生物转化γ-氨基丁酸酿酒酵母的筛选及其在桑葚酒酿造中的应用
2017-07-31曾林谭霄张庆杨颖唐洁
曾林,谭霄,张庆*,杨颖,唐洁
1(西华大学 食品与生物工程学院,四川省食品生物技术重点实验室,四川 成都,610039)2(西华大学 古法发酵(酿造)生物技术研究所,四川 成都,610039)3(山东省食品药品检验研究院,山东 济南,250101)
生物转化γ-氨基丁酸酿酒酵母的筛选及其在桑葚酒酿造中的应用
曾林1,2,谭霄1,张庆1,2*,杨颖3,唐洁1
1(西华大学 食品与生物工程学院,四川省食品生物技术重点实验室,四川 成都,610039)2(西华大学 古法发酵(酿造)生物技术研究所,四川 成都,610039)3(山东省食品药品检验研究院,山东 济南,250101)
为获得产γ-氨基丁酸(γ-aminobutyric,GABA)并酿造桑葚酒性能良好的酵母菌,采用纸色谱定性分析从四川泡菜中分离筛选出3株产GABA酵母菌,经生理生化和18S rRNA测序分析,菌株JM009和JM037被鉴定为酿酒酵母(S.cerevisiae),菌株JM024被鉴定为C.tanzawaensis。通过高效液相色谱对酵母菌GABA表达能力评估发现,酿酒酵母 JM037发酵产GABA的能力较强,达670 mg/L。进一步对其进行耐受性和桑葚发酵性能分析,结果表明,酿酒酵母JM037对酒精、SO2、葡萄糖和pH的综合耐受性良好,并在发酵桑葚果汁后,乙醇产量在168 h达到38.36 g/L,残糖质量浓度在216 h降至3.06 g/L,GABA质量浓度在144 h提高到1 145 mg/L,以安琪酵母为参考,乙醇产量和残糖质量浓度差异不大,但酿酒酵母JM037具有独特的产GABA能力。因此,酿酒酵母 JM037具有酿制富含GABA桑葚酒的潜力。
γ-氨基丁酸;酿酒酵母;耐受性;筛选;鉴定;桑葚酒
γ-氨基丁酸(γ-aminobutyric,简称GABA)是由谷氨酸脱羧生成的一种非蛋白质类氨基酸,广泛存在于微生物、植物和动物中[1]。在脊椎动物中,GABA以一种重要的抑制性神经递质存在于中枢神经系统中[2-3],具有降低血压、利尿、增强记忆等多种生理功能[4-5]。随着人们生活水平的提高和保健意识的增强,富含GABA 的功能性食品已经成为研究和开发的热点。
神经生理及神经医学的研究表明,在人脑中,GABA是一种重要的活性物质,可由脑部的谷氨酸在谷氨酸脱羧酶(GAD)作用下转化而成,但随着年龄的增长和精神压力的加大,GABA的转化和积累比较困难,而通过日常饮食补充可有效改善这种状况[6]。1994年,TAKAYO等[7]在研究水浸泡处理米胚芽的氨基酸分布时,发现经过发酵处理的米胚芽中,GABA含量积累到200~300 mg/kg。最近,日本科学家已利用米胚芽等原料开发制造的富含GABA的功能食品配料,广泛地应用于饮料、果酱、调味料等制品中[8]。此外,还有报道采用微生物发酵法开发高浓度GABA饮料,以及利用乳酸菌等制作的富含GABA功能食品,可以直接作为抗高血压,改善脑机能及肝功能的功能食品。KIM等[8]利用短乳杆菌发酵制备富含GABA黑树莓果汁,GABA含量为505 mg/kg;NEJATI等[9]利用植物乳杆菌发酵制备富含GABA酸奶,GABA含量为77.4 mg/kg;在我国,有关GABA食品的研究报道较少,吴岱熹等[10]利用米曲霉和大米为原料制备富含GABA清酒,GABA含量为439 mg/L。但是对于产GABA酵母菌筛选,并应用于桑葚酒的酿造,制备富含GABA桑葚酒尚未报道。
