放牧对卧龙大熊猫栖息地草本植物物种多样性与竹子生长影响
2017-09-15黄金燕张明春周世强李仁贵张晋东周小平李德生张和民
黄金燕,刘 巅,张明春,谢 浩,周世强,李仁贵, 张晋东,周小平,黄 炎,李德生,张和民
(1.中国大熊猫保护研究中心,四川 都江堰 611830; 2.西华师范大学,四川 南充 637000)
放牧对卧龙大熊猫栖息地草本植物物种多样性与竹子生长影响
黄金燕1,刘 巅1,张明春1,谢 浩1,周世强1,李仁贵1, 张晋东2,周小平1,黄 炎1,李德生1,张和民1
(1.中国大熊猫保护研究中心,四川 都江堰 611830; 2.西华师范大学,四川 南充 637000)
该文研究了放牧对卧龙国家级自然保护区大熊猫(Ailuropodamelanoleuca)栖息地草本层植物物种组成、多样性、高度、数量的影响及主食竹生长对放牧的响应。结果表明:放牧改变了大熊猫栖息地草本层植物组成,增加了物种的多样性和集中性,但物种分布的均匀程度略有降低;受损大熊猫栖息地和未受损大熊猫栖息地草本植物群落植物组成中等相似;放牧对大熊猫主食竹冷箭竹的生长也造成了不利影响,基径、株高和生物量降低,与未受损大熊猫栖息地有极显著差异。
放牧;大熊猫栖息地;物种多样性;主食竹
大熊猫(Ailuropodamelanoleuca)是世界最可爱的动物之一,为中国所特有的珍稀濒危动物,现今除人工圈养外在自然界仅呈岛屿状分布于中国四川省的秦岭山系四川段、岷山山系四川段、邛崃山系、大相岭山系、小相岭山系和凉山山系及陕西省的秦岭山系陕西段、甘肃省的岷山山系甘肃段[1-3]。大熊猫分布区多为经济不发达地区,放牧一直是这些区域居民增加经济收入的传统方式之一,但现今不断扩大的放牧活动在不少地区和大熊猫保护出现了明显冲突,家畜放牧在目前的大熊猫分布地区普遍存在,其野外遇见率达到33.9%,居于人为干扰因子的第一位,而且海拔跨度较大,几乎覆盖了大熊猫的最适分布区[4-10]。牲畜进入大熊猫栖息地,大熊猫主食竹被取食,竹子出现生长减缓、枯萎,竹林杂草丛生,干扰严重的区域甚至成片死亡,无疑造成了大熊猫栖息地质量下降或丧失,压缩了野生大熊猫的生存空间,受损大熊猫栖息地的自然生态系统一定程度上出现了退化。受损大熊猫栖息地恢复重建受到各级政府、大熊猫保护机构与研究人员密切关注,对此采取了积极的保护措施,开展了大熊猫保护工程,并资助了受损大熊猫栖息地研究项目。
放牧对大熊猫栖息地的影响仅有少量研究报道,有的作为干扰因素被调查报道而没有详细研究[2-4,11-14],有的就放牧对大熊猫生境的影响[5,15-16]、大熊猫与家畜的生境选择[6]、大熊猫栖息地干扰调查[7]、大熊猫与家畜的竞争关系[8]、大熊猫与家畜的空间利用格局[17]等进行了具体而比较深入的研究,但放牧影响大熊猫栖息地植物物种组成、多样性及竹子生长指标方面的研究尚未被涉及。放牧对受损大熊猫栖息地的影响主要表现在草本层的物种组成及其数量特征和竹子生长上的变化,故本文对此进行分析。现就在卧龙自然保护区开展受损大熊猫栖息地恢复重建调查研究时所收集的有关放牧对大熊猫栖息地植物物种组成及竹子生长影响的数据整理分析报道于后,供大熊猫保护管理者决策和科研人员开展相关研究时参考。
1 研究区域的位置与自然地理概况
