烟草病程相关蛋白NtPR10基因克隆与表达分析
2017-07-05张增林蒋彩虹常爱霞杨爱国罗成刚王绍美王元英
张 玉,张增林,蒋彩虹,常爱霞,杨爱国,罗成刚,王绍美,王元英*
(1.中国农业科学院烟草研究所,青岛 266101;2.中国农业科学院研究生院,北京 100081)
烟草病程相关蛋白NtPR10基因克隆与表达分析
张 玉1,2,张增林1,蒋彩虹1,常爱霞1,杨爱国1,罗成刚1,王绍美1,王元英1*
(1.中国农业科学院烟草研究所,青岛 266101;2.中国农业科学院研究生院,北京 100081)
为鉴定烟草病程相关蛋白PR10的生物学功能,从普通烟草G28中克隆得到了NtPR10基因,基因全长483 bp,编码160个氨基酸。基因结构分析表明,该基因编码的蛋白包含病程相关蛋白家族Bet_v_I保守域,具有与核酸酶活性相关的“P-Loop”结构。利用TMHMM、SignalP、PrositeScan等软件分析发现,NtPR10不含跨膜区,无信号肽,有胞内定位特征。采用实时荧光定量PCR分别分析了TMV诱导下该基因在抗、感品种枯斑三生和G28中的表达模式。结果表明,在TMV诱导下,NtPR10基因在感病品种G28中显著上调表达;而在抗病品种枯斑三生中,TMV侵染后6 h,NtPR10基因显著上调表达,第12小时至第8天显著下调表达,而后至第16天逐渐升高至侵染前水平。综合以上结果,NtPR10应答了TMV侵染过程,并且在抗、感品种中整体呈现出相反的表达趋势,暗示了NtPR10在TMV侵染过程中具有重要功能,为深入分析NtPR10的抗病生物学功能奠定了基础。
烟草;病程相关蛋白;PR10;基因克隆;表达
病程相关蛋白(Pathogenesis-Related Proteins, PRP)是植物体内的一类蛋白质,受病原体或其他外界因子的胁迫而诱导表达,在植物抵御疾病、响应外界压力以及适应不良环境方面发挥着重要作用[1-2]。对于病程相关蛋白的深入研究,不仅能揭示植物抗病、抗逆反应的分子机制,还能为植物抗病分子育种以及病害防治奠定理论和物质基础。
根据病程相关蛋白家族成员的氨基酸组成、结构及生物学功能等特点,将其分为17类,即PR1-PR17。其中,病程相关蛋白10(PR10)包括较大的一个基因家族,其编码的蛋白分子量在15~19 kD,等电点偏酸性,无信号肽序列、属胞内蛋白(intracellular pathogenesis-related proteins,IPR)[3]。目前,在辣椒[4]、花生[5]、水稻[6]、刺茄[7]和小麦[8]等20多种植物中,已有PR10基因相关序列和PR10蛋白相关功能的报道。基因序列分析表明,大部分PR10基因开放读码框编码的氨基酸中都具有一个高度保守的“P-Loop”结构域(GXGGXG)[9]。“P-Loop”是一类广泛存在于磷酸化激酶和核酸结合蛋白的结构域,该区域的磷酸化可能与其核酸酶活性相关[10-11]。PARK等[4]研究证实,辣椒PR10蛋白(CaPR10)比非磷酸化的CaPR10具有更高的核酸酶活性,并发现这种核酸酶活性在抗病毒途径中能降解病毒RNA,且可以抑制辣椒疫霉菌(Phytophthora capsici)的菌丝生长;HE[12]等发现,从葡萄中分离的PR10蛋白(VpPR10.2)同时具有DNA酶活性和RNA酶活性;CHADHA等[5]从花生中克隆的AhPR10基因原核表达的融合蛋白具有抗尖孢镰刀菌(F. oxysporum)和立枯丝核菌(R. solani)活性;WU等[13]研究表明,PR10蛋白超表达可增强水稻对生物胁迫和非生物胁迫的抗性。因此,近年来因其具有的体外核酸酶活性[14-16]和抗菌活性[4,7,17-19],PR10蛋白逐渐成为了研究热点。另外,PR10蛋白还直接参与植物的防卫反应,除了与植物的生物及非生物胁迫密切相关,还在植物的生长发育、次级代谢等过程中起着重要作用。
烟草作为植物病理学研究的模式植物,迄今未见关于PR10蛋白基因及功能的研究报道。本研究首次克隆得到了烟草病程相关蛋白NtPR10基因序列,对其基因结构及编码蛋白特征进行了生物信息学分析,并在TMV诱导下,分析了NtPR10基因在抗、感烟草品种中的表达差异,为深入研究植物PR10蛋白功能及参与的抗病相关分子机理奠定了基础。
1 材料与方法
1.1材料