本研究利用纸色谱定性分析筛选产GABA酵母菌,并对其进行生理生化试验和分子鉴定,再通过HPLC对酵母菌发酵产GABA的能力进行评估,进一步对高产GABA酵母菌进行耐受性和桑葚酒初步发酵性能分析。这不仅拓宽了产GABA菌株资源,而且为后续开发富含GABA桑葚酒提供有效数据。
1 材料与方法
1.1 材料与仪器
泡菜液,取于四川新繁食品有限公司;PDA培养基(马铃薯 200 g,葡萄糖 20 g)、YEPD培养基(葡萄糖 20 g,蛋白胨 20 g,酵母膏10 g)、酵母合成培养基(葡萄糖20 g,酵母提取物5 g,蛋白胨l0 g),pH均调节至pH6.0;GABA(含量≥99%),上海金穗生物科技有限公司;L-谷氨酸钠(L-MSG,含量≥98.5%),成都市科龙化工试剂厂;DH5α感受态细胞,TaKaRa Biotechnology(Dalian);pGEM-T载体,天根生化科技有限公司;安琪酿酒酵母,安琪酵母股份有限公司。
Allegra X-15R冷冻离心机,贝克曼库尔特公司;720BR/01492电泳凝胶成像分析系统,Biometra T1 PCR仪,BIO-RAD公司;BHC-1300ⅡA2生物安全柜,苏州安泰空气技术有限公司;Waters2695液相色谱仪(配有Waters2998紫外检测器),美国Waters公司。
1.2 酵母菌株的分离
取泡菜汁或各种发酵蔬菜样品25 g,加入225 mL无菌生理盐水的均质袋中,用拍击式均质器Bagmixer均质5 min。取适量均质液用无菌生理盐水10倍梯度稀释后涂布于酵母合成培养基进行分离,28 ℃培养48 h,挑取单菌落划线纯化3次。
1.3 产GABA酵母菌菌株的初筛
酵母菌菌株接种到PDA液体培养基培养24 h作为发酵种子,按4%的接种量接入50 mL YEPD(含1%L-MSG)的250 mL三角瓶中,28 ℃、120 r/min发酵72 h。发酵液于8 000 r/min条件下离心5 min,取上清液,备用。
采用纸色谱定性分析法初筛产GABA酵母菌。展开剂组分:V(正丁醇)∶V(冰乙酸)∶V(水)=5∶3∶2,再添加1.2%茚三酮。以L-MSG标品溶液(5 g/L)和GABA标品溶液(5 g/L)为对照,吸取2 μL发酵液,在滤纸上逐个点样,并编号。再将滤纸在30 ℃恒温展开,展开后将滤纸放进70 ℃烘箱中显色10 min[11]。
1.4 生化特征分析
按照《酵母菌的特征与鉴定手册》[12]中的鉴定方法进行。
1.5 生化特征分析
根据李可[13]等的方法提取真菌DNA。18S rRNA扩增引物:18S-F(5’-AACCTGGTTGATCCTGCCAGT-3’) 和18S-R(5’-TGATCCTTCTGCAGGTTCACCTAC-3’)。扩增条件:94 ℃预变性5 min,94 ℃变性1 min、55 ℃退火1 min、72 ℃延伸55 s,36循环后72 ℃保持10 min。