四川卧龙国家级自然保护区位于四川盆地向青藏高原过渡的高山峡谷区,行政区划上属于阿坝藏族羌族自治州四川汶川卧龙特别行政区,北纬30°45′-31°25′,东经102°52′-103°24′,始建于1963年,面积2 000 km2。保护区地势由东南向西北递增,最低点木江坪海拔1 150 m,最高峰四姑娘山6 250 m,东西长52 km,南北宽62 km。区内山高谷深、沟壑纵横、水流湍急,主要河流包括横穿保护区中心地带的西北至东南走向的皮条河、耿达河,北部区域的正河,以及南面的中河与西河。随着海拔高度的变化和山地气候效应的影响,区内不仅形成了明显的植被分布垂直带谱,从低海拔到高海拔递次是常绿阔叶林、常绿落叶阔叶混交林、落叶阔叶林、针阔混交林、亚高山暗针叶林、高山灌丛、高山草甸、流石滩稀疏植被,而且孕育了众多植物资源,保存了不少古老孑遗物种和特有物种,成为动植物广谱基因库和避难所,已发现植物4 000多种,昆虫1 700多种,脊椎动物450多种,其中兽类109种,鸟类298种[18-19];野生大熊猫种群数量约占全国的十分之一[2-3]。
在四川卧龙国家级自然保护区,大熊猫栖息地放牧主要发生在耿达镇“牛坪”至“九大包”一线山脊及其周边,幸福沟的“牛道坪”“黄草坪”“金鸡沟”及其周边,“七层楼沟”及其两边,“老鸦山沟”两边山上的“德拉姑”“傻傻姑”“水井湾”等地;卧龙镇的三村“鱼丝洞”牛场周边的“三岔沟”“白石头”“黄家沟”,“转经楼沟”至“牛头山”垭口沿线及周边的“爬爬沟”“空洞树”,“洞口”牛场周边,“五一棚大熊猫野外生态观察站”研究范围内的“方子棚”“芹菜湾”等地。
为研究受损大熊猫栖息地恢复重建机制,2015-2016年在保护区东南部受放牧影响的野生大熊猫核心栖息地“三岔沟”“白石头”和“牛头山”3个地域(图1)实施了相关试验研究,进行了样方调查。各区域放牧影响面积3~6 hm2不等,调查面积(含受损栖息地和未受损栖息地)均约为4.80 hm2。这些区域地形相对平缓,坡度10°~40°,海拔2 850~3 000 m;气候属于青藏高原气候区,夏季凉爽多雨,冬季寒冷干燥;植被为落叶阔叶林和针叶林,林下竹子均为冷箭竹(Bashaniafaberi)[18-20],对应的土壤类型为山地棕壤和山地亚高山暗针叶林土[21]。
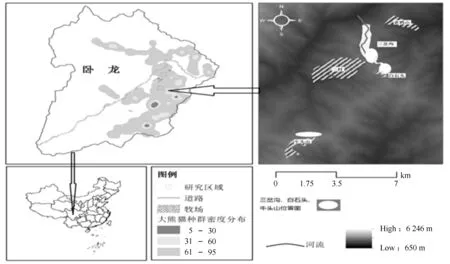
图1 研究区域位置Fig.1 The location of study area
放牧对该3个试验研究区域大熊猫栖息地的破坏主要发生在2008年地震后,产生的危害也最大。先是以马匹危害栖息地大熊猫主食竹为主,2012年12月马匹清除后变为牦牛、绵羊、黄牛等破坏栖息地为主,破坏的栖息地面积3~6 hm2不等。
2 研究方法
2.1 样方调查
在研究中,为比较被放牧破坏的大熊猫栖息地(受损栖息地)和未被放牧破坏的大熊猫栖息地(未受损栖息地)间植物群落物种组成和多样性的差异,了解放牧对大熊猫栖息地主食竹生长的影响,调查采用了“样方法”。在3个调查研究区域,分别未受损栖息地和受损栖息地,沿开展动物活动调查的样带上各设置20 m×20 m的固定样方各10个,得到未受损栖息地和受损栖息地各30个20 m×20 m植物群落植被特征调查样方。样方中分乔、灌、草3层记录反映植物群落植被特征的指标,鉴于放牧仅影响竹子的生长和草本层植物种类及其生长,这里就不对乔木层和灌木层群落植被特征的调查方法进行赘述。在草本层中,记录草本覆盖度(%)、植物名称、均高、多度等。