防止和修复湖泊水库富营养化渔业操纵的探讨
2017-07-24王宇庭叶金云郑荣泉
王宇庭,叶金云,郑荣泉
(1.湖州师范学院浙江省水生生物资源养护与开发技术研究重点实验室,中国水产科学研究院水生动物繁育与营养重点实验室,浙江湖州313000;2.浙江师范大学浙江省野生动物生物技术与保护利用重点实验室,浙江金华321004)
防止和修复湖泊水库富营养化渔业操纵的探讨
王宇庭1,叶金云1,郑荣泉2
(1.湖州师范学院浙江省水生生物资源养护与开发技术研究重点实验室,中国水产科学研究院水生动物繁育与营养重点实验室,浙江湖州313000;2.浙江师范大学浙江省野生动物生物技术与保护利用重点实验室,浙江金华321004)
为防止和修复湖泊水库富营养化,通过渔业操纵实现营养物质的上岸,找到适合所有水体的生态治理方式,本研究中系统归纳了在不同营养类型、不同水体形态和不同营养物质输入节奏水体的渔业操纵对策。结果表明:对于受面源污染较小,营养物质输入分散的贫、中营养水体,应以保护生物多样性的长食物链的渔业形式为主,即实行以肉食性鱼类为主要对象的渔业操纵;随着水体富营养化程度的提高或营养物质输入集中度的提高,逐渐增加杂食性鱼类的比例,其中滤食性鱼类和底层杂食性鱼类的放养比例应根据水体深度和面积进行调整,不应夸大滤食性鱼类在所有形态水体中的除藻效果,尤其在浅而大的湖泊中,底层杂食性鱼类可通过吞食或刮食被湖盆吸附的藻类,实现除藻作用;对于超富营养水体,除了以杂食性鱼类为主的渔业操纵方式外,还需要增加水体的异质性,以提高水体的生物多样性,以及水中营养物质(鱼产量)被浓缩上岸的速度。本研究中首次提出对大而浅的富营养水体进行围栏分割,将大生态系统分割为互相依托的小生态系统景观集合体的生态修复方法,同时还阐述了渔业操纵效果的评价方法。
渔业操纵;富营养化;异质性;生态修复
中国各地因水文、气候和土壤等差异较大,各地水质状况也不同。为充分利用中国有限的淡水资源,需要从保护水质角度出发,优化水生态系统的结构以提高水体的自净能力。富营养化(Eutrophication)是中国水资源面临的严峻问题,尤其大中型湖泊水库的富营养化[1-3],给科研人员带来了严峻挑战,也是70年来水科学领域在世界范围内的难题[4-10]。中国作为湖泊水库(以下简称为湖库)渔业开发力度最大的国家,如何进行更合理的湖库放牧渔业,实现防止和修复湖库的富营养化,即渔业操纵(Fisheries manipulation),已成为中国渔业产业的重要任务。近10年来,中国的湖库渔业一直在该领域不断探索,至今尚无重复的渔业操纵方法大幅减少藻类的范例,虽然武汉东湖有“水华”消失的良好状况,但使用相同的方法——用滤食性鱼类抑制藻类,在其他大型浅水湖泊,如太湖、巢湖和天池并无理想的效果,太湖仍然在人工捞藻。同时,另一种相反的措施——禁止任何渔业行为的水源水库保护措施,也在长三角部分地区实行。因此,有必要对用渔业方式优化水生态结构的方法进行总结和反思,以便为科研、环保和渔业部门提供湖库鱼类调控的全面思路。
1 富营养化的发生
富营养化发生的外部原因是水层营养物质浓度达到一定程度,同时光照、水温适当;内部原因是藻类的主要消费者生物量不足或生物多样性过低,使得部分或全部藻类群落因不能及时消耗,而得以大量滋生。当前,外源营养物质的输入和内源营养物质的释放是导致水层营养盐过高的原因,其中,外源营养物质输入的点源和面源两个方式中,面源污染在中国季风气候区因降雨的不均尤其严重[2]。由此发现,夏季汛期水位一定范围内越高,水位下降后营养盐的密度就越大,并导致水体生产力增加[11]。内源营养物质的释放是由于河床或湖盆经长期吸附水层营养盐而形成的营养盐库,经物理搅动或水质波动(如pH降低),再次释放出来[12]。对中国水体营养盐指标、形态和地理位置与富营养化特征指标的回归分析发现,总磷和水体营养特征与水体高程、水体深度、体积和纬度相关,同时,不同水体表现出对营养物质的输入有不同的缓冲能力[13]。本研究中进一步探讨了水体特征和富营养化发生的关系。
