共生菌Arsenophonus、水稻品种和温度对褐飞虱黄绿绿僵菌发病率的影响
2017-12-02朱欢欢陈洋万品俊王渭霞赖凤香傅强
朱欢欢 陈洋 万品俊 王渭霞 赖凤香 傅强
(中国水稻研究所 水稻生物学国家重点实验室,杭州 310006;*通讯联系人,E-mail: fuqiang@caas.cn)
共生菌Arsenophonus、水稻品种和温度对褐飞虱黄绿绿僵菌发病率的影响
朱欢欢 陈洋 万品俊 王渭霞 赖凤香 傅强*
(中国水稻研究所 水稻生物学国家重点实验室,杭州 310006;*通讯联系人,E-mail: fuqiang@caas.cn)
【目的】研究共生菌 Arsenophonus、水稻和温度对褐飞虱黄绿绿僵菌发病率的影响,揭示影响褐飞虱黄绿绿僵菌发病的重要因子。【方法】通过共生菌Arsenophonus(感染和未感染)与水稻品种(TN1、IR56和Mudgo)以及 Arsenophonus与温度(21℃、23℃、25℃、27℃、29℃和 31℃)两个二因素完全随机区组试验,观察喷黄绿绿僵菌孢子悬浮液后不同时间褐飞虱黄绿绿僵菌的发病率。【结果】喷黄绿绿僵菌孢子悬浮液后,不同水稻品种上褐飞虱发病率均以含Arsenophonus的试虫较低,而不同温度下的褐飞虱发病率则仅喷菌处理后第3 d和5 d时在25℃、27℃和29℃时含Arsenophonus的试虫发病率显著较低,其他温度下无显著差异。双因素方差结果表明,Arsenophonus与水稻品种双因素试验中均以含 Arsenophonus褐飞虱黄绿绿僵菌的发病率显著较低,而Arsenophonus与温度双因素试验中仅在喷菌处理后第3 d、5 d以含Arsenophonus褐飞虱黄绿绿僵菌的发病率显著较低。温度显著影响褐飞虱的发病率,23℃~29℃条件下褐飞虱的发病率较高,LT50较短,为黄绿绿僵菌侵染褐飞虱的适温范围。其中,27℃时发病率最高,LT50最短,最为适宜;31℃和21℃条件下褐飞虱发病率较低,LT50较长,不利于黄绿绿僵菌对褐飞虱的侵染。温度还影响含Arsenophonus与不含Arsenophonus试虫LT50的相对大小。其中,含Arsenophonus试虫LT50在27℃、29℃时相对较长,在其他温度下则相同甚至较短。【结论】共生菌Arsenophonus显著抑制黄绿绿僵菌对褐飞虱的致病力,且该作用受到温度的显著影响;温度亦显著影响褐飞虱的黄绿绿僵菌发病率,而水稻品种以及Arsenophonus和水稻品种间、Arsenophonus和环境温度间的交互作用均无显著影响。
褐飞虱;黄绿绿僵菌;Arsenophonus;温度;水稻品种
褐飞虱[Nilaparvata lugens (Stål)]属于半翅目飞虱科(Hemiptera: Delphacidae),是我国及南亚、东南亚国家水稻上的主要害虫[1];主要通过口针吸食水稻茎秆韧皮部的液汁为害,消耗稻株水分和养分,造成谷粒灌浆不饱满,千粒重下降,严重时整株枯死,田间形成“虱烧”;还可以传播水稻草状矮缩病(Rice grassy stunt virus,RGSV)和齿叶矮缩病(Rice ragged stunt virus,RRSV)等病毒,间接对稻株造成危害[2-4]。据统计,我国自20世纪80年代以来,褐飞虱的发生面积约占水稻种植面积的50%,年均损失稻谷仍达10亿~15亿kg[1],亚洲范围内每年由褐飞虱引起的水稻产量的损失高达3亿美元[5]。
目前,化学杀虫剂是防治褐飞虱的主要手段。然而,由于化学杀虫剂的长期、大量、不合理使用,导致褐飞虱对吡虫啉、噻嗪酮等生产上常用杀虫剂产生了高抗药性[5-7],进一步加剧农药的过量使用。利用病原微生物是减少褐飞虱防控对化学农药依赖程度的重要手段之一。
黄绿绿僵菌(Metarhizium flavoviride Gams amp;Rozsypal)是一类重要的稻飞虱病原真菌[8-10]。环境温度及寄主昆虫褐飞虱自身生理状况是影响绿僵菌致病力的重要因素,但有关的研究不多。关于环境温度的影响主要为温度对绿僵菌菌株生长、孢子萌发的影响。常金梅等[11]研究发现,绿僵菌MC3在23℃~26℃时,随着温度的升高,生长速度显著加快;岳梅等[12]报道绿僵菌LA菌株在前12 h孢子萌发速率从高到低为30℃>35℃>25℃>20℃>40℃,24 h内20℃~35℃下孢子萌发率达到90%以上,而40℃下无萌发现象;李佳颖等[13]研究发现黄绿绿僵菌SM076在17℃~32℃温度范围内均可生长和产孢,26℃下菌落生长速率最快,孢子萌发率最高,而23℃左右则最适产孢。关于绿僵菌对稻飞虱的侵染致病温度条件的研究较少。杜光祖等[14]指出,24℃~30℃下,黄绿绿僵菌对灰飞虱3龄若虫均有致病力,其中,27℃和30℃下的致病力较强。
