曲线,第一个峰值高于第二个峰值;胞间CO2浓度日变化曲线为扁平的“W”形;水分利用效率日变化为不规则“V”形,谷底在15:00左右。8个光皮树无性系在净光合速率、蒸腾速率、气孔导度、胞间CO2浓度、水分利用效率日均值之间差异不显著,其中GY8040、JX3、LC7、JX6的净光合速率值较高,均达到10.0 μmol·m-2s-1以上。环境因子中,光照强度直接或通过大气温度、大气湿度影响植物光合生理,大气CO2浓度仅与胞间CO2浓度呈显著相关。净光合速率与气孔导度和蒸腾速率呈极显著正相关,各因素对光皮树净光合速率影响从大到小依次为:气孔导度>蒸腾速率>光照强度>大气湿度>大气温度>水分利用效率>胞间CO2浓度>大气CO2浓度。
光合作用是植物物质生产的基础,也是全球碳循环及其它物质循环的重要环节[17]。光合速率日变化的研究,有助于环境因子与光合特性之间的互作关系,了解其光合日变化的特征规律,揭示其光合作用的基本生理生态学特征,分析生长的适宜生态条件,为制定丰产栽培技术措施和扩大栽培提供科学理论依据。
[1]梁仰贞. 光皮树的栽培技术[J]. 中国土特产,1997,(2):13-14.
[2]成训妍. 光皮树是珍贵的木本食用油料资源[J].生物与特产,1990, (6): 28 .
[3]李昌珠, 蒋丽娟, 程树棋. 生物柴油研究现状与商业化应用前景[C]//中国生物质能技术研讨会论文集. 南京: 2002 .
[4]李正茂, 邓新华, 李党训. 光皮树经济性状及生物质液体燃料开发研究构想[J]. 湖南林业科技, 1996, 23(2): 11-13.
[5]刘汝宽, 肖志红, 李昌珠, 等. 光皮树果实内含物含量的测定与综合利用[J]. 中南林业科技大学学报, 2013, 33(12): 19-21.
[6]申爱荣, 谭著明, 蒋丽娟,等. 不同提油方法对制取光皮树油的影响[J]. 中南林业科技大学学报, 2010, 30(11): 129-135.
[7]李昌珠, 张良波, 李培旺. 油料树种光皮树优良无性系选育研究[J]. 中南林业科技大学学报, 2010, 30(7): 1-8.
[8]宋庆安, 李昌珠, 童方平, 等.光皮树优良无性系光合生理特性对光强的响应[J]. 湖南林业科技, 2008, 35(6): 3 -6 .
[9]宋庆安, 李志辉, 李昌珠, 等. 光皮树不同无性系开花期光合生理特性日变化规律研究[J]. 中国农学通报, 2009, 25(10):143-147, 7.
[10]梁文斌, 蒋丽娟, 刘 强, 等. 光皮树不同无性系苗木气孔及光合特性比较[J]. 经济林研究, 2011, 29(2): 42-4.
[11]蒋 燚, 李志辉, 朱积余, 等. 不同红锥种源幼林光合特性日变化测定与分析[J].中南林业科技大学学报,2013,33(6):43-47.
[12]梁开明, 曹洪麟, 徐志防, 等. 台湾青枣及野生种的光合作用日变化及光响应特征[J]. 园艺学报, 2008, 35(6): 793-798.
[13]李合生. 现代植物生理学[M]. 北京: 高等教育出版社, 2002.
[14]许大全. 光合作用气孔限制分析中的一些问题[J]. 植物生理学通讯,1997,33(04):241-244.
[15]Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis[J]. Annu. Rev. Plant Physiol., 1982, 33: 317-345.
[16]周小玲, 田大伦, 许忠坤, 等. 中亚热带四川桤木与台湾桤木幼林的光合生态特性[J]. 中南林业科技大学学报, 2007,27(1): 40-49, 54.
[17]葛 滢, 常 杰, 陈增鸿, 等. 青冈净光合作用与环境因子的关系[J]. 生态学报, 1999, 19(5): 683-688.
[本文编校:谢荣秀]
Research on diurnal variation of photosynthetic characteristics ofCornus wilsonianaclones during friuting season
WU Linying1,2, DENG Wenjian3, ZHANG Yingzhong1,2, LIN Jun3, DENG Zhixi3, ZHANG Liangbo4,5, HE Zhenxiang6, WANG Minghuai1,2
(1. Guangdong Provincial Key Laboratory of Silviculture, Protection and Utilization, Guangzhou 510520, Guangdong, China;2.Guangdong Academy of Forestry, Guangzhou 510520, Guangdong, China; 3. Lechang Longshan Forest Farm, Lechang 512221,Guangdong, China; 4.Hunan Academy of Forestry, Changsha 410004, Hunan, China; 5.Hunan Engineering Research Center of Biodiesel, Changsha 410004, Hunan, China; 6.School of Life Sciences, Nanjing University, Nanjing 210093, Jiangsu, China)
In order to explore the diurnal variation of photosynthetic characteristics ofCornus wilsonianaclones, the diurnal variation of photosynthesis of 8Cornus wilsonianaclones were measured in friuting season and analyzed using an Li-6400 portable photosynthetic analyzer. The results showed as follows: (1) The diurnal variation of net photosynthetic rate (Pn) and stomatal conductance(Gs) of all tested clones showed single-peak curve, and the peak of most clones is at 9:00 am. The diurnal variation of transpiration rate (Tr)revealed double-peak curve, except JX13 and GY8040, and thefirst peak is higher than the second. The diurnal variation of intercellular concentration (Ci) appeared tabular W-type. The diurnal variation of water use efficiency (WUE) showed V-shape curve, and the minimum values is at about 15:00 pm. (2) The daily average values ofPn,Tr,Gs,Ci, WUE of 8Cornus wilsonianaClones had not significant difference, respectively. ThePnof GY8040, JX3, LC7, JX6 are higher than others, over 10.0 μmol·m-2s-1. (3) In the effect of environment factors, PAR directly influence the photosynthesis physiological characteristics or indirectly byTaand RH,Cashowed remarkable positive correlation withCi.Pnhad very significant positive correlation withGsandTr, The importance of factors effect onPnwasGs>Tr> PAR > RH >Ta> WUE >Ci>Ca.
Cornus wilsoniana; clone; diurnal variation of photosynthetic characteristics; environment factor; correlation
S718.43
A
1673-923X(2017)09-0032-06
10.14067/j.cnki.1673-923x.2017.09.006
2016-10-17
广东省省级科技计划项目“油料植物光皮树良种资源收集与无性系选育”(2014A020208033)
吴林瑛,助理工程师,硕士
王明怀,研究员,硕士;E-mail:1073526066@qq.com
吴林瑛,邓文剑,张应中,等. 光皮树不同无性系果期光合日变化研究[J].中南林业科技大学学报, 2017, 37(9): 32-37, 56.