1.1 易发生富营养化水体的水文和生态特征
1.1.1 水文特征与富营养化的关系 两者的关系主要表现在:(1)中国季风气候使得降雨量不均匀,导致集水区内面源营养盐输入总量大且集中,因而集水区内的工农业规模和人口密度,直接影响面源输入营养物质的总量,如人口密度大的发达地区,汛期后的湖泊容易发生富营养化现象[12];(2)降雨量与富营养化的关系密切相关,除与集水区内的人口密度和工农业生产规模有关外,也与该区域的土壤和植被有关,通常降雨量越大,面源污染物越多,富营养化越易发生,但降雨量过大,又会稀释营养盐,降低富营养化发生的机会;(3)作为人工湖泊的水库,尤其是山谷型水库,出水口较低(死库容以上),因而能将营养物质比较丰富的下层水排放出去,从整体上更有效地降低了水体的营养盐浓度,因此,发生富营养化的机会相对天然湖泊要低;(4)水位波动剧烈的水体,底层积累大量营养物质的机会较低,所以突发富营养化的机会也较低;(5)入水口与出水口的距离与水体周长的比值同样影响营养物质的积累,也影响富营养化的发生。
1.1.2 生态特征与富营养化的关系 两者的关系主要表现在:(1)当浮游动物的生物多样性较高但生物量较低时,浮游动物增殖速度较慢,若外界环境条件变得有利于植物的生长,浮游植物会迅速滋生,超过浮游动物的增殖速度,如春、秋季水的上下对流使内源营养物质大量释放到水层,藻类便持续增加直到营养盐缺乏或形成自荫现象,之后被规模不断增大的浮游动物摄食而藻类规模开始下降,一般此类情况季节性发生在分层湖库中;(2)初级消费者多样性较低,水体缺乏消耗水中各种藻类的动物群落,使得个别藻类大量繁殖,尤其是小型藻类,例如滤食性或小型杂食性鱼类较多的水体,因这些鱼类对大型藻类和浮游动物的摄食,使得小型藻类在缺少捕食者和竞争者的情况下大量滋生;(3)当上述富营养化发生后,藻类优势种群密度过大且发生老化,同时产生藻毒素,如微囊藻的微囊藻毒素,抑制竞争者[14]和部分捕食者[15-16],使水体的富营养化持续时间更长。
1.2 易发生富营养化水体的形态特征
水体的形态特征决定水环境的空间异质性(Spatial heterogeneity),空间异质性决定水体现时的生物多样性[17],同时,在捕食者和被捕食者的关系中,空间异质性决定着被捕食者庇护所的密度,从而影响捕食者和被捕食者关系的紧张程度。由此,本研究中推测水体的空间异质性影响着滤食性鱼类对浮游动物的压力,影响着水体浮游动物与浮游植物生物量的比值(Bz∶Bp)。Bz∶Bp可作为评价水体对富营养化缓冲能力的重要指标[18]。由于湖盆粗糙度难以测定,本研究中用湖盆表面积与水面面积的比值表示水体的异质性,进一步简化用湖库周长(P)×水深(Z)/水面面积(S)表示,当样本水体形状一致时,再简化为根据李德尚等[19]的调查数据和何志辉[20]引用的数据,分别计算出二者的关系如图1所示,即:
(1)深度大、面积小的水体,浮游动、植物生物量比值在较小范围内波动,缓冲性较差,当营养盐浓度增加,容易发生富营养化。
(2)随着水深与水面面积比值的增加,浮游动、植物生物量比值的波动范围迅速增大并达到最高值,即在该数值附近的湖泊水库中,营养盐过量也不易导致富营养化,因为浮游动物的现存量可增加到相对较高的比值,从而抑制浮游植物的滋生。
(3)李德尚等[19]的研究中,山东水库异质性数据在0.017时,浮游动、植物比值最大(0.9),何志辉[20]的研究中,湖泊异质性数据在0.7时,浮游动、植物比值最大(1.8)。