褐飞虱自身生理状况的影响则主要集中在褐飞虱的龄期及所含共生菌的研究。耿博闻等[15]指出黄绿绿僵菌对不同发育阶段褐飞虱的毒力不同,其毒力顺序为成虫期>高龄若虫期>低龄若虫期,对成虫的LT50为5.66 d;李茂业等[16]报道,黄绿绿僵菌Mf82对褐飞虱除2龄若虫以外的其余各个虫态的累积死亡率均大于50%,其中褐飞虱成虫的致死率可达85.7%。类似地,杜光祖等[14]发现黄绿绿僵菌KM1104对灰飞虱成虫的致病力高于3龄若虫。陈宇等[17]则报道含有Arsenophonus类共生细菌的褐飞虱对黄绿绿僵菌的抵抗能力显著提高。
Arsenophonus是褐飞虱体内的一类非专性内生共生细菌,属于变形菌门(Proteobacteria)γ亚类肠杆菌科(Enterobacteriaceae),由母体垂直传递给子代[18],其提高褐飞虱对黄绿绿僵菌抵抗力的作用是否受到环境温度的影响尚不清楚。此外,采用水稻抗性品种是防治褐飞虱的经济有效措施[19],不利于褐飞虱的生长发育或繁殖,是否会进而使Arsenophonus对褐飞虱黄绿绿僵菌致病力的影响发生变化,这亦是一个值得关注的重要问题。为此,本研究拟通过两个双因素实验开展两方面的研究:1)共生菌Arsenophonus与水稻品种(TN1、IR56和Mudgo)对黄绿绿僵菌致病褐飞虱的影响;2)Arsenophonus和温度(21℃、23℃、25℃、27℃、29℃和31℃)对绿僵菌侵染褐飞虱的影响。
1 材料与方法
1.1供试材料
供试昆虫:褐飞虱采自浙江富阳田间,参照王渭霞等[18]的方法筛选出含Arsenophonus和不含Arsenophonus的两个褐飞虱种群,用TN1稻苗室内饲养。羽化24 h内的雌成虫用于本研究。
供试菌株:采自浙江富阳稻田的褐飞虱罹病虫体。参照吴发红等[21]的方法进行分离及鉴定,确定为黄绿绿僵菌ARSEF 1764。其孢子悬浮液的配制步骤如下:将黄绿绿僵菌接(平行划线)到PDA培养基平板上,在(25±1)℃下黑暗培养10 d。待菌株充分产孢后,用灭菌过的接种环轻轻将该菌株的分生孢子粉刮到盛有10 mL 0.05% Tween-20湿润剂的三角瓶中,在涡旋混合器上充分振荡30 min,无菌纱布过滤菌液,获得孢子悬浮液母液。用血球计数器计数每mL母液的分生孢子含量,然后配制成1.0×108个/mL的孢子悬浮液,备用。
1.2共生菌Arsenophonus和水稻品种对褐飞虱黄绿绿僵菌发病率的影响
取含Arsenophonus和不含Arsenophonus共生菌的褐飞虱初羽化雌成虫,喷5 mL的黄绿绿僵菌孢子悬浮液后分别转移至TN1、Mudgo和IR56稻苗上笼罩饲养,以喷5 mL 0.05% Tween-20为对照,共12个处理,每个处理重复5次。每个重复接虫40头。黄绿绿僵菌孢子悬浮液或Tween-20均匀喷至稻苗与试虫体表。处理后置于温度(25±1)℃、相对湿度(85±5)%和光周期14 L/10 D的培养箱中饲养。
处理后第1天去除死虫,将剩余活虫作为基数从喷菌后第3天开始观察褐飞虱的死亡情况,隔日观察至处理后第9天。每次检查后移出死虫,转移至培养皿中继续保湿培养,3 d后记录各重复的发病虫数(包括体表长出黄绿绿僵菌菌丝的僵虫及经PCR检测阳性的未长出菌丝的死虫),计算各处理的发病率。若未喷绿僵菌的试虫出现病虫,则并以同次观察的同一品种上同类试虫的Tween-20对照为参照计算校正发病率。其中死虫黄绿绿僵菌的PCR检测参照吴发红等[21]的方法;校正发病率按校正发病率(%)= (处理组发病率-对照组发病率)/(1-对照组发病率)×100计算。
1.3共生菌Arsenophonus和不同温度对褐飞虱黄绿绿僵菌发病率的影响
取含Arsenophonus或不含Arsenophonus的褐飞虱初羽化雌成虫,经黄绿绿僵菌喷菌处理后接到TN1稻苗上,以0.05% Tween-20为对照。试验在不同温度(21℃、23℃、25℃、27℃、29℃和31℃)的培养箱(相对湿度 85%±5%、光周期14 L/10 D)中进行。每个处理重复3次,每个重复接虫40头。
处理后第2 d开始,每日观察褐飞虱的死亡情况直至试虫全部死亡,并按“1.2”的方法观察病虫,计算发病率和校正发病率。
