Fe2+和Mn2+对水稻根表铁膜及镉吸收转运的影响*
2017-10-18董明芳郭军康冯人伟王瑞刚丁永祯徐应明范稚莲
董明芳 郭军康 冯人伟 王瑞刚 丁永祯 徐应明 范稚莲
(1.广西大学农学院,广西 南宁 530003;2.农业部环境保护科研监测所农田重金属污染修复创新团队,天津 300191;3.陕西科技大学环境科学与工程学院,陕西 西安 710021;4.农业部环境保护科研监测所,天津 300191)
Fe2+和Mn2+对水稻根表铁膜及镉吸收转运的影响*
董明芳1,2郭军康3#冯人伟2王瑞刚2丁永祯4徐应明2范稚莲1
(1.广西大学农学院,广西 南宁530003;2.农业部环境保护科研监测所农田重金属污染修复创新团队,天津300191;3.陕西科技大学环境科学与工程学院,陕西 西安710021;4.农业部环境保护科研监测所,天津300191)
采用水培实验,研究外源添加Fe2+和Mn2+对水稻幼苗根表铁膜形成、抗氧化酶活性及镉吸收转运的影响。结果表明:(1)添加Fe2+使水稻幼苗根表铁膜中铁元素显著增加114.9%(质量分数,下同),锰和镉元素则分别显著降低了55.2%和42.3%(P<0.05);添加Mn2+使铁膜中锰元素显著增加180.1%,镉元素则显著降低了18.0%。(2)添加Fe2+能显著降低水稻幼苗叶片中活性氧(ROS),缓解ROS对水稻幼苗的毒害;添加Mn2+使得叶片中ROS、超氧化物歧化酶和谷胱甘肽转移酶增加,Mn2+处理引起的ROS升高可能起到诱导水稻胁迫应答,缓解镉毒害作用。
水稻 根表铁膜 铁 锰 镉
铁膜是水稻等植物适应环境胁迫的重要机制之一。有报道表明,铁膜能通过吸附和共沉淀等物理化学作用影响金属离子在土壤中的化学行为和生物活性[1]336。水稻在淹水条件下,根际土壤中大量的Fe2+和Mn2+等被水稻根部通气组织分泌的氧气氧化,形成一层覆盖在水稻根表及根质外体上的铁膜[2]。关于水稻根际铁膜的形成机制报道有很多。傅友强等[3]1528指出,水稻根表铁膜由结晶态和无定型态铁氧化物或氢氧化物等组成。陈学萍等[4]则发现,铁膜主要由水铁矿组成,并含有少量针铁矿和赤铁矿等晶体。根部氧化作用和根际Fe2+浓度是根表铁膜形成的两个重要条件,水稻根部氧化能力主要依靠根部氧化酶、根部分泌氧化性物质、氧气和根际氧化性微生物等[5]。水稻根部存在氧化物酶系,能把Fe2+氧化成Fe3+[1]338。
铁膜对植物根部吸收矿质元素的影响除与环境条件有关外,主要与铁膜的厚度和性质有关,适量铁膜可促进水生植物对磷、锌等养分的吸收,但铁膜过多会抑制根部对养分和污染物的吸收[3]1529,[6],这为降低水稻吸收富集镉提供新思路,即通过调节根表铁膜,控制对镉等重金属的吸收与转运[7-10],[11]434-436。
铁膜形成也受根际环境(CO2、pH、锰等)、水分和植物种类等因素的影响。研究发现,CO2促进根部铁膜(根尖除外)的形成[12]。在水培条件下,过量的CO2易于形成针铁矿,无CO2或低CO2时Fe2+快速氧化和水解,形成纤铁矿[13]。在一定pH(3~6)内,水培的植物铁膜数量与pH正相关。研究发现,芦苇生长在pH较高(6.0)条件下形成的铁膜较厚,pH较低(3.5)时形成的铁膜相对很薄[14-15]。锰的存在也增加了铁膜的吸附能力,锰氧化物虽然在铁膜中所占的比例不大,但锰膜比铁膜的表面活性和催化能力更强,对重金属的吸附更大[16-17]。