供试烤烟品种G28为感TMV品种,枯斑三生(SumsunNN)含有抗TMV基因N,为抗TMV品种,均由中国烟草种质资源平台提供;烟草花叶病菌为普通小种(TMV-C),由中国农业科学院烟草研究所植物保护研究中心提供。在烟苗长至六叶期时利用摩擦法接种TMV病毒,每个品种接种20株,每株接种第3、第4片叶,以无菌水接种作为对照,接种后0 h、6 h、12 h、1 d、2 d、4 d、8 d、16 d取第5、第6片叶,用液氮速冻后于-80 ℃保存备用。同时,在接种后2 d和16 d参照烟草病虫害分级及调查方法(GB/T23222—2008)调查TMV发病情况,并按照下列公式计算病情指数:
病情指数(disease index)=100×∑(各级病叶数×各级代表值)/(调查总叶数×最高级代表值)。
1.2方法
1.2.1 样品总RNA提取与cDNA合成 采用RNApure Plant Kit试剂盒提取叶片总RNA,SuperRT cDNA第一链合成试剂盒合成cDNA,用于NtPR10基因的RT-PCR克隆和荧光定量PCR实验。两个试剂盒均购自北京康为世纪生物科技有限公司。
1.2.2NtPR10基因的电子克隆 以茄科作物辣椒的CaPR10基因mRNA序列(Accession No.:AF244121)为“种子”序列,在从NCBI下载的烟草EST数据库中进行Blastn比对,利用SeqMan软件进行EST的拼接和校对,利用NCBI站点的ORF finder程序进一步筛选。
1.2.3NtPR10基因的PCR扩增与克隆 以电子克隆得到的序列为基础,利用Premier 5.0 软件设计特异引物,用于扩增NtPR10基因开放阅读框(Open Reading Frame,ORF)。(FP:5'-ATGGGTGTCACAA CCTATACTC-3';RP:5'-TTAGGCATAGACGGTAG AATTGG-3')。以TMV接种后2 d的的感病品种G28烟草叶片cDNA 为模板,进行PCR扩增,回收目的条带并克隆、测序。PCR反应程序为:95 ℃4 min;95 ℃ 30 s,57 ℃ 30 s,72 ℃ 40 s,35个循环;72 ℃延伸10 min;4 ℃保温。PCR产物回收后连接至pMD®18-T Simple Vector载体(购自宝生物工程生物公司),由上海生工生物工程有限公司进行测序。
1.2.4NtPR10基因序列及编码蛋白序列分析 利用生物信息学网站或软件对NtPR10基因序列及编码蛋白序列进行分析。采用NCBI的ORF finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html)预测开放阅读框;用Blastp进行蛋白序列保守结构域分析;使用ExPAsy网站的Compute pI/MW tool(http:// www.expasy.org/tools/pi_tool.html)计算蛋白质的等电点及分子量;利用TargetP(http://www. cbs.dtu.dk/ services/SignalP/)预测蛋白质信号肽;利用TMpred(http://www.ch.embnet.org/software/TMPRED_for m.html)进行氨基酸跨膜预测;利用PrositeScan(http://npsa-pbil.ibcp.fr/cgi-bin/pattern_prosite.pl)分析蛋白motifs。利用MEGA 6.0软件中的邻接法构建系统进化树[20]。
1.2.5 荧光定量PCR试验 根据NtPR10序列设计荧光定量PCR引物(F:CCTATACTCATGAGGCAT CAACCA;R:AACACCACCATCACCTTCGAC AG),选择烟草肌动蛋白基因(Actin gene,Accession NO.U60495)作为内参基因(F:GCAGCATGAAG ATTAAGGTTGTTG;R:GTGCTAAGGGATGCGA GGAT)。应用Applied Biosystems公司的StepOne PLUS荧光定量PCR仪,以8个取样时间点的cDNA为模板,进行PCR扩增。PCR反应体系见表1。反应程序为:95 ℃ 1 min;95 ℃ 10 s,56 ℃ 20 s, 72 ℃ 40 s,40个循环。反应结束后分析荧光值变化曲线和融解曲线。用Prizm4统计分析软件进行数据统计分析。

表1荧光定量PCR反应体系Table 1 Fluorescence quantitative PCR reaction system
2 结 果
2.1接种TMV后试验材料感病情况