小孙啊,你不要紧张,我找你来随便聊聊。前天下午,我收到你夫人赵颖春给我写的一封信……周书记说到这里故意停顿了一下,而我的头已经大了,心里骂道,这个该死的臭婆娘,嘴里却赶紧说,周书记,对不起,颖春她不懂事……
1.4统计分析
利用DPS数据处理软件(V14.10)[22]进行统计分析。处理后各个时间同一品种或同一温度下含Arsenophonus与不含Arsenophonus试虫发病率间的单因素比较采用两样本率比较法进行。Arsenophonus与水稻品种或环境温度的双因素方差分析则采用二因素完全随机区组模块,百分数数据进行分析前反正弦平方根转换;各因素内不同处理间采用邓肯氏新复极差法进行多重比较。不同温度处理下的致死中时间(LT50)计算则采用死亡率-时间机率值分析模块进行。
2 结果与分析
2.1共生菌Arsenophonus和水稻品种对褐飞虱黄绿绿僵菌发病率的影响
黄绿绿僵菌处理后,各水稻品种上含Arsenophonus与不含Arsenophonus褐飞虱的发病率均以含Arsenophonus的试虫为低,无一例外,且除TN1上处理9 d后未达显著水平,TN1上处理后3 d和Mudgo上处理后7 d仅达0.10显著水平外,其余均达到了显著水平(P<0.05)或极显著水平(P<0.01)(图1)。
不喷菌试虫的发病率均为零,我们以喷菌试虫的发病率进行共生菌Arsenophonus与水稻品种的双因素方差分析,结果表明,处理后3~9 d,Arsenophonus对褐飞虱发病率有显著影响(P<0.05),且均以含Arsenophonus的褐飞虱发病率显著低于不含Arsenophonus的褐飞虱(图2);而水稻品种及其与共生菌的交互作用对发病率均无显著影响(P>0.05)。显然,Arsenophonus降低了褐飞虱对黄绿绿僵菌的感染率,而水稻品种的抗性及其与Arsenophonus的交互作用对褐飞虱绿僵菌感染率无显著影响。
从图2还可以看出黄绿绿僵菌处理后褐飞虱发病率的时间动态,处理后第3天发病率较低,之后迅速上升,至第7天趋于稳定。
2.2共生菌Arsenophonus和温度对褐飞虱黄绿绿僵菌发病率的影响结果
黄绿绿僵菌处理后,6个温度条件下,含Arsenophonus与不含Arsenophonus褐飞虱发病率的差异在21℃、23℃、31℃下各观察时间均不显著(P>0.05);而在25℃、27℃和29℃下,仅喷菌处理后3 d和5 d时差异显著(P<0.05)或极显著(P<0.01),且均以含Arsenophonus的试虫发病率较低,但处理后7 d、第9 d这种差异缩小,未达到显著水平(P>0.05)(图3),表明Arsenophonus对褐飞虱黄绿绿僵菌发病率有抑制作用,且该作用受到温度的显著影响。
不喷黄绿绿僵菌时,有超过半数的对照(56.3%)出现0.8%~4.9%的发病率,因此以校正发病率为指标进行Arsenophonus共生菌与温度的双因素方差分析,结果表明,Arsenophonus对褐飞虱校正发病率的影响仅在处理后3 d、5 d显著(P<0.05),其中,以含Arsenophonus试虫的校正发病率较低;而处理后7 d、9 d不显著(P>0.05)。温度对褐飞虱校正发病率在处理后4个时间均有显著影响(Plt;0.05),温度显著影响褐飞虱的绿僵菌发病率。其中,处理后3 d,21℃~29℃的校正发病率无显著差异,且高于31℃,但只有25℃、27℃与31℃差异显著;处理后5 d,27℃的校正发病率显著高于其他5个温度,而其他5个温度间无显著差异;处理后7 d、9 d,25℃、29℃的校正发病率与27℃无显著差异,23℃则显著高于21℃和31℃,但显著低于27℃,其中处理后第7 d还显著低于25℃和29℃(图4)。总体上,6个温度下的校正发病率由高到低依次为27℃、25℃、29℃、23℃、21℃和31℃。
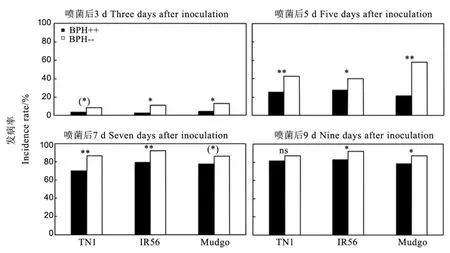
图1 黄绿绿僵菌处理后三个水稻品种上含Arsenophonus与不含Arsenophonus褐飞虱发病率的比较Fig. 1. Incidence rate of Metarhizium flavoviride-infected brown planthoppers with and without Arsenophonus reared on three rice varieties.