然而,外源添加Fe2+和Mn2+对根表铁膜形成及其对植物生长和生理影响的相关研究不多。本研究通过探讨外源添加Fe2+和Mn2+对水稻幼苗根表铁膜形成及其对镉吸收转运和水稻幼苗抗氧化酶活性的影响,为抑制重金属镉在水稻体内的迁移提供理论依据。
1 材料与方法
1.1实验材料
供试水稻品种:“扬稻6号”,江苏里下河地区农业科学研究所选育。
水培营养液为国际水稻培养液配方[18],其中镉元素为1.0mg/L(见表1)。实验设置3个处理(不添加Fe2+和Mn2+的处理记为CK;加入Mn2+至1.00mg/L记为TMn;加入Fe2+至4.00mg/L记为TFe),每个处理设置5次重复。
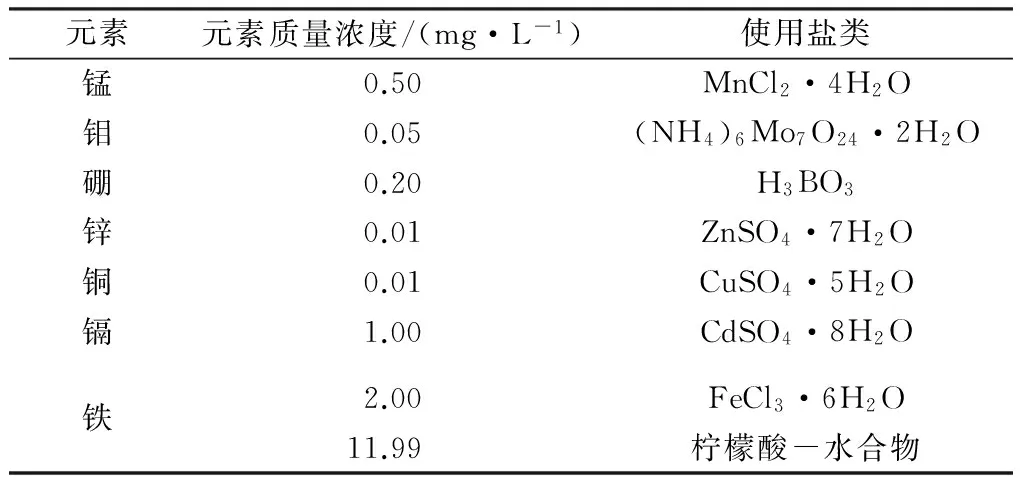
表1 营养液微量元素配方表
1.2 实验处理
挑选籽粒饱满的水稻种子用3%(质量分数)的NaClO消毒15 min后,将种子用去离子水清洗5~6次,将表面消毒后种子播种到以蛭石作为基质的穴盘中,将穴盘置于控温、控湿和固定光照时间的人工气候室内培养。培养条件:日光照时长为14 h,黑夜时长10 h,白昼时段温度为(26±2) ℃,黑夜时段温度为(20±2) ℃,湿度为60%。培养两周待水稻发育后,选取大小均一的四叶期水稻幼苗移栽至盛有1 L营养液的黑色聚氯乙烯(PVC)培养罐(高14 cm,底部直径11 cm)中进行1周适应性培养,再进行实验处理,每盆种植两株水稻苗,每周换营养液1次。上述处理置于人工气候室培养3周后收样。
1.3 测定项目及方法
1.3.1 水稻地上与地下部镉元素含量测定
从人工气候室水培条件下培养3周的水稻幼苗上取0.20 g左右叶片鲜样,液氮速冻后转移至-80 ℃冰箱低温保存待用。沿茎基部将水稻幼苗地上部和根部分开,用去离子水冲洗地上部3次,根部浸泡入5.0 mmol/L乙二胺四乙酸(EDTA)溶液中15 min,以去除根表附着镉,然后用去离子水冲洗5次,分别将地上部与地下部水稻样品装入纸袋,75 ℃杀青,65 ℃烘干至恒量待用。将上述水稻幼苗根部和地上部茎叶分别磨碎,称取0.25 g左右混匀后的样品,加入硝酸15 mL,摇匀静置6~8 h,然后放置于电热消解仪(DigiBlock ED54)中(80 ℃下1.5 h、120 ℃下1.5 h、150 ℃下3.0 h)消解,摘掉消解管盖子在175 ℃赶酸至体积<1 mL,消解液用超纯水定容至25 mL,过滤后用电感耦合等离子体(ICP)-质谱(MS)仪(iCAP-Qc,德国ThermoFisher)测定镉元素浓度[19]。空白和标准物质GBW07605(国家标准物质中心提供)与样品同步处理,以确保测定的准确度。
1.3.2 水稻叶片抗氧化酶测定
称取0.20 g左右叶片,液氮研磨,加入5 mL提取缓冲液(50 mmol/L磷酸钾缓冲液(pH=7)),1 mmol/L EDTA,1%(质量分数)PVPP-30,1 mmol/L抗坏血酸(仅在抗坏血酸过氧化物酶(APX)测定时加入),匀浆4 ℃,10 000 r/min离心10 min,去上清液待测。采用酶联免疫分析试剂盒[20],应用双抗体夹心法测定植物中的超氧化物歧化酶(SOD)、活性氧(ROS)、丙二醛(MDA)、谷胱甘肽转移酶(GST)、APX。用纯化的植物酶抗体包被微孔板,加入酶样品,再与HRP标记的酶抗体结合,形成抗体-抗原-酶标抗体复合物,彻底洗涤后加底物TMB显色,在HRP酶的催化下转成蓝色,在酸的作用下最终转化成黄色,用酶标仪(iMark,BIO-RAD)在450 nm波长下测定吸光度,并通过标准曲线计算样品酶浓度。
1.3.3 根表铁膜铁、锰和镉元素含量测定