(4)水体深度和水面面积的比值大于该数值后,浮游动、植物的比值会缓慢下降,即水体对富营养化的缓冲能力会缓慢下降。这表明,水体的形态影响着浮游动、植物的比例,因而影响着浮游动物对浮游植物的抑制作用,当水体浊度较低且日照合适时,水体易发生“水华”。
2 渔业操纵对策
从物质流动的角度实现水质保护就是最大程度地以鱼类等水生生物为载体,从水中提取出磷等植物营养物质,减少其在水层中的负荷和在水底的淤积,提高水的纯净程度和延长湖库的寿命。其中,以鱼类为载体的渔业操纵行为是最经济的措施。为实现此目标,使得营养物质的输入等于或小于营养物质的输出,需要相应的湖库生态系统具备完备的食物网和高效的牧食链(Grazing food chain)。相对于海洋生态系统几十亿年的历史,湖库食物网组织的历史较短,且因水体规模小而易受到环境突变的干扰,所以需要人为干预,以构建和维护相应的牧食链和食物网。本研究中,主要探讨防止、抑制和修复富营养化的3种渔业操纵方式。
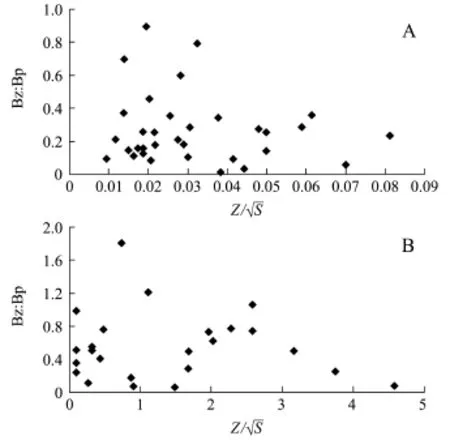
图1 水库湖泊形态(Z/S)与浮游动物、植物生物量比值(Bz:Bp)的关系Fig.1 Relationship between morphologic factor and biomass ratio of zooplankton to phytoplankton Bz:Bp in reservoirs and lakes
2.1 以肉食性鱼类为主的渔业操纵
此类水体中,磷的输入量、负载量和水体初级生产力均较低,决定了鱼产量也较低,因而不论以哪种鱼类为主,只要牧食链各环节没有障碍,现存量合适,产量大体一致,与牧食链的长度无关,因为在牧食链各环节没有障碍的情况下,其局部反复运行实现的营养物质的运输和浓缩到食物链末端的速度等于或超过营养物质输入的速度。因此,这类水体应该以高附加值的肉食性和土著鱼类的增殖为主,既抑制水体的富营养化,又保护生物多样性,同时提高经济效益(因肉食性鱼类的经济价值一般比杂食性鱼类高)。
该方法与流行于欧美的以修复水体富营养化为目的的生物操纵(Biomanipulation)[5-8]方法类似,不同的是生物操纵方法注重抑制杂食性鱼类(Omnivores),而不注重人为收获肉食性鱼类(Piscivores),使肉食性鱼类年龄组成不合理,生产力下降,水生态系统食物链的基本功能——传递水中营养物质并使之上岸,得不到发挥。统计显示,欧美生物操纵方法仅使外源营养物质得到有效控制,对面积小于0.4 km2的大部分浅水体(平均为0.18 km2)有效[7]。本研究中所述渔业操纵更注重食物链各环节中最大生产力的实现,以实现食物链的畅通和高效。
在新合拢的水库中,由于长牧食链和对应食物网形成需要的时间较长,加上肉食性鱼类容易被垂钓,资源量容易遭到破坏,管理部门更倾向投放食物链短的鲢、鳙鱼类,或放流银鱼等小型经济鱼类。但鲢、鳙切短食物链,使鱼类和浮游生物的多样性降低[21-23];小型鱼类产量会因自身的密度调节和水文情况的变化而大幅波动[24]。因此,正确的做法是:对食物链薄弱环节的生物群落进行必要的增殖活动,以实现其最大产量;其次是适当投放底层杂食性鱼类,如鲤、鲫、河蟹等,以搅动底泥,抑制沉积,释放底泥中积累的营养物质。这方面许多水库均有过失误,如辽宁省碧流河水库在大规模放养鲢、鳙后,香鱼和斑鳜明显减少。也有些小型水库为了提高鲢、鳙产量,而大量施肥,导致一些对水质要求高的土著经济鱼类难以恢复。