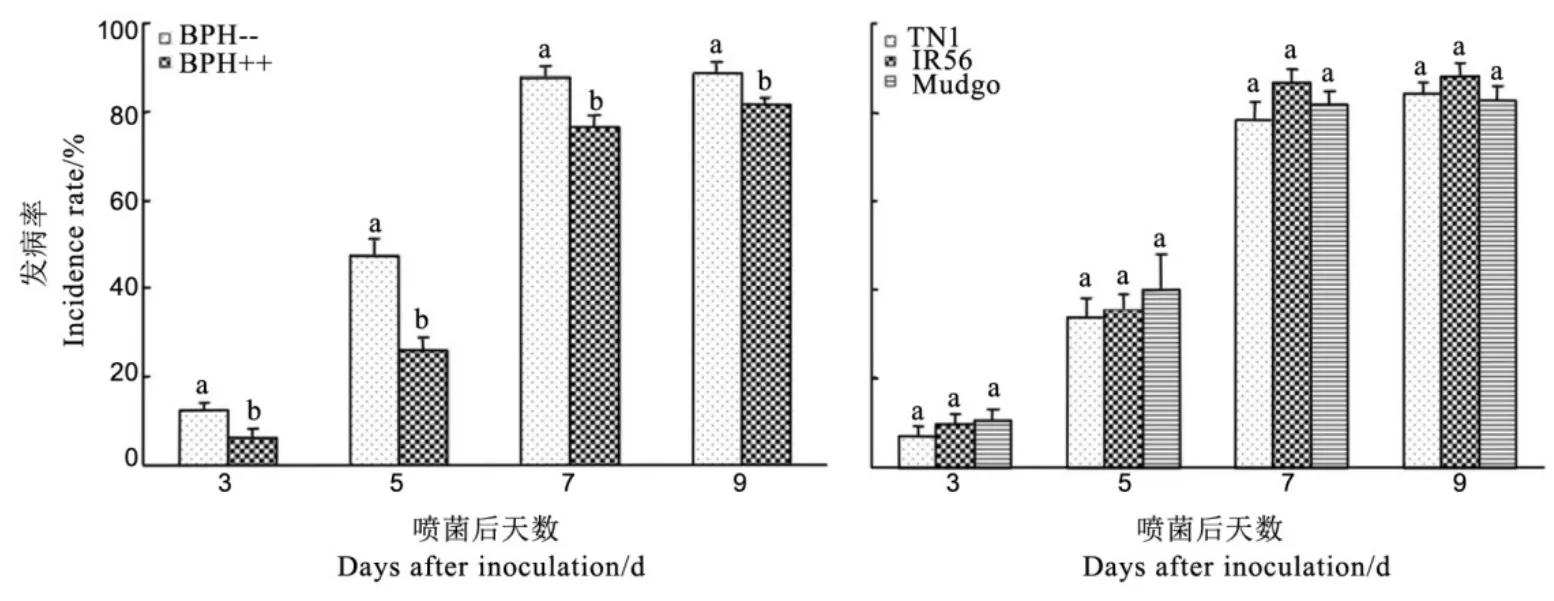
图2 共生菌Arsenophonus(左)和水稻品种(右)对褐飞虱黄绿绿僵菌发病率的影响(平均值±标准误)Fig. 2. Effects of Arsenophonus (left) and rice varieties (right) on the incidence rate of Metarhizium flavoviride-infected brown planthoppers ( Mean ± SE ).
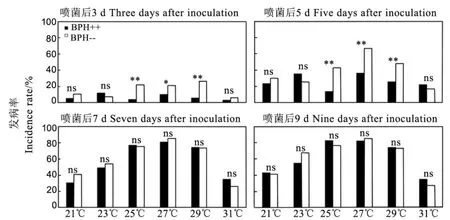
图3 黄绿绿僵菌处理后在不同温度条件下含Arsenophonus与不含Arsenophonus褐飞虱发病率的比较Fig. 3. Incidence rate of Metarhizium flavoviride-infected brown planthoppers with and without Arsenophonus under different temperatures,respectively.
进一步分析不同温度条件下褐飞虱试虫的致死中时间(LT50)(表1)发现,23℃~29℃条件下LT50较短(4.7~7.6 d),且27℃时最短(4.7~5.3 d);21℃、31℃时均较长,其中,不含Arsenophonus的试虫超过10.2 d,含Arsenophonus的试虫在8.8 d以上。结合前文不同温度下试虫校正发病率的排序结果,可以认为23℃~29℃是黄绿绿僵菌侵染褐飞虱的适宜温度范围,且以27℃最为适宜,温度过高(31℃)或过低(21℃)均不利于黄绿绿僵菌对褐飞虱的侵染。
含Arsenophonus与不含Arsenophonus试虫的LT50相比,27℃、29℃时均以含Arsenophonus试虫较长,但25℃时两类试虫相同,且21℃、23℃、31℃三个温度下均以含Arsenophonus试虫的LT50较短,进一步说明环境温度影响Arsenophonus共生菌对褐飞虱黄绿绿僵菌发病率的抑制作用。
3 讨论
绿僵菌(Metarhizium. Sp)是一类重要的生防真菌,全世界大约有200多种昆虫、线虫及螨类可以被其感染致死[23-25],是全世界研究应用最多的虫生真菌之一。黄绿绿僵菌作为一种潜在的稻飞虱生防菌倍受关注[14-16,26],但迄今尚未在生产上大面积应用。探明黄绿绿僵菌侵染稻飞虱的重要影响因子是推进其应用的重要基础。绿僵菌对昆虫的侵染,除取决于绿僵菌菌株自身因素(如菌株的生化特性[27]、菌株继代次数[28])之外,寄主昆虫体内的共生菌[17,29]和环境温度[12,30-31]是重要因素。
本研究进一步证实感染Arsenophonus可降低褐飞虱黄绿绿僵菌的发病率。类似的现象亦见于其他昆虫的相关研究报道。豌豆蚜体内的Rickettsia insecticola降低了真菌的传播速度,有效地抵抗真菌的侵染[29]。欧洲狼蜂(Philanthus Triangulum)可以利用触角中的Streptomyces保护幼虫免受致病真菌侵害[32]。木虱Psyllid体内的Arsenophonus可以抵抗拟寄生物Psyllaphaegus bliteus的寄生[33]。目前,共生菌影响昆虫抵御病原菌或其他天敌的机制尚不清楚,有学者推测共生菌通过诱导宿主昆虫免疫反应来提高宿主抗性[32]。笔者亦观察到,黄绿绿僵菌侵染褐飞虱时,含Arsenophonus的褐飞虱雌成虫的浆血胞数量显著高于不含Arsenophonus的褐飞虱(待发表资料),似表明Arsenophonus共生菌通过血细胞免疫系统而影响褐飞虱黄绿绿僵菌感病性,但尚需进一步探明。
水稻品种抗性是控制稻飞虱的第一道屏障,含Bph1抗虫基因的水稻品种曾在东南亚及我国水稻生产中得到大面积应用,近年来含Bph3抗虫基因的水稻品种的选育亦在我国得到重视[1]。本研究明确水稻品种[TN1、IR56(含抗虫基因Bph3)和Mudgo(含抗虫基因Bph1)]对褐飞虱黄绿绿僵菌的发病率无显著影响,且与褐飞虱Arsenophonus共生菌无交互作用,这可能表明水稻抗性品种尽管不利于褐飞虱的生长发育和繁殖,但并未影响褐飞虱对黄绿绿僵菌的敏感性。不过,本研究中的水稻品种有限,该现象是否适合于其他抗性水稻品种尚不清楚。