在这些20 m×20 m样方的中心设置1 m×1 m的固定小样方,计60个;在3个区域受损大熊猫栖息地中各设置10个2 m×2 m的试验样方,2 m×2 m样方再分为4个1 m×1 m的小样方,计120个,所有小样方均调查记录冷箭竹当年生(1 a)、头年生(2 a)、多年生(≥3 a)各龄级竹子的数量并随机选择各龄级10株竹子测定高度(cm)、基径(mm)等。生物量的测算是在每个区域分未受损栖息地和受损栖息地按当年生、头年生和多年生3个龄级测定35株竹子的基径、高和生物量(g),建立竹子基径、高与生物量的回归方程,并由此根据小样方调查的竹子生长指标计算单位面积竹子的生物量。
2.2 数据整理分析
2.2.1 竹子生长调查数据的统计分析 调查数据的统计分析与整理均采用Microsoft Office Excel 2013软件和SPSS 19.0软件完成。
2.2.2 草本层群落物种多样性指标计算 群落的物种多样性是反应其组织水平的重要生态学特征之一,它是群落结构类型、组织水平、发展阶段、稳定程度和生境差异的体现[22]。群落的多样性指数由丰富度(绝对密度) 和均一性(相对密度)组成[23],是其种数、个体总数和均匀程度的综合概念[24],是把物种数和均匀度结合起来的一个单一的统计量[25]。为综合评价放牧对卧龙大熊猫栖息地草本层植物群落物种多样性的影响,这里采用了物种丰富度指数、物种多样性指数和均匀度指数测度群落的多样性,各计算公式[25-26]如下:
草本层植物物种重要值:重要值(IV)=(相对盖度RC+相对频度RF+相对高度RH)×100/3。物种丰富度:Gleason 物种丰富度指数dGL=S/lnA。物种多样性指数:Simpson指数D=1-ΣPi2。Shannon-Wiener指数H′=-ΣPilnPi。Pielou均匀度指数:JP=H′/lnS。
其中:S为草本层植物群落物种数目,A为样方面积,Pi物种i的相对重要值。
2.2.3 草本层物群落相似性系数计算 群落相似性是不同群落结构特征的相似程度。在植物物种构成相近的群落中,除用共有物种数量来比较群落的相似性外,各群落物种种数和群落共有物种种数的综合性参数——群落相似性系数较适宜表达群落间物种组成的相似程度。群落相似系数计算方法有多种,这里选用杰卡德(Jaccard)群落相似性系数Cj=j/(a+b-j)对未受损大熊猫栖息地与受损大熊猫栖息地的草本层植物群落进行相似性系数计算,以此数量指标判断这两种群落的相似性,式中j为群落A(未受损大熊猫栖息地草本层植物群落)、B(受损大熊猫栖息地草本层植物群落)共有物种数,a为A群落所有物种数,b为群落B所有物种数。
3 研究结果与分析
3.1 受损大熊猫栖息地和未受损大熊猫栖息地草本层植物群落物种组成及其数量特征
3.1.1 受损大熊猫栖息地和未受损大熊猫栖息地草本层植物群落物种组成差异 放牧对大熊猫栖息地草本层植物物种组成产生了影响。调查数据表明,未受损大熊猫栖息地草本层植物平均覆盖度20.00%,植物种类53种,分属23科41属,从各草本植物在样方中出现频率看,以早熟禾、钝叶楼梯草、蕨类、高原露珠草、四叶葎、六叶葎、散序地杨梅、四川碎米荠、凉山悬钩子、酢浆草、瓜叶乌头等植物为主,其余43种为少见种,代表了此区域大熊猫栖息地草本层植物的自然组成;而受损大熊猫栖息地草本植物物种有27科、54属71种,平均覆盖度达45.83%,其中样方中出现比较多的物种是早熟禾、四川碎米荠、六叶葎、高原露珠草、弹裂碎米荠、独叶草、珠芽艾麻、迎阳报春、四叶葎、瓜叶乌头、蕨类、白苞蒿、酢浆草、钝叶楼梯草、散序地杨梅、高乌头、长药隔重楼、卵叶韭、蛇果黄堇、繁缕、峨参叶紫堇、支柱蓼、耳翼蟹甲草、三脉紫菀、四叶重楼、鹿药等,其余46种为少见种,代表了此区域大熊猫栖息地受放牧影响后草本层植物组成的自然变化。