适用于该方法的水体,一般是贫、中营养型的水库和湖泊,如位于长江、黄河、辽河、鸭绿江、图们江和松花江的上游、支流,以及珠江流域的大部分河段和沿海丘陵的短河流域[1-3],除珠江下游外的水体,一般属山谷型水库或高原湖泊,全年大部分时期水质清澈,只有在枯水期有大量藻类发生,同时透明度下降。水库的透明度在不同断面的差异,主要是由泥沙悬浮量决定。
2.2 以杂食性鱼类为主的渔业操纵
此类水体主要位于长江、黄河和辽河的中下游,包括中游山谷型水库、下游平原型水库和各河段的湖泊[1-3]。这类水体水质通常季节变化明显,一年中至少有一段时间发生“水华”。要实现营养物质的最少积累,仅靠肉食性鱼类是不够的,因为营养物质的输入速度已经超过了由长牧食链的运输和浓缩的速度,需要增加食物链短的滤食性和底层杂食性鱼类,以抑制富营养化的趋势,稳定水质,使之即无“水华”也无异常清澈的时期。其中,营养物质输入速度过快,主要表现在两个方面:一方面,营养物质年际输入的总量过大;另一方面,营养物质的输入过于集中,在一段时间内超过了食物链传递的速度,造成营养物质积累。
中国的非经典生物操纵(Non-traditional biomanipulation)理论和实践与此类似[25],只是目前该理论的实际应用中,仅重视了鲢、鳙对藻类的直接摄食作用,而忽视了其他杂食性鱼类通过吞食或刮食被湖盆吸附藻类的食藻作用。本研究中强调所有杂食性鱼类的作用,包括中上层滤食性鱼类和底层杂食性鱼类,尤其后者在面积大、水浅的湖泊中作用更大。因此,在确定富营养化水体中食藻鱼鲢、鳙的比例时,应结合水体的形态、湖盆土质和气候:(1)水体形态方面,应根据湖盆表面积与水体积的比例,确定底层杂食性鱼类与滤食性鱼类的放养比例,即表面积比例越大,浅、小、岸线发育学指数高的水体,应放养更高比例的底层鱼类,尤其夏季没有稳定温跃层的持续循环湖库,但具体数值尚缺乏定量的研究成果,如笔者在黄河三角洲平原型湖泊(水库)的渔业操纵经验是,滤食性鱼类白鲢和底层杂食性鱼类鲤、鲮和梭鱼等作为主要放养鱼类,其中底层鱼类所占比例均为牧养鱼总量的40%~60%,白鲢为30%~50%,花鲢仅为10%;(2)湖盆土质影响悬浮物颗粒大小和丰度,从而影响对小型藻类的吸附作用,如长江水稻土、黄河土母质和东北黑钙土均有不同的吸附藻类能力,因而底层杂食性鱼类的放养比例在相同形态的水体中也应不同;(3)气候特点决定水体的风成环流和垂直对流的强度,从而影响底质颗粒的悬浮和运动,以及对藻类的吸附作用,因而温度日差较大、风大的地区,底层鱼类应投放较高比例。
2.3 增加异质性为主杂食性鱼类为辅的渔业操纵
在富营养化程度严重的浅湖泊,上述渔业操纵实现的鱼产品携带营养物质上岸的速度,已经不足以实现所有过量营养物质上岸的目标。这类水体目前主要分布于长江、淮河流域,属平原型湖泊或水库,尤其是浅而大的水体,水平环流和垂直对流比较充分,水环境的异质性较低,浮游动物更易受到滤食性鱼类的压制,不能发挥抑制小型浮游植物的作用,使得牧食链的运行不够通畅,微型藻类大量滋生,如水深为2 m、面积为2300 km2的太湖。滇池水深为4.4 m、面积为330 km2,但因海拔高,昼夜温差大,密度环流较强,不能形成稳定温跃层,浮游动物同样容易被压制,而使浮游植物大量滋生。为了发挥杂食性鱼类短食物链的优势,又发挥浮游动物抑制微型藻类的作用,需减少滤食性鱼类尤其是鳙的比例,增加底层杂食性鱼类的比例,同时增加水体的异质性,例如使用生态庇护所对浮游动物的现存量进行保护,以提高其生产力,抑制微型藻类[26]。目前,中国集约化养殖中的综合技术已经有此理论的运用[27]。增加水体异质性的方法包括人工鱼礁、围栏、围隔等,其中在不同的围栏内投放不同鱼类,并成为相互协调的类似景观生态系统的斑块和廊道的功能体系的方法,应该最为有效。目前,该方法国内外尚属空白,国内有关部门或专家学者尚未给予关注和支持。