温度是影响黄绿绿僵菌对褐飞虱的侵染的重要因素,这与前人[14,30]的结果类似。温度对绿僵菌的作用体现在对产孢能力和致病力两个方面的影响[11,12,31]。绿僵菌孢子侵染寄主的过程中需产生蛋白酶、几丁质酶和脂肪酶来溶解寄主表皮以完成侵染,而绿僵菌的蛋白酶活性是其毒力的决定因素之一[34-36],温度通过对酶活的影响进而影响绿僵菌孢子的侵染和毒力。对东亚飞蝗的研究表明,在极限高温和极限低温时,绿僵菌LA菌株的胞外蛋白酶产量明显减少,其对飞蝗5龄若虫的毒力降低;在最适生长温度时,LA菌株的毒力也较强[12]。在温度的具体影响方面,本研究发现23℃~29℃是黄绿绿僵菌ARSEF1764致病褐飞虱的适温范围,27℃为最适宜温度,温度为31℃或21℃时绿僵菌致病力显著降低,这与李佳颖等[13]的黄绿绿僵菌SM076在26℃下菌落生长速度最快、孢子萌发率最高以及杜光祖等[14]黄绿绿僵菌对灰飞虱3龄若虫在27℃下致病力较强的结果基本一致;但与李佳颖等[13]报道的23℃最适黄绿绿僵菌SM076产孢的结果相差较大,推测黄绿绿僵菌产孢与菌落生长、孢子萌发的适宜温度有所不同。此外,杜光祖等[14]还发现黄绿绿僵菌在30℃下对灰飞虱3龄若虫亦有较强致病力;岳梅等[12]则报道绿僵菌LA菌株在30℃、35℃孢子萌发速率最快,结果与本文有所不同,究其原因可能有:1)绿僵菌菌株不同,不同菌株对温度的反应可能存在一定的差异;2)宿主昆虫不同,也可能造成温度反应的差异。
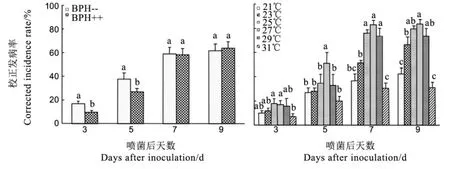
图4 共生菌Arsenophonus (左)和温度(右)对褐飞虱黄绿绿僵菌校正发病率的影响(平均值±标准误)Fig. 4. Effects of Arsenophonus (left) and temperatures (right) on the corrected incidence rate of Metarhizium flavoviride-infected brown planthoppers(Mean ± SE).

表1 不同温度下黄绿绿僵菌处理后褐飞虱的致死中时间LT50Table 1. Median lethal time (LT50) of the brown planthoppers after Metarhizium flavoviride infestation at different temperatures. d
本研究中,尽管褐飞虱含菌状况与温度对黄绿绿僵菌发病率无显著的交互作用,但温度可能影响Arsenophonus对褐飞虱绿僵菌发病率的抑制作用,表现在两方面:1)不同温度条件下含Arsenophonus与不含Arsenophonus试虫的发病率仅在25℃、27℃、29℃时有显著差异,而21℃、23℃、31℃时各观察时间均无显著差异;2)不同温度下含Arsenophonus与不含Arsenophonus试虫的致死中时间(LT50)仅27℃、29℃时以含Arsenophonus的试虫相对较长,与Arsenophonus抑制褐飞虱黄绿绿僵菌发病的结果不一致,其他温度下含Arsenophonus的试虫则相同甚至较短,与Arsenophonus抑制褐飞虱黄绿绿僵菌发病的结果一致。推测Arsenophonus对黄绿绿僵菌侵染褐飞虱的抑制作用可能局限在一定的温度范围之内。因Arsenophonus抑制黄绿绿僵菌侵染褐飞虱的机制还不清楚,这种温度影响机制亦需进一步研究。
本文Arsenophonus与水稻品种、Arsenophonus与温度两个双因素实验的方差分析中,处理后7 d、9 d时Arsenophonus对褐飞虱黄绿绿僵菌发病率的影响并不一致,即前者影响显著(图2,左)。而后者不显著(图4,左)。这与前者体现不同水稻品种的综合影响而后者体现不同温度的综合影响有关。如前所述,不同温度下Arsenophonus对褐飞虱黄绿绿僵菌发病率的抑制作用有所不同,而水稻品种则无明显影响,在处理后7 d、9 d含Arsenophonus与不含Arsenophonus试虫发病率的差值较处理后5 d时有所收窄的前提下,可能导致Arsenophonus与温度双因素实验中Arsenophonus的影响达不到显著水平。
[1] 郭予元. 中国农作物病虫害.3版:上册. 北京: 中国农业出版社,2015:93-104.Guo Y Y. Crop Diseases and Insect Pests in China. 3rd ed:Vol. I. Beijing: China Agriculture Press,2015: 93-104. (in Chinese)
[2] Douglas A E. Nutritional interactions in insect-microbial symbioses. Annu Rev Entomol,1998,43: 17-37.