从重要值来分析,未受损栖息地以钝叶楼梯草、凉山悬钩子、糙野青茅、蕨类、亲族薹草、早熟禾为主,而受损栖息地以钝叶楼梯草、四川碎米荠、早熟禾、凉山悬钩子、散序点地梅、卵叶韭、峨参叶紫堇、高乌头、蕨类占优势(表1)。大熊猫栖息地受到放牧影响后,竹子长势减弱甚至死亡,杂草得到了更大的生存与扩展空间,一些喜阳光草本植物种类得到发展或侵入,一些喜阴的草本植物受到抑制或灭亡但其变化无以改变整个草本层的发展趋势,由此草本植物种类和覆盖度增加。

表1 大熊猫栖息地草本层植物种类及其重要值

续表1
注:(1)表中列出了研究区域内种子植物的科、属的分布区类型,其名称前面的数字是其分布区类型[27-29];(2)未受损栖息地Intact Habitats(缩写为IH,下同),受损栖息地Damaged Habitats(缩写为DH,下同)。
划分和分析整理植物的分布区类型可以理解植物区系各种成分的特征与性质。从草本层种子植物物种所属26个科的分布区类型看,世界广布(即数字1)19个科,热带广布(即数字2)3个科,东亚(热带、亚热带)及热带南美间断(即数字3)1个科,北温带(即数字8)1个科,北温带和南温带间断分布(即数字8-4)2个科。
3.1.2 受损大熊猫栖息地和未受损大熊猫栖息地草本层植物群落物种多样性比较 生物群落是在一定地理区域内生活在同一环境下的不同种群的集合体,它在组成、结构、功能和动态方面表现出丰富多彩的差异即群落具多样性,从而在一定区域内呈现景观的异质性。群落多样性研究的理论意义在于认识群落的结构和功能,发现群落组成和结构的一般规律,了解群落动态的内在机制;群落多样性研究的实践意义主要包括保护与监测两个方面,在生物多样性保护实践中人们常以多样性指数为依据评价群落或生态系统的状况从而采取相应的保护措施[26]。
调查数据分析表明(表2),放牧会造成大熊猫栖息地草本层植物物种数目增加,物种丰富度指数、Shannon-wiener指数增大说明受损大熊猫栖息地草本植物群落的复杂程度增高;Simpson指数即生态优势度,它是多样性的反面即集中性的度量,它的微小增加表明放牧会造成大熊猫栖息地草本层植物物种集中性略微增大;Pielou均匀度指数降低,反映放牧使草本植物群落中物种多度分布的均匀度减小。
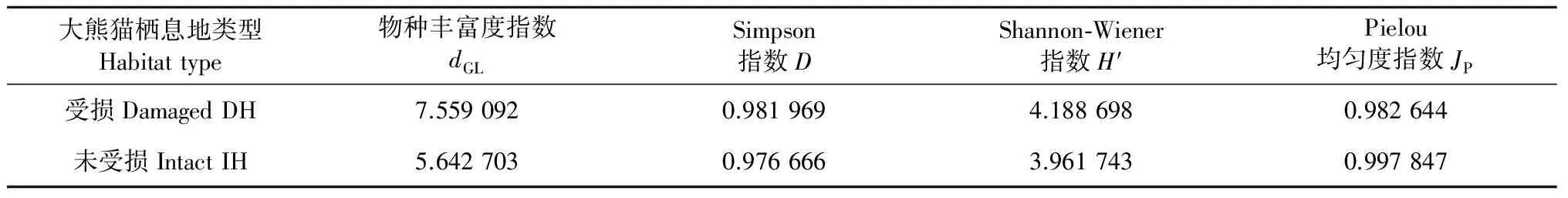
表2 不同大熊猫栖息地草本层植物群落物种多样性指数比较
3.1.3 受损大熊猫栖息地和未受损大熊猫栖息地草本层群落物种相似性 根据Jaccard相似性原理,当Cj为0~0.25时为极不相似,当Cj为0.25~0.50时为中等不相似,当Cj为0.50~0.75时为中等相似,当Cj为0.75~1.00时为极相似[30]。计算所得的Cj表明,受损大熊猫栖息地和未受损大熊猫栖息地草本层植物群落为中等相似,相似系数为0.590。这主要是由于放牧造成冷箭竹被啃食甚至死亡,即直接减少了现存竹子的空间占有量,又直接抑制了竹子的生长,微生境发生变化,原有植物间空间利用的平衡被打破,为新物种的入侵或原有物种的扩张生长增加了生存空间,使大熊猫栖息地草本植物种类和数量增加,草本植物群落物种异质性增大。