分别于接种TMV后2 d和16 d调查感病品种G28和抗病品种枯斑三生的发病情况,并计算病情指数(图1)。接种后2 d发现枯斑三生的接种叶片均表现出了过敏性坏死反应,出现了坏死斑点。接种16 d后G28接种叶片和大部分非接种叶片均表现出了花叶症状,病情指数为42.22,而枯斑三生除接种叶片残留部分枯死斑外,没有出现花叶症状,表现出了较好的抗病性。

图1接种后16 d G28和枯斑三生病情指数Fig. 1 The disease index of G28 andSumsunNN 16 days postinoculation
2.2NtPR10基因克隆
以辣椒序列为种子序列,以烟草EST数据库为基础,利用Blastn和SeqMan进行序列比对和拼接,得到一条长度为864 bp的序列,利用NCBI网站的ORF finder程序预测,这条序列含有一个长度为483 bp,编码160个氨基酸的完整的开放阅读框(图2),与辣椒CaPR10开放阅读框一致性超过了70%,将这条序列命名为NtPR10。以NtPR10开放阅读框序列设计特异引物,以感病品种G28接种2 d的样品cDNA为模板,经PCR扩增,得到一条与NtPR10电子克隆序列ORF完全一致的核苷酸序列(图3)。目前,该基因已提交到GenBank,注册号为JQ041907。

图2NtPR10序列及其编码的蛋白序列Fig. 2 The sequence ofNtPR10and its encoded protein

图3NtPR10基因PCR扩增Fig. 3 The PCR amplification ofNtPR10gene
2.3NtPR10基因序列及其编码蛋白特征分析
NtPR10基因序列预测编码蛋白经ComputepI/MW软件预测,其等电点为5.56,分子量为17.77 kDa。InterProScan结构域分析表明该编码蛋白2~155位氨基酸为病程相关蛋白Bet_v_I家族保守域,该结构域对外来生物入侵具有防御功能;TMHMM-2.0预测该蛋白不含跨膜区;SignalP 4.0预测该蛋白无信号肽,不属于分泌蛋白;PrositeScan分析将该蛋白定位在胞内。
将烟草(Nicotiana tabacum)NtPR10编码的蛋白序列与辣椒(Capsicum annuum)、马铃薯(Solanum tuberosum,Accession Number:P17642)、番茄(Solanum lycopersicum,Accession Number: NM_001247423)的PR10蛋白序列进行同源性分析,发现其序列较为保守(图4)。其中,烟草与番茄的序列一致性最高,达到了80%,与马铃薯的一致性为74.38%,与辣椒的一致性为65%。进一步分析可知,烟草和辣椒的蛋白序列具有与核酸酶活性相关的“P-Loop”结构(GXGGXG),而马铃薯和番茄PR10蛋白则不具有。已有的研究证明,辣椒PR10蛋白具有体外核酸酶活性[4],而马铃薯PR10蛋白没有[21],由此推测烟草PR10蛋白可能具有核酸酶活性。
2.4系统进化分析

图4 PR10蛋白同源性分析Fig. 4 The homologous Analysis of PR10 proteins
从NCBI数据库中选取了B. pendula(CAA54696)、C. sativa(CAD10374)、P. persica(ABB78006)、M. domestica(CAK93672)、F. sylvatica(CAA10235)、V. vinifera(XP_002274108)、D. glomerata(CAD33532)、P. dulcis(ACE80949)、M. truncatula(XP_003609710)、C. annuum(AAX20045)、S. tuberosum(P17642)、S. virginianum(AAU00066)、S. lycopersicum(NP_001234122)、N. tabacum(JQ041907)、F. chiloensis(ADN05762)、P. domestica(ADN05762)、O. sativa(XP_015631105)、T. aestivum(ACG68733)等18个来自不同植物PR10基因编码的氨基酸序列,用邻接法(neighbor-joining)建立了一个无根的分子系统进化树(图5)。系统进化分析将18个不同植物的PR10蛋白划分为4组,辣椒(C. annuum)、黄果茄(S. virginianum)、马铃薯(S. tuberosum)、番茄(S. lycopersicum)、烟草(N.tabacum)等5种茄科植物的PR10蛋白共同形成了一个进化分枝,亲缘关系较近,也说明PR10蛋白在不同植物中结构较为保守。