[3] Zchori-Fein E,Brown J K. Diversity of prokaryotes associated with Bemisia tabaci (Gennadius) (Hemiptera:Aleyrodidae). Ann Entomol Soc Am,2002,95(6):711-718.
[4] Werren J H. Arsenophonus // Garrity T M. Bergey's Mannual of Systematic Bacteriology. New York: Springer Verlag,2005.
[5] Min S,Lee S W,Choi B R,Lee S H,Kwon D H.Insecticide resistance monitoring and correlation analysis to select appropriate insecticides against Nilaparvata lugens (Stål),a migratory pest in Korea. J Asia-Pacific Entomol,2014,17 (4): 711-716.
[6] 王彦华,王强,沈晋良,吴声敢,俞瑞鲜,赵学平,苍涛,吴长兴,陈丽萍. 褐飞虱抗药性研究现状. 昆虫知识,2009,46 (4): 518-524.Wang Y H,Wang Q,Shen J L,Wu S G,Yu R X,Zhao X P,Cang T,Wu C X,Chen L P. Current status of insecticide resistance in the brown planthopper,Nilaparvata lugens. Entomol Knowl,2009,46 (4):518-524. (in Chinese with English abstract)
[7] Zhang X L,Liao X,Mao K K,Zhang K X,Wan H,Li J H.Insecticide resistance monitoring and correlation analysis of insecticides in field populations of the brown planthopper Nilaparvata lugens (stål) in China 2012-2014. Pestic Biochem Physiol,2016,132: 13-20.
[8] 耿博闻. 黄绿绿僵菌防治稻飞虱的研究. 广州: 中山大学,2004.Geng B W. Studies on biological control of rice planthopper with Metarrhizium anisopliae var. acridum.Guangzhou: Sun Yat-sen University,2004. (in Chinese with English abstract)
[9] 徐阿妹. 黄绿绿僵菌培养生产技术及其在害虫防治中的应用. 合肥: 安徽农业大学,2013.Xu A M. Culture production technology of Metarhizium flavoviride and its application in pest control. Hefei:Anhui Agricultural University,2013. (in Chinese with English abstract)
[10] Li M Y,Lin H F,Li S G,Xu A M,Feng M F. Efficiency of entomopathogenic fungi in the control of eggs of the brown planthopper Nilaparvata lugens Stål (Homopera:Delphacidae). Afr J Microbiol Res,2012,6(44):7162-7167.
[11] 常金梅,何衍彪,赵燕龙,柳凤,詹儒林. 绿僵菌致病力的制约因素研究. 安徽农业科学,2010,38(6):2815-2816.Chang J M,He Y B,Zhao Y L,Liu F,Zhan R L. Study on the factors restraining the pathogeulcity of Metarhizium sp. J Anhui Agric Sci,2010,38(6): 2815-2816. (in Chinese with English abstract)
[12] 岳梅,雷仲仁,孟涛,王兆勇,姚君明. 温度对绿僵菌LA菌株孢子萌发、酶活力及毒力的影响. 植物保护,2010,36(2): 56-60.Yue M,Lei Z R,Meng T,Wang Z Y,Yao J M.Temperature influence on conidial germination rate,enzyme activity and virulence of Metarhizium anisopliae LA isolate. Plant Prot,2010,36(2): 56-60. (in Chinese with English abstract)
[13] 李佳颖,许齐爱,代晓彦,汪郭凰,任顺祥. 黄绿绿僵菌 SM076菌株生物学特性研究. 广东农业科学,2013(9): 71-73.Li J Y,Xu Q A,Dai X Y,Wang G H,Ren S X. Study on the biological characteristics of Metarhizium flavoviride.J. Guangdong Agric Sci,2013(9): 71-73. (in Chinese with English abstract)
[14] 杜光祖,杨卫秋,周跃能,和淑琪,陈斌,桂富荣,李正跃. 黄绿绿僵菌 KM1104对灰飞虱的毒力. 植物保护,2015,41(5): 207-211.Du G Z,Yang W Q,Zhou Y N,He S Q,Chen B,Gui F R,Li Z Y. Virulence of Metarhizium flavoviride strain KM1104 against Laodelphax striatellus (Fallén). Plant Prot,2015,41(5): 207-211. (in Chinese with English abstract)
[15] 耿博闻,张润杰. 低浓度唾嗓酮与黄绿绿僵菌对褐飞虱协同作用的生物测定. 植物保护学报,2005,32(1):53-56.Geng W B,Zhang R J. Interactive effect of low concentration buprofezin Metarhizium anisopliae var.acridum on the mortalities of Nilaparvata legens(Hemiptera: Delphacidae) nymphs and adults in laboratory bioassays. Acta Phytophy Sin,2005,32(1):53-56. (in Chinese with English abstract)