3 生物操纵效果的评价
评价生物操纵的效果,首先要检验环境化学和环境生物学指标是否好转,以及环境生物是否受到了合适的影响,其次是评价关键种——大型动植物群落结构是否稳定而合理,从而确定生态系统是否稳定,环境指标的变化是一种趋势还是暂时状况。环境指标在所有相关研究中均可观察到,但大型动植物的指标却经常被忽视,即在研究不同水草对环境影响时,忽略不同水草在不同生长阶段的差异;研究鱼类对环境影响时,仅仅关注了藻类等环境指标的变化,而没有关注鱼类本身是否生长正常,鱼类群体结构是否可以持续稳定[28]。
3.1 环境化学和环境生物学指标
实践中这是用以判断环境质量的直接指标,通过监测其时空变化,结合大型动植物(鱼类和水草)的结构和生长周期,可以预报水环境的发展趋势。主要化学指标包括:水层总磷、矿化度(或氯度、电导率)、pH和溶解氧,以及底泥总磷、底泥生物有效磷等。这些指标包含水体营养状态和发展趋势的信息,如矿化度较高意味着水在地表的历程较长,其营养物质的积累较大程度是因不断浓缩造成;pH的持续弱碱性意味着有机物产生的速度持续超过分解的速度等。
环境生物学的指标包括:叶绿素含量、浮游植物现存量及其群落结构、浮游动物现存量及其群落结构、底栖动物现存量及其群落结构,以及生物多样性指数和空间异质性指数。现存量特别是叶绿素a含量和浮游植物的现存量可以用来评价水质状况,而多样性指数和空间异质性指数可以作为评价鱼类下行影响的敏感指标——鱼类的下行影响首先会使低等生物群落的多样性指数和空间异质性降低到一定水平[17]。持续一段时间直到鱼类的生长速度减慢,再人为减少鱼类的现存量,藻类个体增大、丰度下降,营养盐(水层总磷和底泥总磷)含量下降,水质得到改善。
3.2 鱼类组成指标
作为水生态系统中的大型动物,鱼类通常是控制和影响整个水生态系统的关键种,因此,要关注水中现存鱼类的生长状况和群体结构,以确保生物操纵过程中鱼类的正常生长,且正在实现最大鱼产量。因为如果鱼类的生产量未达到最大,或小于营养物质沉积的速度,富营养化将不可避免。分析评价时,首先检测鱼类生长是否正常,其次是评估放养捕捞维持的鱼类群体结构和捕获量是否合理。通过测定其体长组成,结合个别样本的鳞片、鳍条或耳石的年轮鉴定,可以确定该群体的年龄结构和年生长速率,再结合肥满度指标,可以断定该鱼的生长速度、群体生产量和放养捕捞量是否达到最大[29]。
判断鱼类的群体结构和捕获量是否合理,相对比较复杂。首先通过饵料生物的丰度和最大个体大小的变化判断饵料生物的丰欠程度,最佳状态是饵料生物现存量没变,但个体大小略有下降;其次是通过竞争者的丰度变化,如野生小型杂食性鱼类丰度提高,意味着主要投放的杂食性鱼类密度不足;通过野生大型肉食性鱼类丰度的变化,如果其丰度增加,意味着主要经济鱼类放养不足或放养个体规格偏小。
生产实践中通常情况下是鱼类现存量不足,导致生产量不足,进而营养物质积累,富营养化速度加速。鱼类现存量不足的原因:一方面,捕捞过度或滥捕无序,这从太湖的渔业状况中可见一斑[30-31];另一方面,管理部门未随湖库营养状态的提高增加现存量以提高产量,即增大放养量或提高捕捞规格,如辽宁大伙房水库,富营养化程度提高[32],但鱼产量反而降低。
4 结语
受人类行为的影响,内陆水域水层营养物质的增加成为必然,因而强化水体生态功能和结构成为维持水体功能的必然需要。国内外的非经典和经典生物操纵理论,强调了生物要素间的关系,本研究中提出的渔业操纵理论更关注营养物质沿食物链流动和上岸的畅通性,提出了根据水体的营养状态、营养物质输入节奏和水体形态确定最经济的富营养化防止和修复的渔业操纵方案。
[1] 张萍,高丽娜,孙翀.中国主要河湖水生态综合评价[J].水利学报,2016,47(1):94-100.