扩增产物连接到pGEM-T 载体后克隆入感受态细胞E.coliDH5α中,提取重组质粒对18S rRNA测序。所得序列提交至NCBI进行Blast比对,利用MEGA 5.0软件构建系统发育树。
1.6 GABA表达能力评估
利用HPLC对初筛菌株产GABA能力进行复筛。参照张术聪等[14]的方法对发酵液中GABA进行柱前衍生。HPLC分析条件:色谱柱为岛津-GL INERTSIL ODS-3(4.6 mm×150 mm,5 μm);紫外检测波长254 nm;柱温30 ℃;进样量20 μL;流动相A为甲醇,流动相B为醋酸钠(pH 6.2)∶甲醇∶四氢呋喃(84∶25∶1,V/V/V);流速1 mL/min;梯度洗脱时,流动相 B比例为:0~6 min,80%~50%;6~9 min,50%~20%;9~10 min,20%~0%;10~11 min,维持0%;11~15 min,返回80%。
1.7 耐受性分析
采用杜氏管发酵法测定酵母菌对乙醇、SO2、葡萄糖和pH耐受性。在10 mL YEPD 液体培养基的带杜氏管的试管中,分别加入不同酒精梯度( 6%、8%、10%、12%、14%、16%、18%和20% vol)、不同SO2质量浓度(100、150、200、250、300、350和400 mg/L)、不同葡萄糖质量分数(10%、20%、30%、40%、50%、60%和70 %)、不同pH(1.5、2.0、2.5、3.0、3.5、4.0和4.5),并分别接入产GABA酵母菌(接种量为1×108CFU/mL)28 ℃恒温静置培养5 d,以安琪酵母菌为对照,观察杜氏管中气泡产生情况[15]。
1.8 发酵性能分析
以安琪酵母菌为参考,将筛选酵母菌接种到PDA液体培养基培养24 h作为发酵种子,按6%的接种量接种于250 mL pH 3.75的灭菌桑葚汁(含5 g/LL-MSG和100 mg/L SO2)中,用灭菌蔗糖溶液调节糖度至20 °Brix后,28 ℃静置培养,当残糖含量为8%~9%时,转移至15 ℃进行低温陈酿,至残糖质量浓度降至4 g/L以下陈酿结束。每隔12 h取样,测定不同取样时间样品的生物量浓度、乙醇质量浓度、残糖质量浓度和GABA质量浓度。
生物量浓度:采用分光光度法进行检测,以OD600值和干重之间的关系曲线(1 OD600nm= 0.45 g/L)进行计算[16]。
乙醇质量浓度:采用气相色谱法,参照谢文等[17]在检测葡萄酒果酒等低含量酒精度的样品时采用甲醇稀释法,色谱条件:初始温度45 ℃,保持5 min,以3 ℃/min 升温至60 ℃,保持3 min,以20 ℃/min升温至200 ℃,保持5 min。色谱柱为Rtx-Wax毛细管柱(30 m×0.25 mm×0.25 μm),载气为高纯度氦气,载气流速1.0 mL/min;进样口温度:200 ℃,分流比为1∶20。以正丙醇为内标物,用色谱级甲醇稀释2倍后注入色谱柱进行分析。
残糖质量浓度:按照GB /T15038—2006的方法进行检测。
GABA质量浓度:同试验方法1.6。
2 结果与分析
2.1 产GABA酵母菌的初筛结果