2.5 TMV诱导下的NtPR10表达分析
实时荧光定量PCR结果显示(图6),以各个时间点的对照样品为对照,感病品种G28在接种TMV病毒后6 h、12 h、1 d、2 d、4 d、8 d,NtPR10的表达量分别升高5.41、5.56、2.13、3.74、8.81、6.62倍,差异达到极显著水平(P<0.01),TMV侵染4 d时NtPR10达到表达高峰,然后呈现下降趋势,侵染后16 d其表达量降低到对照的44.9%;在抗病品种枯斑三生中,NtPR10基因在TMV侵染后6 h表达量显著上调,达到了高峰,自第12小时至第16天显著低于对照,并于第2天表达量达到最低值,而后至第16天逐渐升高。分析结果可以得知,NtPR10应答了TMV的侵染过程,且其在感病品种和抗病品种中的响应模式存在差异。
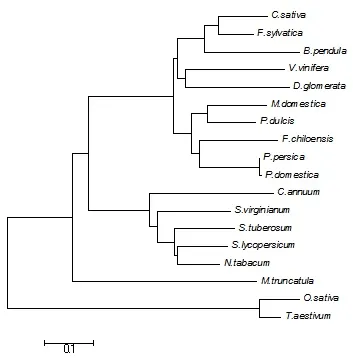
图5 PR10蛋白的系统进化分析Fig. 5 The phylogenetic tree of PR10 proteins

图6 TMV诱导后不同时期NtPR10在感病品种G28和抗病品种枯斑三生中的表达Fig. 6 The expression pattern ofNtPR10in different periods after TMV induction
3 讨 论
普通烟草是异源四倍体,基因组庞大,结构复杂,给一些重要功能基因的克隆带来不便。随着科学技术的不断发展,数据库的逐渐扩大,以及电子克隆技术日趋成熟,从根本上改变了人们对基因克隆的看法,改变了基因克隆长久以来沿用的策略,极大地提高了基因克隆效率,使研究人员的研究焦点更多地转向基因表达与功能的研究。迄今为止,研究人员已经利用电子克隆技术开展了许多重要功能基因的研究[22-24]。张岗等[8]利用电子克隆技术在小麦上成功克隆了病程相关蛋白基因TaPR10,本研究采用电子克隆结合RT-PCR技术成功克隆了烟草NtPR10基因,快捷、高效地获得了目的基因。
烟草NtPR10基因ORF全长483 bp,编码160个氨基酸组成的蛋白序列,其蛋白序列在不同植物中非常保守,不含跨膜区、无信号肽、具有胞内蛋白特征,与前人对于其他植物PR10蛋白的研究结果一致[9],符合PR10的基本特征。已有研究表明,植物PR10蛋白具有体外核酸酶活性和抗菌活性。然而,并非所有的植物PR10蛋白都具有核酸酶活性,从辣椒[4]、刺茄[7]和花生[5]中分离纯化的PR10蛋白均被证实具有体外核酸酶活性,而从马铃薯[21]和苜蓿[25]中分离纯化的PR10蛋白却无体外核酸酶活性,这可能与不同植物PR10蛋白的结构差异相关。本文通过分析得出,烟草PR10蛋白具有与核酸酶活性相关的“P-Loop”结构,而马铃薯PR10蛋白不具有,进一步证明了“P-Loop”结构与核酸酶活性的相关性,推测烟草PR10蛋白具有核酸酶活性,参与烟草抗病生物过程。
植物中PR10蛋白基因的诱导激发因子很多,在不同植物中,PR10蛋白编码基因的诱导表达既有共同的激发因子也存在特异的激发因子,这可能是植物对多样化的逆境胁迫的一种适应。本研究中,在TMV的诱导下,NtPR10基因出现了应答反应,说明TMV是NtPR10基因诱导表达的激发因子。而对于NtPR10基因在感病品种和抗病品种中的表达差异原因,可能是由于抗病品种“枯斑三生”基因组内含有N基因,介导对Ob小种以外的TMV发生免疫反应[26]。在TMV侵染初期NtPR10和N基因共同应答病原菌的侵染,启动防御机制,所以NtPR10基因的表达量有所升高,伴随着TMV的侵染,“枯斑三生”自身的防御机制发生了变化,N基因主导了对TMV的抗性,介导发生了过敏性坏死反应,将TMV病毒限制在枯斑范围内,从而使NtPR10基因表达量有所降低,但枯斑反应发生后,烟株逐渐恢复正常,NtPR10基因的表达量又逐渐恢复到对照水平。因此,分析病原菌诱导下NtPR10基因在抗、感品种中的差异表达机制,以及其与抗病基因的互作将是未来研究的重点。
[1] CHRISTENSEN A B, CHO B H O, NASBY M, et al. The molecular characterization of two barley proteins establishes the novel PR-17 family of pathogenesisrelated proteins[J]. Molecular Plant Pathology, 2002, 3(3):135-144.