3.2 受损大熊猫栖息地和未受损栖息地冷箭竹生长发育差异
放牧对大熊猫栖息地的竹子生长有重要影响,竹子的基径、株高、生物量发生了明显变化(表3和图2)。
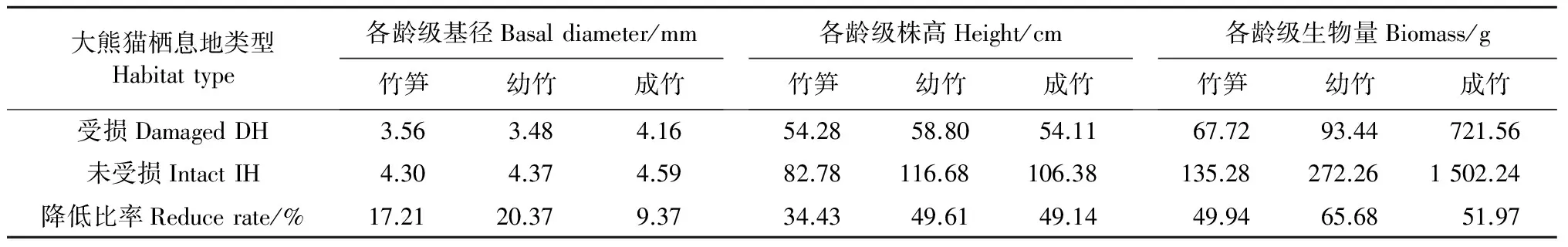
表3 不同大熊猫栖息地各龄级竹子生长参数
调查数据表明,受损大熊猫栖息地冷箭竹竹笋(当年生竹)、幼竹(头年生竹)、成竹(多年生竹)等各龄级竹子的基径、株高、生物量均值与未受损大熊猫栖息地有差异,通过显著性检验,受损大熊猫栖息地与受损大熊猫栖息地竹子生长具有极显著差异(P<0.01),其各龄级竹子基径、株高和生物量均大大减少,分别降低9.37%~20.37%、34.43%~49.61%、49.94%~65.68%。家畜在大熊猫栖息地啃食竹子,对竹子来说,一方面造成竹子高度变矮,另一方面使竹子进行光合作用的主要器官——竹叶大大减少甚至完全丧失;竹子高度变矮主要表现在2种类型栖息地中幼竹与成竹植株高度有极大的差异,竹笋刚出土不久,在6月份调查时牲畜的影响微弱,竹子高度上的差异不特别大,而竹叶大量丧失直接影响了光合产物的数量,进而影响竹子高度、基径的增长。同时,牲畜啃食竹子后造成杂草种类与数量增加,和竹子竞争营养空间,进而也影响着竹子高、径生长。
图2 受损大熊猫栖息地与未受损大熊猫栖息地竹子生长指标比较Fig.2 The bamboo growth index in intact vs damaged habitats of Giant Panda
[1] 胡锦矗.大熊猫研究[M].上海:上海科技教育出版社,2001:167-184.
[2] 国家林业局.全国第三次大熊猫调查报告[M].北京:科学出版社,2006:10-188.
[3] 四川省林业厅.四川的大熊猫——四川省第四次大熊猫调查报告[M].成都:四川科学技术出版社,2015:2-162.
[4] 古晓东,王鸿加,刘富文.岷山山系大熊猫自然保护区2003年生物多样性监测[J].四川动物,2005,24(2):168-170.
[5] 冉江洪,刘少英,王鸿加,等.放牧对冶勒自然保护区大熊猫生境的影响[J].兽类学报,2003,23(4):288-294.
[6] 冉江洪,刘少英,王鸿加,等.小相岭大熊猫与放牧家畜的生境选择[J].生态学报,2003,23(11):2253-2259.
[7] 冉江洪,刘少英,王鸿加,等.小相岭大熊栖息地干扰调查[J].兽类学报,2004,24(4):277-281.
[8] 康东伟,赵联军,宋国华.四川王朗国家级自然保护区大熊猫与家畜竞争关系[J].东北林业大学学报,2011,39(7):74-76.
[9] Hull Vanessa,Zhang Jindong,Zhou Shiqiang,etal.Space use by endangered giant pandas[J].Journal of Mammalogy,2015,96(1):230-236.
[10] 唐小平,贾建生,王志臣,等.全国第四次大熊猫调查方案设计及主要结果分析[J].林业资源管理,2015(1):13-16.
[11] 李晓鸿.甘肃大熊猫栖息地干扰现状分析[J].中国科技信息,2008(13):18-19.
[12] 田联会,樊金拴,王秋玲,等.秦岭大熊猫栖息地保护现状、问题与对策[J].西北林学院学报,2009,24(5):114-117.