从图1可以看出,利用纸色谱定性方法从样品中分离的67 株酵母菌菌株中筛选出3 株产GABA酵母菌,且底物L-MSG消耗明显。根据茚三酮显色反应的颜色深度初步判断,酵母菌JM037发酵液中GABA含量较高。

a-JM009; b-JM024; c-JM037; d-GABA标准品; e-L-MSG标准品图1 菌株发酵液纸色谱图Fig.1 Chromatography of fermentation broth
2.2 产GABA酵母菌的生理生化及系统发育树的构建
对筛选出的3 株产GABA酵母菌进行生理生化试验,结果见表1。查阅《酵母菌的特征与鉴定手册》,推测菌株JM009,JM037为酵母属酿酒酵母(S.cerevisiae);菌株JM024为C.tanzawaensis。为了更明确显示各菌株的分类学地位和系统发育关系,3 株菌株全部进行测序,测得序列经Mallard 1.02检测后,再通过软件BioEdit 7.0检测,无异常序列发现。选取相似菌株和代表菌株序列,用MEGA5.0软件以Neighbor-Joining法构建系统发育树。结果见图2,JM009和JM037被鉴定为S.cerevisiae;JM024被鉴定为C.tanzawaensis,与生理生化试验结果相符。
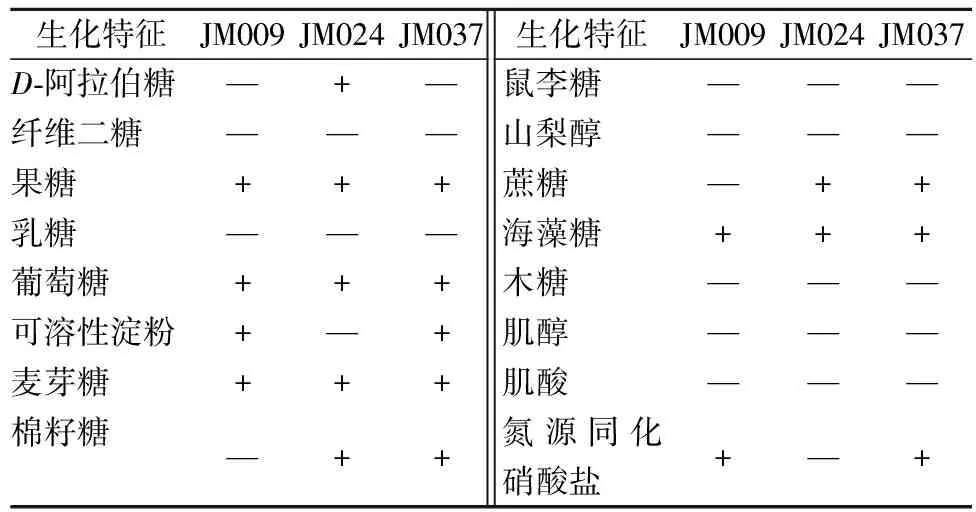
表1 生理生化试验结果
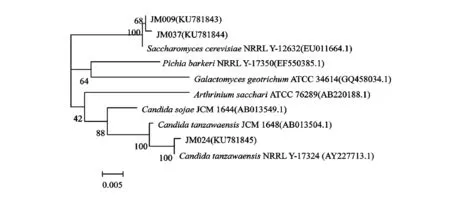
图2 基于酵母菌18S rRNA基因序列构建的系统发育树Fig.2 Phylogenetic tree of yeast based on 18S rRNA gene sequence
2.3 GABA表达能力评估
利用HPLC定量分析对酵母菌产GABA能力进行评估。其标准曲线为Y=9E+06X-18007(R2=0.999 2)。因此,可用于酵母菌发酵液中的GABA进行定量分析。3 株酵母菌GABA高效液相定量结果如图3所示。酵母菌JM037发酵产γ-氨基丁酸质量浓度最高,达670 mg/L。李亚莉等[18]从传统发酵食品分离出1株假丝酵母,发酵液GABA质量浓度达700 mg/L;乌云达来等[19]从中国科学院微生物研究所工业微生物与生物技术研究室保藏菌株中分离出1株酿酒酵母,发酵液中GABA质量浓度约480 mg/L;因此,可进一步对筛选获得的2 株酿酒酵母进行耐受性和桑葚酒发酵性能分析。
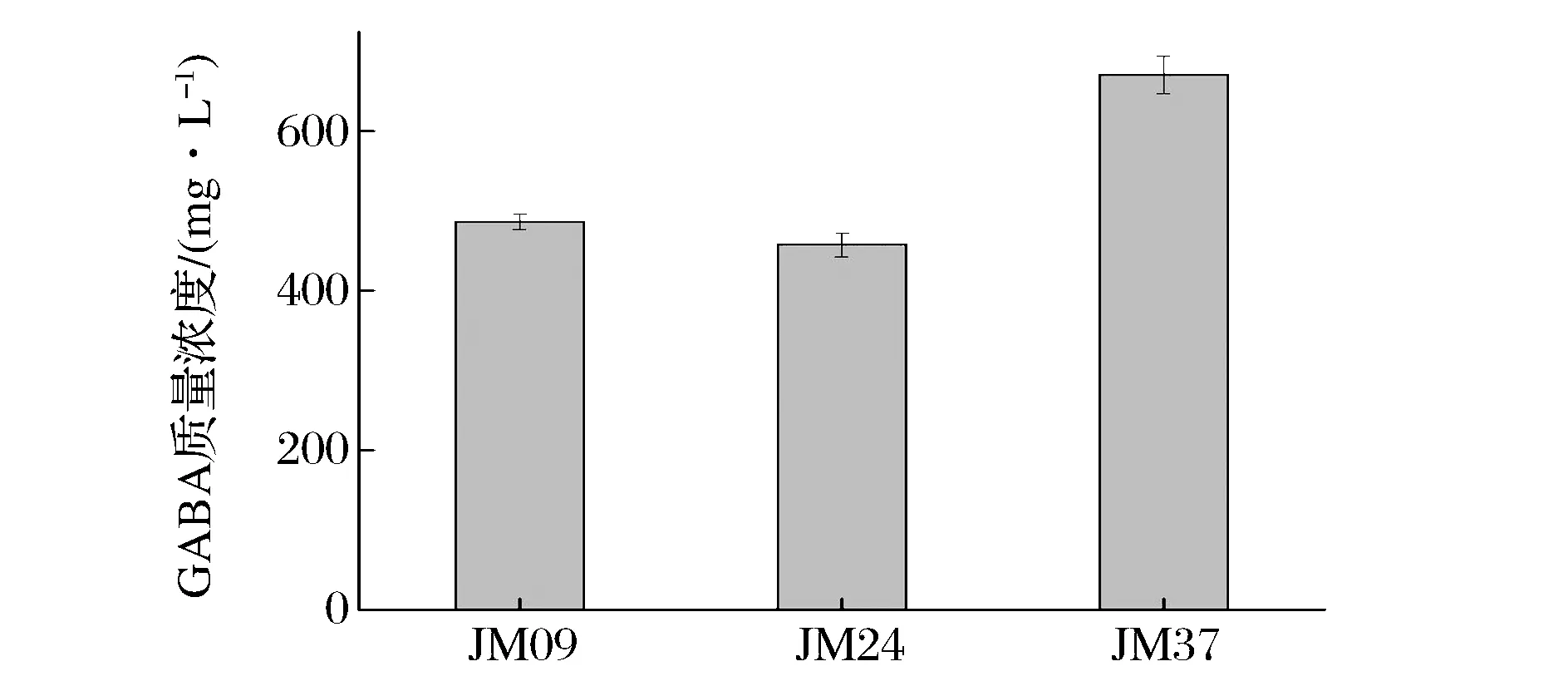
图3 菌株发酵液中GABA的质量浓度Fig.3 GABA mass concentration in the liquid medium fermented by the isolates
2.4 产GABA酵母菌耐受性
2.4.1 酒精耐受性
从贾春凤等[20]研究中可以看出,安琪酵母菌对酒精有较好的耐受性。以安琪酵母菌为参考,由表2可知,C.tanzawaensisJM024较敏感,在酒精含量为6%vol时气体产生较少,而酿酒酵母 JM009和JM037分别在酒精含量为12%vol和14%vol时产生大量气体,并且酿酒酵母JM009在酒精含量为18%vol时也有气体产生,说明酿酒酵母 JM009和JM037分别所耐受的最高酒精含量为12%vol和14%vol。高玉妹等[21]研究发现,酒精含量对酵母的生长有一定的影响,在发酵初期,低含量的乙醇对酿酒酵母有积极的影响,促进发酵顺利进行,当乙醇含量达到一定量(通常10% vol)时,又会对酿酒酵母产生毒害,延缓发酵。在果酒的酿制中,酒精含量一般在7%~18% vol,因此,酿酒酵母 JM009和JM037能应用于低度果酒发酵。