[2] VAN LOON L C, REP M, PIETERSE C M J. Significance of inducible defense-related proteins in infected plants[J]. Annu. Rev. Phytopathol., 2006, 44:135-162.
[3] LIU J J, EKRAMODDOULLAH A K M. The family 10 of plant pathogenesis-related proteins: their structure, regulation, and function in response to biotic and abiotic stresses[J]. Physiological and Molecular Plant Pathology, 2006, 68(1): 3-13.
[4] PARK C J, KIM K J, SHIN R, et al. Pathogenesis-related protein 10 isolated from hot pepper functions as a ribonuclease in an antiviral pathway[J]. The Plant Journal, 2004, 37(2): 186-198.
[5] CHADHA P, DAS R H. A pathogenesis related protein, AhPR10 from peanut: an insight of its mode of antifungal activity[J]. Planta, 2006, 225(1): 213-222.
[6] HASHIMOTO M, KISSELEVA L, SAWA S, et al. A novel rice PR10 protein, RSOsPR10, specifically induced in roots by biotic and abiotic stresses, possibly via the jasmonic acid signaling pathway[J]. Plant and cell physiology, 2004, 45(5): 550-559.
[7] LIU X, HUANG B, LIN J, et al. A novel pathogenesisrelated protein (SsPR10) from Solanum surattense with ribonucleolytic and antimicrobial activity is stress-and pathogen-inducible[J]. Journal of plant physiology, 2006, 163(5): 546-556.
[8] 张岗,李依民,张毅,等. 条锈菌诱导的小麦病程相关蛋白TaPR10基因的克隆及特征分析[J]. 中国农业科学,2009,42(1):110-116.
[9] AGARWAL P, AGARWAL P K. Pathogenesis related 10proteins are small, structurally similar but with diverse role in stress signaling[J]. Molecular biology reports, 2014, 41(2): 599-611.
[10] BANTIGNIES B, SEGUIN J, MUZAC I, et al. Direct evidence for ribonucleolytic activity of a PR-10-like protein from white lupin roots[J]. Plant molecular biology, 2000, 42(6): 871-881.
[11] FERNANDES H, MICHALSKA K, SIKORSKI M, et al. Structural and functional aspects of PR-10 proteins[J]. Febs Journal, 2013, 280(5): 1169-1199.
[12] HE M, XU Y, CAO J, et al. Subcellular localization and functional analyses of a PR10 protein gene from Vitis pseudoreticulata in response toPlasmopara viticolainfection[J]. Protoplasma, 2013, 250(1): 129-140.
[13] WU J, KIM S G, KANG K Y, et al. Overexpression of a Pathogenesis-Related Protein 10 Enhances Biotic and Abiotic Stress Tolerance in Rice[J]. The Plant Pathology Journal, 2016, 32(6): 552.
[14] ZHOU X J, LU S, XU Y H, et al. A cotton cDNA (GaPR-10) encoding a pathogenesis-related 10 protein with in vitro ribonuclease activity[J]. Plant Science, 2002, 162(4): 629-636.
[15] WU F, YAN M, Li Y, et al. cDNA cloning, expression, and mutagenesis of a PR-10 protein SPE-16 from the seeds ofPachyrrhizus erosus[J]. Biochemical and biophysical research communications, 2003, 312(3):761-766.
[16] LEE O R, PULLA R K, KIM Y J, et al. Expression and stress tolerance of PR10 genes from Panax ginseng CA Meyer[J]. Molecular biology reports, 2012, 39(3):2365-2374.