[2] 陈小锋,揣小明,杨柳燕.中国典型湖区湖泊富营养化现状、历史演变趋势及成因分析[J].生态与农村环境学报,2014,30 (4):438-443.
[3] 成小英,李世杰.长江中下游典型湖泊富营养化演变过程及其特征分析[J].科学通报,2006,51(7):848-855.
[4] 刘其根,张真.富营养化湖泊中的鲢、鳙控藻问题:争议与共识[J].湖泊科学,2016,28(3):463-475.
[5] Jørgensen S E,de Bernardi R.The use of structural dynamic models to explain successes and failures of biomanipulation[J].Hydrobiologia,1998,379(1-3):147-158.
[6] Drenner R W,Hambright K D.Biomanipulation of fish assemblages as a lake restoration technique[J].Archiv Fur Hydrobiologie, 1999,146(2):129-165.
[7] Jeppesen E,Meerhoff M,Jacobsen B A,et al.Restoration of shallow lakes by nutrient control and biomanipulation-the successful strategy varies with lake size and climate[J].Hydrobiologia,2007,581 (1):269-285.
[8] Bernes C,Carpenter S R,Gårdmark A,et al.What is the influence of a reduction of planktivorous and benthivorous fish on water quality in temperate eutrophic lakes?A systematic review[J].Environmental Evidence,2015,4:7.
[9] VanMiddlesworth T D,McClelland N N,Sass G G,et al.Fish community succession and biomanipulation to control two common aquatic ecosystem stressors during a large-scale floodplain lake restoration[J].Hydrobiologia,2016,769(1):1-16.
[10] 秦伯强,高光,朱广伟,等.湖泊富营养化及其生态系统响应[J].科学通报,2013,58(10):855-864.
[11] 史为良,夏德昌,董双林,等.大伙房和柴河两水库的环境因子同鲢、鳙生长和产量变动的关系[J].海洋与湖沼,1994,25 (1):77-86.
[12] 史静,于秀芳,夏运生,等.影响富营养化湖泊底泥氮、磷释放的因素[J].水土保持通报,2016,36(3):241-244.
[13] Liu Wenzhi,Zhang Quanfa,Liu Guihua.Lake eutrophication associated with geographic location,lake morphology and climate in China[J].Hydrobiologia,2010,644(1):289-299.
[14] Bittencourt-Oliveira M D C,Chia M A,Oliveira H S B D,et al. Allelopathic interactions between microcystin-producing and non-microcystin-producing cyanobacteria and green microalgae:implications for microcystins production[J].Journal of Applied Phycology,2015,27(1):275-284.
[15] Chen Feizhou,Dai Xi,Shu Tingting,et al.Microcystins derived from lysing Microcystis cells do not cause negative effects on crustacean zooplankton in Lake Taihu,China[J].Aquatic Ecology, 2013,47(4):379-387.
[16] Zhang Junqian,Xie Zhicai,Wang Zhi.Oxidative stress responses and toxin accumulation in the freshwater snail Radix swinhoei (Gastropoda,Pulmonata)exposed to microcystin-LR[J].Environmental Science and Pollution Research,2016,23(2):1353-1361.
[17] Brock W,Kinzig A,Perrings C.Modeling the economics of biodiversity and environmental heterogeneity[J].Environmental and Resource Economics,2010,46(1):43-58.
[18] Havens K E,Beaver J R.Zooplankton to phytoplankton biomass ratios in shallow Florida lakes:an evaluation of seasonality and hypotheses about factors controlling variability[J].Hydrobiologia,2013,703(1):177-187.
[19] 李德尚,焦念志,周春生,等.山东省大中型水库鱼产力的综合评估[J].水产学报,1993,17(2):95-104.
[20] 何志辉.中国湖泊和水库的营养分类[J].大连水产学院学报,1987(1):1-10.