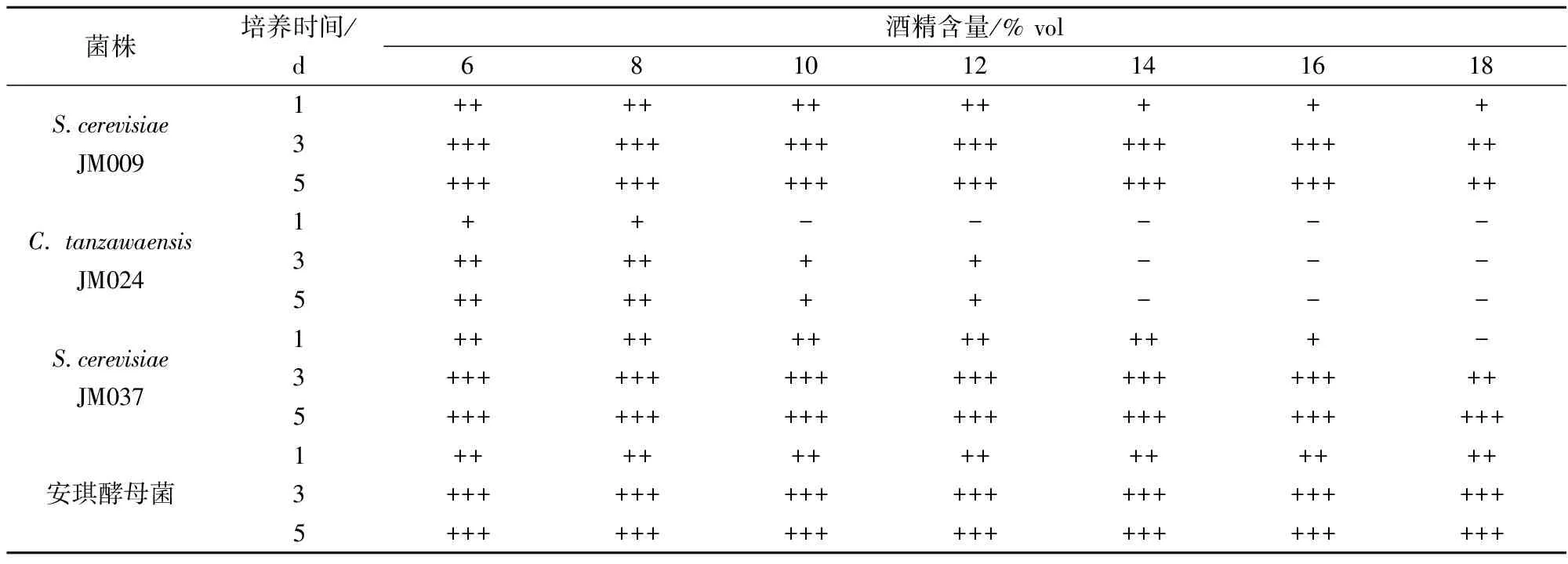
表2 产GABA酵母菌酒精耐受性结果
注: +: 杜氏管内满1/4管; ++: 满1/2管; +++; 满管。
2.4.2 SO2耐受性
果酒在酿制过程中会加入SO2,在抗氧化、杀菌和护色等方面起到关键的作用[22],一般添加量为50~150 mg/L,因此要求酵母菌菌株具有较高SO2耐受性。以安琪酵母菌为参考,由表3可知,酿酒酵母 JM037在SO2质量浓度达到250 mg/L时,有大量气体产生,酿酒酵母JM009在SO2质量浓度达到300 mg/L时,也有较多的气体产生,说明酿酒酵母JM009和JM037对SO2耐受性较好。
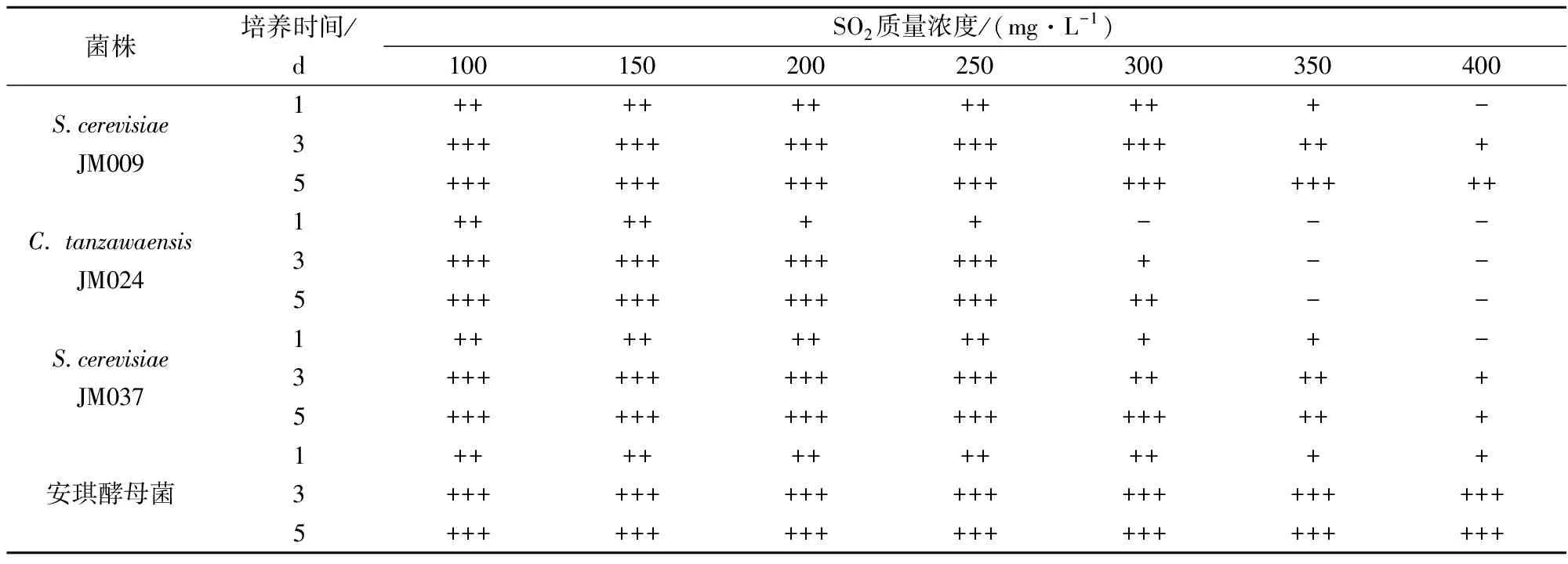
表3 产GABA酵母菌SO2耐受性结果
注: +: 杜氏管内满1/4管; ++: 满1/2管; +++: 满管。
2.4.3 葡萄糖耐受性
在果酒酿造中,糖是酒精发酵的基质,也是酵母菌赖以生长的能源物质,但高质量浓度的糖会对酵母菌的生长和代谢产生抑制,同时,高渗透压会导致酵母细胞水分流失,活性降低[23]。由表4可知,酿酒酵母JM009和JM037均能在葡萄糖质量浓度为300 g/L时产生大量气体,随着葡萄糖质量浓度的增加,气体产量减少,当葡萄糖质量浓度达到600 g/L时无气泡产生。说明酿酒酵母JM009和JM037能耐受300 g/L葡萄糖,略弱于安琪酵母菌的耐受性。于洋等[22]从酿酒葡萄中筛选的1株季也蒙有孢汉逊酵母YF-28能耐受250 g/L葡萄糖,一般果酒酿造的果汁中糖含量大约在250 g/L左右(按葡萄糖计)。本实验筛选的酿酒酵母JM009和JM037在果酒的酿造上具有应用潜力。

表4 产GABA酵母菌葡萄糖耐受性结果
注: +: 杜氏管内满1/4管; ++: 满1/2管; +++: 满管。
2.4.4 低pH耐受性
在低pH环境下,可抑制大部分有害微生物的生长代谢,对低pH耐受性较好的酵母菌菌株应用范围必定较广。由表5可知,酿酒酵母JM009和JM037能耐受pH 2.5以上的培养液,与安琪酵母菌相比,对低pH耐受性较差,但能保证大部分果酒发酵过程中,发酵液在pH在3.5左右下生长良好。
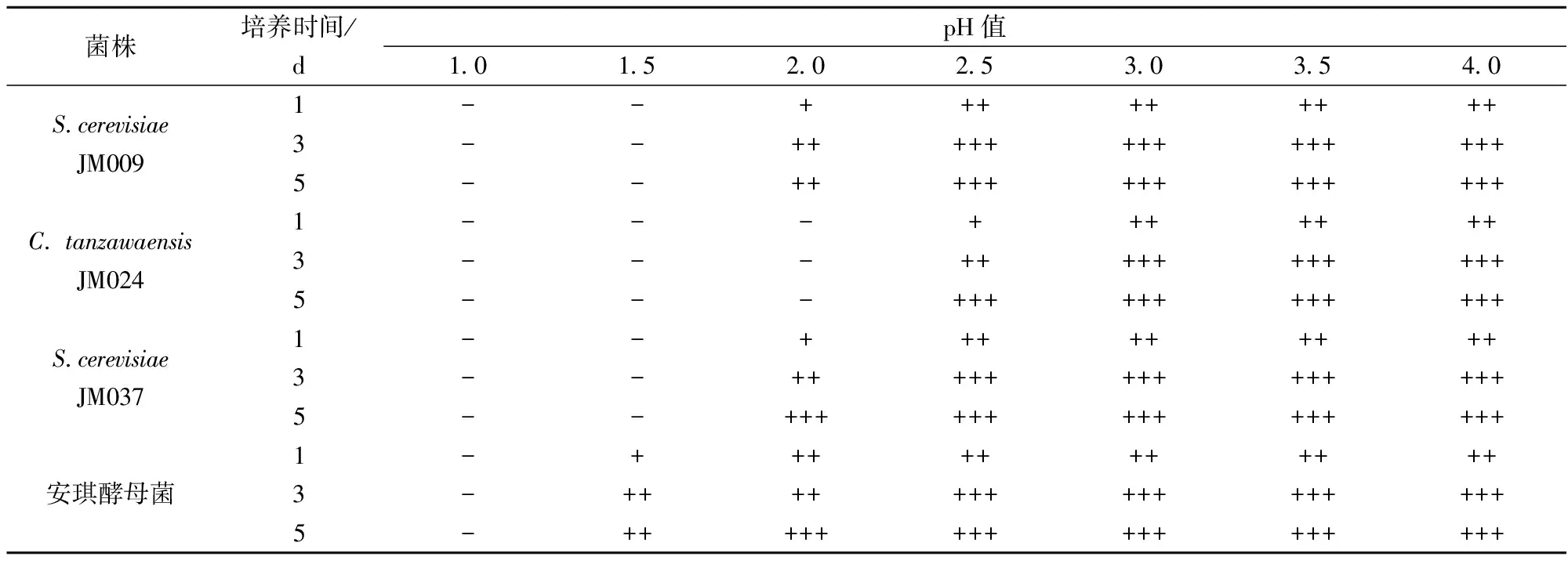
表5 产GABA酵母菌pH耐受性结果
注: +: 杜氏管内满1/4管; ++: 满1/2管; +++: 满管。
2.5 酿酒酵母JM037发酵性能测定