将收获后的水稻幼苗根部用去离子水冲洗两遍,沥干水分后从茎基部将根剪下,置于盛有60 mL DCB提取液(0.03 mol/L柠檬酸三钠和0.125 mol/L NaHCO3)的100 mL烧杯中,同时加入Na2S2O41.0 g,充分混合均匀,室温下静置提取1 h,然后将浸提液过0.45 μm的针孔式水系滤膜,ICP-MS测定铁、锰和镉元素含量,或在4 ℃下保存。
1.4 数据分析
转移镉的能力用转运系数(TF)表示,其中水稻幼苗根表铁膜向地上部、根部向地上部、根表铁膜向根部的TF分别为TF-铁膜(即水稻幼苗地上部与根表铁膜的镉元素质量浓度比值)、TF-根(即水稻幼苗地上部与根部的镉元素质量浓度比值)、TF-铁膜-根(即水稻幼苗根部与根表铁膜的镉元素质量浓度比值)。
使用SPSS 16.0对数据进行统计分析和单因素方差分析,最小显著性差异法(LSD)设置为0.05检验显著性差异。
2 结果与分析
2.1水稻根表铁膜金属元素含量
根据表2,添加Fe2+显著提高了水稻根表铁膜中的铁元素浓度,比CK处理提高了114.9%,但显著降低了锰元素浓度,比CK处理低55.2%;添加Mn2+显著提高了锰元素浓度,比CK处理提高了180.1%,但对铁元素浓度没有显著影响。添加Fe2+和Mn2+均显著降低了镉元素质量浓度,分别比CK处理降低了42.3%、18.0%。

表2 添加Fe2+和Mn2+水培3周后水稻根表铁膜中的金属元素质量浓度1)
注:1)不同字母表示处理间差异显著(P<0.05),相同则差异不显著。图1、图2、表3和表4同。
2.2 水稻根部与地上部生物量及镉元素含量
根据图1,添加Fe2+和Mn2+显著促进水稻生长,根部生物量比CK处理分别增加了96.5%、83.0%,地上部生物量分别增加了43.1%、29.1%。

图1 添加Fe2+和Mn2+水培3周后水稻根部与地上部生物量变化Fig.1 The biomass of rice root and aerial part with different treatments after 3 weeks of hydroponic culture
根据图2,与CK处理相比,添加Fe2+处理后,根部镉元素无显著变化,而地上部镉元素显著降低16.8%;添加Mn2+处理后,根部镉元素显著增加了38.4%,而地上部镉元素无显著变化。
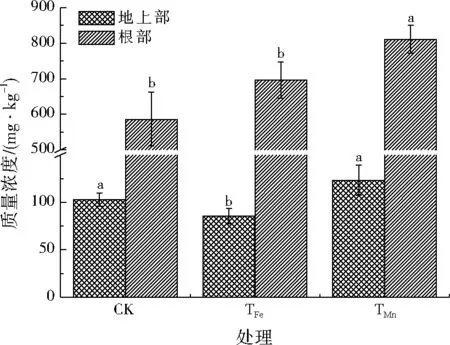
图2 添加Fe2+和Mn2+水培3周后水稻根部与地上部镉元素质量浓度变化Fig.2 The Cd concentrations in rice root and aerial part with different treatments after 3 weeks of hydroponic culture
由表3可见,与CK处理相比,添加Fe2+和Mn2+后TF-铁膜-根和添加Mn2+后TF-铁膜显著升高,添加Fe2+后TF-根显著降低。说明添加Fe2+和Mn2+促进了镉在根部的累积。

表3 镉由水稻根表铁膜、根部向地上部以及铁膜向根部的转运
2.3 水稻叶片抗氧化酶活性
由表4可见,与CK处理相比,添加Fe2+和Mn2+处理的SOD、APX变化不大,但ROS变化显著。与CK处理相比,添加Fe2+后ROS显著降低了84.7%,添加Mn2+后却显著增加了31.8%。添加Fe2+或Mn2+处理的GST与ROS变化趋势一致。与CK处理相比,添加Fe2+后GST降低了12.2%,添加Mn2+后却显著增加了8.1%。