[16] 李茂业,林华峰,李世广,金立. 黄绿绿僵菌 Mf82菌株对不同虫态褐飞虱的毒力. 昆虫学报,2012,55(3):316-323.Li M Y,Lin H F,Li S G,Jin L. Virulence of Metarhizium flavoviride 82 to different developmental stages of the brown planthopper,Nilaparvata legens (Hemiptera:Delphacidae). Acta Entomol Sin,2012,55(3): 316-323.(in Chinese with English abstract)
[17] 陈宇,陈洋,王渭霞,赖凤香,傅强. 褐飞虱杀雄菌属共生菌 Arsenophonus的传递方式与生物学意义初探.中国水稻科学,2014,28(1): 92-96.Chen Y,Chen Y,Wang W X,Lai F X,Fu Q. A preliminary study on the transfer mode and biological significance of endosymbiont Arsenophonus in Nilaparvata lugens. Chin J Rice Sci,2014,28(1): 92-96.(in Chinese with English abstract)
[18] 王渭霞,罗举,赖凤香,傅强. 水稻褐飞虱内生共生细菌 Arsenophonus的鉴定和系统分析. 昆虫学报,2010,53(6): 647-654.Wang W X,Luo J,Lai F X,Fu Q. Identification and phylogenetic analysis of symbiotic bacteria Arsenophonus from the rice brown planthopper,Nilaparvata lugens (Stål) (Hemiptera: Delphacidae). Acta Entomol Sin,2010,53(6): 647-654. (in Chinese with English abstract)
[19] Wang Y,Cao L M,Zhang Y X,Cao C X,Liu F,Huang F K,Qiu Y F,Li Y B,Lou X J. Map-based cloning and characterization of BPH29,a B3domain-containing recessive gene conferring brown planthopper resistance in rice. J Exp Bot,2015,66(19): 6035-6045.
[20] 方中达. 植病研究方法. 北京:中国农业出版社,1998:40-72.Fang Z D. Methodology for plant pathology. Beijing:China Agriculture Press,1998: 40-72. (in Chinese)
[21] 吴发红,黄东益,黄小龙,周鑫,程文杰. 几种真菌DNA提取方法的比较. 中国农学通报,2009,25(8):62-64.Wu F H,Huang D Y,Huang X L,Zhou X,Cheng W J.Comparing study on several methods for DNA extraction from endophytic fungi. Chin Agric Sci Bull,2009,25(8):62-64. ( in Chinese with English abstract )
[22] 唐启义. DPS数据处理系统. 3版. 北京: 科学出版社,2013.Tang Q Y. Applied Statistic Analysis and Date Processing System. 3ed. Beijing: Science Press,2013. (in Chinese)
[23] 蒲蛰龙,李增智. 昆虫真菌学. 合肥: 安徽科学技术出版社,1996.Pu Z L,Li Z Z. Insect Mycology. Hefei: Anhui Science and Technology Press,1996. (in Chinese)
[24] 刘爱英. 中国虫生真菌研究与应用. 北京: 中国农业科技出版社,1991: 159-164.Liu A Y. Study and application of entomogenous fungi in China. Beijing:China Agriculture Science and Technology Press,1991: 159-164.(in Chinese)
[25] Mnyone L L,Kirby M J,Lwetoijera D W,Mpingwa M W,Simfukwe E T,Knols B G,Takken W,Russell T L. Tools for delivering entomopathogenic fungi to malaria mosquitoes: Effects of delivery surfaces on fungal efficacy and persistence. Malar J,2010,9: 246.
[26] Jin S F,Feng M G,Chen J Q. Selection of global Metarhizium isolates for the control of the rice pest Nilaparvata lugens (Homoptera: Delphacidae). Pest Manag Sci,2008,64(10): 1008-1014.
[27] 樊美珍,郭超,王姝清,谢飙.不同菌株绿僵菌酯酶型与其致病力的关系. 西北林学院学报,1990,5(3): 1-6.Fan M Z,Guo C,Wang S Q,Xie B. Relathionship Between esterase type and pathogenic virulence of different strains of Metarhizium spp. J Northwest For Coll,1990,5(3): 1-6. (in Chinese with English abstract)
[28] 孙召朋. 继代培养对球孢白僵菌毒力及毒力相关酶表达的影响. 长春: 吉林大学,2011.Sun S P. Influence on the virulence of Beauveria bassiana and the expression of virulence related enzymes during subculturing. Changchun: Jilin University,2011.(in Chinese with English abstract)
[29] Scarborough C L,Ferrari J,Godfray H C J. Aphid protected from pathogen by endosymbiont. Science,2005,310(5755): 1781-1781.