以安琪酵母菌为参考,以菌体生物量、发酵过程的残糖、GABA和乙醇生成量为指标,对酿酒酵母JM037发酵性能进行初步评估。由图4-A可以看出,随着时间的增加,酿酒酵母M037菌体生物量在180 h达到最大,此时发酵液中含有少量的残糖和较高浓度的乙醇,使菌株细胞机能变弱,进而导致细胞自溶,菌株生物量基本保持不变,从图4-B和图4-C残糖消耗速率和乙醇生产速率可以看出,酿酒酵母JM037发酵速率比对照缓慢,对照组在120 h乙醇产量达到最大(48.57 g/L),在156h残糖降至3.15 g/L,而酿酒酵母JM037在168 h乙醇产量达到最大(38.36 g/L),在216 h残糖降至3.06 g/L。另一方面,安琪酵母菌可以产生酒精,但不产生GABA,酿酒酵母JM037在高产GABA(1145 mg/L)的同时,还具有良好的产酒精的能力,见图4-D。因此,酿酒酵母JM037具有酿制富含GABA果酒的潜能。
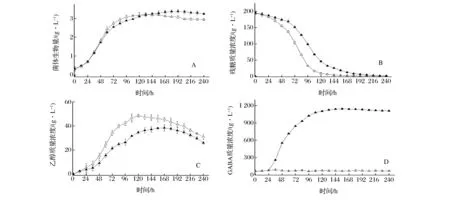
图4 酿酒酵母JM037发酵各项指标Fig.4 The indicators for fermentation of S. cerevisiae JM037
3 结论
从四川泡菜中初筛得到3 株具有产GABA能力的酵母菌。经生理生化和18S rRNA序列分析,菌株JM009和JM037被鉴定为酿酒酵母、菌株JM024被鉴定为C.tanzawaensis。通过HPLC对菌株GABA表达能力进行评估发现,酿酒酵母 JM037产GABA能力最高。进一步对其进行耐受性及发酵性能测试分析,酿酒酵母JM037综合耐受性良好,并在桑葚汁中静置发酵条件下,酿酒酵母JM037既能高产GABA(1 145 mg/L),还具有良好的产酒精(38.36 g/L)的能力。因此酿酒酵母JM037具有酿制富含GABA果酒的潜能。
[1] MARINA D, JOAN Q, MAGDALENA R, et al. Gamma-aminobutyric acid as a bioactive compound in foods: a review[J]. Science Direct, 2014, 10: 407-420.
[2] SUZAN G R, CAROLINE B Q, ELUZA C S, et al. Diphenyl diselenide ameliorates monosodium glutamate induced anxiety-like behavior in rats by modulating hippocampal BDNF-Aktath way and uptake of GABA and serotonin neurotransmitters[J]. Physiology and Behavior, 2016, 155(1): 1-8.
[3] VAN D P, SEUNG W L, SI J P, et al. Production of gamma-aminobutyric acid from glucose by introduction of synthetic scaffolds between isocitrate dehydrogenase, glutamate synthase and glutamate decarboxylase in recombinantEscherichiacoli[J]. Journal of Biotechnology, 2015, 207(10): 52-57.