[13] 刘明时,代拴发.太白山自然保护区大熊猫及其栖息地的干扰因素及保护对策[J].陕西师范大学学报(自然科学版),2006,3(专辑):207-210.
[14] 赵清,杨佳,杨兴中,等.太白山自然保护区大熊猫栖息地干扰因素调查[J].林业勘查设计,2007(4):63-66.
[15] 周世强,张和民,杨建,等.人为活动对大熊猫生境及其种群动态影响的研究[G].//汉桥,欧晓昆.生物圈保护区生物多样性保护与可持续发展.昆明:云南大学出版社,2004:110-114.
[16] Hull Vanessa,Zhang Jindong,Zhou Shiqiang,etal.Impact of livestock on giant pandas and their habitat[J].Journal for Nature Conservation,2014,22:256-264.
[17] 周世强,Hull Vanessa,张晋东,等.野生大熊猫与放牧家畜的空间利用格局比较[J].兽类学报,2016,36(2):138-151.
[18] 卧龙自然保护区管理局,南充师范学院生物系,四川省林业厅保护处.卧龙植被及资源植物[M].成都:四川科学技术出版社,1987:1-194.
[19] 卧龙自然保护区,四川师范学院.卧龙自然保护区动植物资源及保护[M].成都:四川科学技术出版社,1992:3-325.
[20] 易同培.四川竹类植物志[M].北京:中国林业出版社,1997:310-313.
[21] 张万儒.卧龙自然保护区的森林土壤及其垂直分布规律[J].林业科学,1983,19 (3):254-268.
[22] 黄忠良,孔国辉,何道泉.鼎湖山植物群落多样性的研究[J].生态学报,2000,20(2):193-198.
[23] 岳天祥.生物多样性研究及其问题[J].生态学报,2001,21(3):462-467.
[24] Whittaker R H.Evolution of diversity inplant communities[J].Ecology,1969,50:417-428.
[25] 中国科学院生物多样性委员会.生物多样性研究专著1.生物多样性研究的原理与方法[M].北京:中国科学技术出版社,1994:141-165.
[26] 马克平,黄建辉,于顺利,等.北京东灵山地区植物群落多样性的研究[J].生态学报,1995,15(3):268-277.
[27] 吴征镒,周浙昆,李德铢,等.世界种子植物科的分布区类型系统[J].云南植物研究,2003,25(3):245-257.
[28] 吴征镒.世界种子植物科的分布区类型系统的修订[J].云南植物研究,2003,25(5):535-538.
[29] 吴征镒.中国种子植物属的分布区类型[J].云南植物研究,1991,13(S4):1-139.
[30] 梅象信,徐正会,张继玲,等.昆明西山森林公园东坡蚂蚁物种多样性研究[J].林业科学研究,2006,19(2):170-176.
Grazing Impacts on the Herbaceous Species Diversity and Bamboo Growth in the Giant Panda Habitat in Wolong National Natural Reserve
HUANG Jin-yan1,LIU Dian1,ZHANG Ming-chun1,XIE Hao1,ZHOU Shi-qiang1,LI Ren-gui1, ZHANG Jin-dong2,ZHOU Xiao-ping1,HUANG Yan1,LI De-sheng1,ZHANG He-min1
(1.China Conservation Research Center for the Giant Panda,Dujiangyan 611830,Sichuan,China;2.West China Normal University,Nanchong 637000,Sichuan,China)
The grazing impacts on the species composition,diversity,height,number of herbs and the growth of staple food bamboo of Giant Panda (Ailuropodamelanoleuca) were studied in Wolong National Nature Reserve,a habitat for panda.The results showed that the grazing changed the composition of herbaceous species of the herb layer,increased species diversity and concentration .On the other hand,the evenness degree of species distribution was slightly decreased.The species composition of herbaceous plant communities in the damaged habitat of the giant panda was moderately similar to that of the intact one.The grazing significantly hampered the growth of arrow bamboo (Bashaniafaberi),its basal diameter,stem height and biomass in the grazing area were all significantly lower than those under control.
Grazing; Habitat of the giant panda; Species diversity; Staple food bamboo
2016-11-23
香港特别行政区支援四川灾后重建川港卧龙自然保护区持续合作项目;香港海洋公园保育基金资助项目(HK03)
黄金燕,教授级高级工程师,从事以大熊猫为主的保护生物学、生态学及圈养大熊猫野化放归研究。E-mail:huangjinyanabc@sina.com