[30] 汪敏捷,刘强. 不同温湿度下绿僵菌对红缘天牛幼虫致病力的影响. 环境昆虫学报,2014,36(2): 151-156.Wang M J,Liu Q. Pathogenicity of the Metarhizium anisopliae to Asias halodendri larvae at different temperature and humidity conditions. J Eniron Entomol,2014,36(2): 151-156. (in Chinese with English abstract)
[31] 朱彬洲,雷仲仁,徐洪富,王兆勇,梁兴慧. 不同温湿度条件下绿僵菌 LA菌株对东亚飞蝗的致病力. 中国生物防治,2010,26(4): 448-452.Zhu B Z,Lei Z R,Xu H F,Wang Z Y,Liang X H.Virulence of Metarhizium anisopliae against Locusta migratoria manilensis under different temperatures and humidities. Chin J Biol Contr,2010,26(4): 448-452. (in Chinese with English abstract)
[32] 徐红星,郑许松,刘淑平,叶恭银,吕仲贤. 昆虫内共生菌在昆虫防御中的作用. 昆虫知识,2009,46(3):350-354.Xu H X,Zheng X S,Liu S P,Ye G Y,Lu Z X. The role of endosymbionts in insect host resistance against adverse factors. Chin Bull Entomol,2009,46(3): 350-354. (in Chinese with English abstract)
[33] Hansen A K,Jeong G,Paine T D,Stouthamer R.Frequency of secondary symbiont infection in an invasive psyllid relates to parasitism pressure on a geographic scale in California. App Environ Microb,2007,73(23):7531-7535.
[34] 冯明光. 胞外蛋白酶和脂酶活性作为球孢白僵菌毒力指标的可靠性分析. 微生物学报,1998(6): 461-467.Feng M G. Reliability of extracellular protease and lipase activities of Beauveria bassiana isolates used as their virulence indices. Acta Microbiol Sin,1998(6): 461-467.(in Chinese with English abstract)
[35] 林海萍,魏锦瑜,毛胜凤,陈喜,张立钦,马良进. 球孢白僵菌蛋白酶、几丁质酶、脂肪酶活性与其毒力相关性. 中国生物防治,2008,24(3): 290-292.Lin H P,Wei J Y,Mao S F,Chen X,Zhang L Q,Ma L J.Correlation between protease,chitinase and lipase activities and virulence of Beauveria bassiana against Monochamus alternatus. Chin J Biol Contr,2008,24(3):290-292. (in Chinese with English abstract)
[36] St Leger R J,Dumnds P K,Chamley A K,Cooper R M.Role of extracellular chymoelastase in the virulence of Metarhizium anisopliae for Manduca sexta. J Invertebr Pathol,1988,52: 285-293.
Influence of Symbiotic Bacteria Arsenophonus,Rice Variety and Temperature on the Incidence Rate of Nilaparvata lugens to Metarhizium flavoviride
ZHU Huanhuan,CHEN Yang,WAN Pinjun,WANG Weixia,LAI Fengxiang,FU Qiang*
(State Key Laboratory of Rice Biology,China National Rice Research Institute,Hangzhou 310006,China; *Corresponding author,E-mail: fuqiang@caas.cn)
【Objective】 To reveal the significant factors affecting the incidence rate of the brown planthopper (BPH) to Metarhizium flavoviride,laboratory bioassays were carried out to study the influence of symbiotic bacteria Arsenophonus,rice variety and temperature on the incidence rate of BPHs to M. flavoviride.【Method】 Through two completely randomized block trial: 1) symbiotic bacteria Arsenophonus (infected and uninfected) versus rice varieties (TN1,IR56 and Mudgo),2) and Arsenophonus (infected and uninfected) versus temperatures (21°C,23°C,25°C,27°C,29°C and 31°C),BPHs with symptoms of M. flavoviride infection were observed after exposure to the spore suspensions of M.flavoviride for different periods.【Result】 For all tested rice varieties,the incidence rates of M. flavoviride-treated BPHs infected with Arsenophonus (BPH++) were lower than that of BPHs uninfected (BPH--). Out of tested temperatures,only under 25°C,27°C,29°C,BPH++ has lower incidence rates than BPH-- at 3 d and 5 d after spraying. However,there was no significant difference among others. The results of two-way ANOVA analysis between Arsenophonus and rice varieties also indicated that the incidence rate of BPH++ was lower than that of BPH--. Furthermore,two-way ANOVA results between Arsenophonus and temperatures indicated the incidence rate of BPH++ was lower than that of BPH-- at 3 d and 5 d after spraying. The effect of temperature was significant on the virulence of M. flavoviride to BPHs. BPHs had higher corrected incidence rates and shorter LT50under 23°C-29°C than that under 21°C or 31°C. Out of these BPHs,the highest corrected incidence rates and shortest LT50were found at 27°C. It suggested 23°C-29°C was suitable temperature for infection of M. flavoviride,among which 27°C was the best. This suggested that low or high temperatures would negatively affect the virulence of M. flavoviride to BPH. The temperature also affected the LT50of BPHs. The LT50of BPH++ was longer than that of BPH-- under 27°C or 29°C. However,the LT50of BPH++ was equal or even shorter than that of BPH-- under other temperatures.【Conclusion】 Arsenophonus decreased the virulence of M.flavoviride to BPH,which was influenced by temperature. The temperature also affects the incidence rate of BPHs to Metarhizium flavoviride significantly. However,the effects of rice varieties and interactions of Arsenophonus and rice varieties and Arsenophonus and temperatures were not significant on the incidence rate.
Nilaparvata lugens; Metarhizium flavoviride; Arsenophonus; temperature; rice variety
10.16819/j.1001-7216.2017.7012
2017-02-06; 修改稿收到日期:2017-05-04。
国家自然科学基金资助项目(31401738);中央级公益性科研院所基本科研业务费专项(2014RG005);农业现代产业技术体系资助项目(CARS-1-18);中国农业科学院科技创新工程水稻病虫草害防控技术创新团队项目。
S435.112+.3; S476.1; S511.02
A
1001-7216(2017)06-0643-09