[4] PETR U, SERGEI K, HEIKO J L, et al. GABA transporters control GABA ergic neurotransmission in the mouse subplate[J]. Neuroscience, 2015, 304: 217-227.
[5] TRACIE A P, ELIZABETH K C, DANIEL C L, et al. Effects of chronic inhibition of GABA synthesis on attention and impulse control[J]. Pharmacology, Biochemistry and Behavior, 2015, 135: 97-104.
[6] KO C, VICTOR L H, TSAI G. Gamma-aminobutyric acid production in black soybean milk byLactobacillusbrevisFPA 3709 and the antidepressant effect of the fermented product on a forced swimming rat model[J]. Process Biochemistry, 2013, 48(4): 559-568.
[7] NEJATI F, RIZZELLO C, DI C R, et al. Manufacture of a functional fermented milk enriched of angiotensin-I converting enzyme (ACE)-inhibitory peptides and gammaamino butyric acid (gaBa)[J]. LWT - Food Science and Technology, 2013, 51(1): 183-189.
[8] KIM J Y, LEE M Y, JI G E, et al. Production of γ-aminobutyric acid in black raspberry juice during fermentation byLactobacillusbrevisGABA100[J]. International Journal of Food Microbiology, 2009, 130(1): 12-16.
[9] ANUSSARA R, DUANGPORN K, WILAWAN C, et al. Enhancement of γ-aminobutyric acid (GABA) in Nham (Thai fermented pork sausage) using starter cultures ofLactobacillusnamurensisNH2 andPediococcuspentosaceusHN8[J]. International Journal of Food Microbiology, 2013, 167(2): 170-176.