表4 添加Fe2+和Mn2+水培3周后水稻叶片抗氧化酶活性1)
注:1)U为一定条件下、一定时间内将一定量的底物转化为产物所需的酶量;IU为国际标准化规定的一个酶活力单位,即在最适反应条件下,每分钟催化1 μmol底物转化为产物所需的酶量,1 IU=1 000 mIU。
3 讨 论
3.1Fe2+和Mn2+对水稻根部铁膜形成及镉吸收与转运的影响与原理
根际重金属元素能通过共用一些植物生长必需元素离子转运通道的方式进入植物体内。THOMINE等[21]研究发现,缺铁诱导拟南芥铁、锰转运蛋白基因AtNRAMP3表达量增加,导致拟南芥体内镉和锰累积量升高。本研究对水稻幼苗根表铁膜金属元素分析结果表明,与CK处理相比,添加Fe2+和Mn2+使得水稻幼苗根表铁膜铁、锰元素含量显著增加,而镉元素含量均显著降低(见表2)。外源Fe2+和Mn2+可能通过竞争根部表面及铁膜中二价金属离子吸附位点,减少镉在铁膜中累积。刘丹青等[22]研究发现,缺铁导致水稻根表铁膜数量显著下降,同时铁膜中镉含量升高。刘侯俊等[23]研究发现,水培条件下镉(1.0mg/L)处理时,添加铁处理组根表铁膜中镉低于未加铁处理组,与本研究结果相印证。
本研究中,添加Fe2+使得水稻幼苗地上部镉元素含量显著降低,而添加Mn2+显著增加水稻幼苗根部镉元素含量(见图2)。添加Fe2+和Mn2+后TF-铁膜-根均显著升高,造成更多的镉在水稻幼苗根部组织中累积;添加Fe2+后TF-根显著降低,从而减少镉从根部往地上部的转运,有助于降低镉对地上部组织器官的毒害。类似研究结果表明,水稻根表铁膜量与茎叶、籽粒镉含量呈极显著的负相关,根表铁膜抑制镉向水稻地上部转运[11]437。
3.2Fe2+和Mn2+对水稻叶片抗氧化酶等的影响与原理
高浓度的ROS会引起水稻器官与组织生物大分子的氧化损伤,而低浓度的ROS能在植物细胞信号转导途径中作为第二信使,介导对激素或环境胁迫的多种应答,提高植物重金属等环境胁迫抗性[24]。过量重金属胁迫能增加水稻体内ROS含量,进而对组织器官功能造成严重损害,抑制植物正常生长发育[25-26]。本研究中与CK处理相比,添加Fe2+能显著降低水稻幼苗叶片中ROS(见表4),缓解高浓度ROS对水稻幼苗毒害作用;添加Mn2+使得叶片中ROS、SOD和GST增加(见表4),Mn2+处理引起的ROS升高可能起到诱导水稻胁迫应答,提高SOD和GST活性来缓解镉毒害[27]。刘大丽[28]研究发现,水稻体内GST活性增加使得谷胱甘肽(GSH)含量增加,较高浓度GSH能促进植物螯合肽(PC)大量合成,参与体内镉解毒过程,从而缓解水稻镉毒害。与CK处理相比,添加Fe2+和Mn2+能显著增加水稻幼苗根部与地上部生物量(见图1)。水稻幼苗生物量的增加可能与添加Fe2+和Mn2+促进根表铁膜形成,改变抗氧化酶活性,缓解镉对水稻生理毒害有关。
4 结 论
(1) 添加Fe2+和Mn2+能显著增加水稻幼苗根部与地上部生物量。
(2) 添加Fe2+和Mn2+能促进水稻幼苗根表铁膜形成,显著增加TF-铁膜-根;而添加Fe2+后TF-根显著降低,添加Mn2+后TF-铁膜显著升高。
(3) 添加Fe2+和Mn2+对水稻幼苗叶片SOD、APX变化不大;添加Fe2+能显著降低水稻幼苗叶片中ROS,缓解ROS对水稻幼苗毒害;添加Mn2+使得叶片中ROS、SOD和GST增加,Mn2+处理引起的ROS升高可能起到诱导水稻胁迫应答,缓解镉毒害作用。
[1] 赵锋,王丹英,徐春梅,等.水稻氧营养的生理、生态机制及环境效应研究进展[J].中国水稻科学,2009,23(4).