[21] Byers J E.Impact of non-indigenous species on natives enhanced by anthropogenic alteration of selection regimes[J].Oikos,2002, 97(3):449-458.
[22] Domaizon I,Devaux J.Experimental study of the impacts of silver carp on plankton communities of eutrophic Villerest reservoir (France)[J].Aquatic Ecology,1999,33(2):193-204.
[23] 王宇庭,孙建,阎红山.鱼类对生态系统下行影响敏感指标初探[J].莱阳农学院学报,2002,19(2):139-144.
[24] 黄真理,常剑波.银鱼的产量能预报吗[J].生态学报,2001, 21(1):86-93.
[25] Xie Ping,Liu Jiankang.Practical success of biomanipulation using filter-feeding fish to control cyanobacteria blooms:a synthesis of decades of research and application in a subtropical hypereutrophic lake[J].The Scientific World,2001,1:337-356.
[26] 王宇庭.生态庇护所式富营养化水的处理方法:中国, 201610429504.4[P].2016-06-15.
[27] 董双林.中国综合水产养殖的生态学基础[M].北京:科学出版社,2015.
[28] 唐汇娟,谢平.围隔中不同密度鲢对浮游植物的影响[J].华中农业大学学报,2006,25(3):277-280.
[29] 王宇庭,赵文,张久国,等.根据渔获物评价鲢、鳙放养比例的方法及应用[J].大连水产学院学报,2001,16(2):99-105.
[30] 刘恩生,刘正文,鲍传和.太湖鲫鱼数量变化的规律及与环境间关系的分析[J].湖泊科学,2007,19(3):345-350.
[31] 何俊,谷孝鸿,王小林,等.太湖鱼类放流增殖的有效数量和合理结构[J].湖泊科学,2012,24(1):104-110.
[32] 焉鸿启,赵文,郭凯,等.辽宁省6座水源水库富营养化状况的分析与评价[J].大连海洋大学学报,2016,31(2):180-184.
Insight into fisheries manipulations to inhibit and restore eutrophications in lakes and reservoirs
WANG Yu-ting1,YE Jin-yun1,ZHENG Rong-quan2
(1.Zhejiang Provincial Key Laboratory of Aquatic Resources Conservation and Development,Key Laboratory of Aquatic Animal Genetic Breeding and Nutrition,Chinese Academy of Fishery Sciences,Huzhou University,Huzhou 313000,China;2.Zhejiang Provincial Key Laboratory of Biotechnology and Protective Utilization of Wild Animals,Zhejiang Normal University,Jinhua 321004,China)
The fisheries manipulation strategies were surveyed in lakes and reserviors with various trophic types, basin morphology,and nutrient input rhythm to inhibit and restore eutrophications in lakes and reservoirs.The statistic data collected from some reservoirs and lakes indicate that long food chain piscivorous fish and indigenous fish should be protected to increase the biodiversties in oligotrophic waters with less non-point pollution and nutrient input.Omnivores are stocked into the waters as eutrophication become more and more serious,and ratio of filtering feeder to benthivorous fish should be controlled mainly depending on water depth and area,instead of exaggeration of the algae removal effect by filter feeding fishes in all waters.Especially,benthivorous fish should be stocked more to remove algae in shallow lakes by swallowing and scraping the algae that are adhered by lake basins.In ultra eutrophic lakes,the restoration must be carried out by omnivorous fish together with increase of the water heterogeneity such as implanting refuges for zooplankton in order to improve biodiversity and fish production,or increase in the rate of nutrients being concentrated and unloaded from the waters.It is first suggested that fence segmentation be conducted in shallow and large eutrophic waters so that a large ecosystem is divided into small ecosystem landscape complexes that depend on each other.
fisheries manipulation;eutrophication;heterogeneity;ecological restoration
Q954.4
A
10.16535/j.cnki.dlhyxb.2017.04.013
2095-1388(2017)04-0451-06
2016-12-02
浙江省野生动物生物技术与保护利用重点实验室开放项目(20161106);浙江省水生生物资源养护与开发技术研究重点实验室项目(2013E10014);浙江省重点研发计划项目(2015C03018)
王宇庭(1966—),男,副教授。E-mail:wang.yt@hotmail.com
郑荣泉(1969—),男,博士,教授。E-mail:zhengrq@zjnu.cn