[10] 吴岱熹, 吴非. 富含γ-氨基丁酸清酒酒曲的研究[J]. 食品工业科技, 2012, 33(21): 188-191.
[11] 李海星. 短乳杆菌CCTCCM 208054生物转化制备γ——氨基丁酸及其GAD系统关键基因分析[D]. 南昌:南昌大学, 2012.
[12] BARNETT J A, PAYNE R W, YARROW D, et al. Yeasts: Characteristics and Identification, 2nd edn[M]. Guangzhou:Cambridge University Press, 1990.
[13] 李可. 中国浓香型白酒发酵黄水中微生物群落结构及多样性解析[D]. 成都: 西华大学, 2014.
[14] 张术聪. 固定化植物乳杆菌合成γ-氨基丁酸及分离纯化的初步研究[D]. 无锡: 江南大学, 2010.
[15] 何晶龙, 彭超, 王欣, 等. 笃斯越桔中酿酒酵母菌的筛选、鉴定及特性研究[J]. 中国食品学报, 2011, 11(3): 77-88.
[16] TRIGUEROS D E G, FIORESE M L, KROUMOV A D, et al. Medium optimization and kinetics modeling for the fermentation of hydrolyzed cheese whey permeate as a substrate forSaccharomycescerevisiaevar.boulardii[J]. Biochemical Engineering Journal, 2016, 110: 71-83.
[17] 谢文, 丁慧瑛, 章晓氡, 等. 葡萄酒、果酒中乙醇含量的测定方法[J]. 色谱, 2004, 22(5): 561-561.
[18] 李亚莉, 秘鸣, 魏珍珍, 等. 一株产GABA酵母菌的筛选及鉴定[J]. 食品科技, 2013, 38(6): 16-21.
[19] 乌云达来, 王肇悦, 郭雪娜, 等. 产γ-氨基丁酸酵母菌的筛选及菌种鉴定[J]. 内蒙古农业大学学报, 2013, 34(6): 110-114.
[20] 贾春凤, 贾志军, 朱维红,等. 红提葡萄酒发酵酵母菌的分离及耐受性研究[J]. 中国酿造, 2016, 35(6): 76-80.
[21] 高玉妹, 陶能国, 刘跃进, 等. 白酒酒曲中9株酵母菌的分离与特性研究[J]. 食品工业科技, 2010, 31(5): 195-201.
[22] 于洋, 粟春燕, 陈晶瑜. 3株葡萄酒非酿酒酵母的生长与发酵特性[J]. 食品科学, 2015, 36(19): 106-111.
[23] 张大为, 张洁, 洪磊东, 等. 发酵梨酒中产香酵母的分离、鉴定及生物学特性的研究[J]. 食品工业科技, 2013, 34(2): 179-181.
Screening for bioconversion γ-aminobutyric acid-producingSaccharomycescerevisiaeand its application in mulberry wine brewing
ZENG Lin1,2,TAN Xiao1,ZHANG Qing1,2*,YANG Ying,TANG Jie1
1(Provincial Key Laboratory of Food Biotechnology of Sichuan, College of Food and Bioengineering, Xihua University, Chengdu 610039, China)2(Biotechnology Institute of Ancient Brewing, Xihua University, Chengdu 610039, China)3(Food and Drug Institute of Shandong, Shandong, Jinan 250101, China)
In order to obtain yeasts which could produce γ-aminobutyric(GABA) and brew good performance mulberry wine, 3 strains of GABA-producing yeast were screened from Sichuan pickles by paper chromatography. Then, they were identified asS.cerevisiaeandC.tanzawaensisusing the physiological and biochemical tests and the phylogenetic analysis. The performance of GABA-producing yeasts was assessed by high-performance liquid chromatography (HPLC) method. The results showed that theS.cerevisiaeJM037 produced higher yield of GABA, and the content of GABA reached 670 mg/L. ThenS.cerevisiaeJM037 was further analyzed for its tolerance properties and mulberry wine brewing ability. The results showed thatS.cerevisiaeJM037 had good comprehensive tolerance properties and it GABA-producing capacity increased to 1145 mg/L at 144 h. Ethanol production was 38.36 g/L at 168 h, while residual sugar dropped to 3.06 g/L at 216 h. In addition, ethanol synthesis rate and the residual sugar consumption rate ofS.cerevisiaeJM037 was relatively slow compared with angel yeast, butS.cerevisiaeJM037 has a unique capacity to produce GABA. Therefore,S.cerevisiaeJM037 has potential for brewing fruit wine with GABA.
γ- aminobutyric acid;S.cerevisiae; tolerance properties; screening; identify; mulberry wine
硕士研究生(张庆副教授为通讯作者,E-mail:biozhangq@163.com )。
四川省应用基础项目(2016JY0253);教育部春晖计划项目(Z2014060);四川省食品生物技术重点实验室项目(szjj2016-019)
2017-01-03,改回日期:2017-02-07
10.13995/j.cnki.11-1802/ts.201706020