[2] CROWDER A A,COLTMAN D W.Formation of manganese oxide plaque on rice roots in solution culture under varying pH and manganese (Mn2+) concentration conditions[J].Journal of Plant Nutrition,1993,16(4):589-599.
[3] 傅友强,于智卫,蔡昆争,等.水稻根表铁膜形成机制及其生态环境效应[J].植物营养与肥料学报,2010,16(6).
[4] 陈学萍,朱永官,洪米娜,等.不同肥处理对水稻根表铁和砷形态的影响[J].环境化学,2008,27(2):231-234.
[5] 郑芸芸,李忠意,李九玉,等.铁膜对水稻根表面电化学性质和氮磷钾短期吸收的影响[J].土壤学报,2015,52(3):690-696.
[6] XU Defu,XU Jianming,HE Yan,et al.Effect of iron plaque formation on phosphorus accumulation and availability in the rhizosphere of wetland plants[J].Water,Air,and Soil Pollution,2009,200(1):79-87.
[7] LIU Houjun,ZHANG Junling,CHRISTIE P,et al.Influence of iron plaque on up take and accumulation of Cd by rice (OryzasativaL.) seedlings grown in soil[J].Science of the Total Environment,2008,394(2/3):361-368.
[8] LIU Jianguo,CAO Changxun,WONG M H,et al.Variations between rice cultivars in iron and manganese plaque on roots and the relation with plant cadmium uptake[J].Journal of Environmental Sciences,2010,22(7):1067-1072.
[9] YE Z H,CHEUNG K C,WONG M H.Cadmium and nickel adsorption and uptake in cattail as affected by iron and manganese plaque on the root surface[J].Communications in Soil Science and Plant Analysis,2003,34(19/20):2763-2778.
[10] CHENG Hao,WANG Maoyi,WONG M H,et al.Does radial oxygen loss and iron plaque formation on roots alter Cd and Pb uptake and distribution in rice plant tissues?[J].Plant & Soil,2014,375(1/2):137-148.
[11] 胡莹,黄益宗,黄艳超,等.不同生育期水稻根表铁膜的形成及其对水稻吸收和转运Cd的影响[J].农业环境科学学报,2013,32(3).
[12] WANG T G,PEVERLY J H.Iron oxidation states on root surfaces of a wetland plant (Phragmitesaustralis)[J].Soil Science Society of America Journal,1999,63(1):247-252.
[13] ST CYR L,CROWDER A A.Iron oxide deposits on the roots ofPhragmitesaustralisrelated to the iron bound to carbonates in the soil[J].Journal of Plant Nutrition,1988,11(6/7/8/9/10/11):1253-1261.
[14] TAYLOR G J,CROWDER A A,RODDEN R.Formation and morphology of an iron plaque on the roots ofTyphalatifoliaL. grown in solution culture[J].American Journal of Botany,1984,71(5):666-675.
[15] BATTY L C,BAKER A J M,WHEELER B D,et al.The effect of pH and plaque on the uptake of Cu and Mn inPhragmitesaustralis(Cav.) Trin ex. Steudel[J].Annals of Botany,2000,86(3):647-653.
[16] ST CYR L,CROWDER A A.Manganese and copper in the root plaque ofPhragmitesaustralis(Cav.) Trin. ex Steudel[J].Soil Science,1990,149(4):191-198.
[17] YE Z H,CHEUNG K C,WONG M H.Copper uptake inTyphalatifoliaas affected by iron and manganese plaque on the root surface[J].Canadian Jouanal of Botany,2001,79(3):314-320.
[18] 毛达如.植物营养研究方法[M].北京:中国农业大学出版社,2002.
[19] 魏竞智,段妮.干、湿法消解-石墨炉原子吸收法测定茶叶中铅镉对比[J].广东微量元素科学,2014,21(3):7-12.
[20] 王敏.SOD1模型化合物调控干旱胁迫下水稻细胞的抗氧化活性[D].武汉:华中师范大学,2011.
[21] THOMINE S,LELIVRE F,DEBARBIEUX E,et al.AtNRAMP3,a multispecific vacuolar metal transporter involved in plant responses to iron deficiency[J].The Plant Journal,2003,34(5):685-695.
[22] 刘丹青,陈雪,葛滢.缺Fe预处理对Fe、Cd根际吸附与水稻吸收和转运的影响[J].农业环境科学学报,2014,33(2):224-230.
[23] 刘侯俊,李雪平,韩晓日,等.镉处理根表铁膜对水稻吸收镉、锰、铜、锌的影响[J].植物营养与肥料学报,2013,19(6):1356-1365.
[24] SCHELER C,DURNER J,ASTIER J.Nitric oxide and reactive oxygen species in plant biotic interactions[J].Current Opinion in Plant Biology,2013,16(4):534-539.
[25] 刘俊祥,许新桥,钱永强,等.多年生黑麦草抗氧化酶和植物络合素对Cd2+胁迫的应答[J].生态学杂志,2013,32(7):1787-1793.
[26] GILL S S,TUTEJA N.Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants[J].Plant Physiology and Biochemistry,2010,48(12):909-930.
[27] BAXTER A,MITTLER R,SUZUKI N.ROS as key players in plant stress signalling[J].Journal of Experimental Botany,2014,65(5):1229-1240.
[28] 刘大丽.谷胱甘肽合成相关酶在重金属污染生物修复中的分子机制及比较研究[D].哈尔滨:东北林业大学,2013.
EffectsofFe2+andMn2+onricerootironplaqueformationandCduptakeandtransportation
DONGMingfang1,2,GUOJunkang3,FENGRenwei2,WANGRuigang2,DINGYongzhen4,XUYingming2,FANZhilian1.
(1.DepartmentofAgriculture,GuangxiUniversity,NanningGuangxi530003;2.InnovationTeamofRemediationforHeavyMetalContaminatedFarmlandofChineseAcademicofAgriculturalSciences,InstituteofAgro-EnvironmentalProtection,MinistryofAgriculture,Tianjin300191;3.SchoolofEnvironmentalScienceandEngineering,ShaanxiUniversityofScinence&Technology,Xi’anShaanxi710021;4.Agro-EnvironmentalProtectionInstitute,MinistryofAgriculture,Tianjin300191)
In this study,exogenous Fe2+and Mn2+were used to study the influence of them on rice root iron plaque formation,antioxidase activities and Cd uptake and transportation in hydroponic culture. The results showed that the Fe concentration in root iron plaque was significantly increased by 114.9% (mass fraction,the same below) with exogenous Fe2+addition treatment,compared to controls (P<0.05). However,the Mn and Cd concentration were significantly decreased by 55.2% and 42.3%,respectively. Exogenous Mn2+addition significantly increased the Fe concentration by 180.1% in root iron plaque,while the Cd concentration was significantly decreased by 18.0% comparedto controls. The reactive oxygen species (ROS) in leaves of rice seedlings was significantly reduced with adding exogenous Fe2+and the toxic effect of ROS on rice seedlings was also reduced. The contents of ROS,superoxide dismutase and glutathione transferase were increased in the leaves with the adding exogenous Mn2+. The stress response of rice appeared with the increasing of ROS under the experiments of adding exogenous Mn2+and the toxicity of Cd was also reduced.
rice; root iron plaque; Fe; Mn; Cd
董明芳,女,1988年生,硕士研究生,研究方向为土壤环境生态。#
。
*国家自然科学基金资助项目(No.41001191、No.41473115);中央级公益性科研院所基本科研业务费专项资金(农业部环境保护科研监测所)资助项目(No.2015-szjj-gjk-01)。
10.15985/j.cnki.1001-3865.2017.03.005
2016-04-17)