[17] 谢纯政,刘海燕,李玲,等. 植物病程相关蛋白PR10研究进展[J]. 分子植物育种,2008,6(5):949-953.
[18] CHEN Z Y, BROWN R L, DAMANN K E, et al. PR10 expression in maize and its effect on host resistance againstAspergillus flavusinfection and aflatoxin production[J]. Molecular plant pathology, 2010, 11(1):69-81.
[19] HE M, XU Y, CAO J, et al. Subcellular localization and functional analyses of a PR10 protein gene fromVitis pseudoreticulatain response toPlasmopara viticolainfection[J]. Protoplasma, 2013, 250(1): 129-140.
[20] TAMURA K, STECHER G, PETERSON D, et al. MEGA6: molecular evolutionary genetics analysis version 6.0[J]. Molecular biology and evolution, 2013, 30(12): 2725-2729.
[21] MATTON D P, BRISSON N. Cloning, expression, and sequence conservation of pathogenesis-related gene transcripts of potato[J]. Molecular plant-microbe interactions: MPMI, 1988, 2(6): 325-331.
[22] 张德礼,孙晓静,凌伦奖,等. 人类SR蛋白超家族新成员—SFRS12(SRrp508)的基因克隆和特性分析[J].遗传学报,2002,29(5):377-383.
[23] LIU X, LIN F, WANG L, et al. The in silico map-based cloning of Pi36, a rice coiled-coil-nucleotide-binding site-leucine-rich repeat gene that confers race-specific resistance to the blast fungus[J]. Genetics, 2007, 176(4):2541-2549.
[24] WU H, NI Z, YAO Y, et al. Cloning and expression profiles of 15 genes encoding WRKY transcription factor in wheat (Triticum aestivemL.)[J]. Progress in Natural Science, 2008, 18(6): 697-705.
[25] SAMAC D A, PENUELA S, SCHNURR J A, et al. Expression of coordinately regulated defence response genes and analysis of their role in disease resistance inMedicago truncatula[J]. Molecular plant pathology, 2011, 12(8): 786-798.
[26] DAWSON W O, HILF M E. Host-range determinants of plant viruses[J]. Annual review of plant biology, 1992, 43(1): 527-555.
Cloning and Expression Analysis of a Pathogenesis Related Protein Gene NtPR10 in Tobacco (Nicotiana tabacum)
ZHANG Yu1,2, ZHANG Zenglin1, JIANG Caihong1, CHANG Aixia1, YANG Aiguo1, LUO Chenggang1, WANG Shaomei1, WANG Yuanying1*
(1. Tobacco Research Institute of Chinese Academy of Agricultural Sciences, Qingdao 266101, China; 2. Graduate School of Chinese Academy of Agricultural Sciences, Beijing 100081, China)
In order to investigate the biological function of the tobacco pathogenesis-related protein PR10,NtPR10gene was obtained from the tobacco variety G28, which is 483 bp in length, encoding 160 amino acids. The NtPR10 protein belongs to the pathogenesis related protein family containing the “Bet_v_I” domain and the “P-loop” domain which is related to nuclease activities. The bioinformatics analysis indicated that NtPR10 did not contain transmembrane region, no signal peptide, and exhibited intracellular localization features by TMHMM, SignalP and PrositeScan. The expression pattern of the gene in the resistant and susceptible cultivars was studied by RT-qPCR. The results showed that theNtPR10gene was significantly up-regulated in the susceptible cultivar G28 after TMV treatment. While in resistant cultivarSumsunNN,NtPR10was up-regulated at 6 hours after TMV infection, and down-regulated between 12 hours and 8thday, and then increased gradually to the pre-infection level after 16thday. From above analysis,NtPR10responses to TMV infection, and the expression trend is opposite in resistant and susceptible cultivars, suggesting that theNtPR10might have an important function in the process of TMV infection. Results from this study provide the foundation for further analysis of the biological function ofNtPR10.
tobacco; pathogenesis-related protein; PR10; gene clone; expression
S572.03
1007-5119(2017)03-0001-07 DOI:10.13496/j.issn.1007-5119.2017.03.001
国家自然科学基金项目“烟草病程相关蛋白PR10抗病分子机理研究”(31240013);中国农业科学院烟草研究所青年科学基金项目“对烟草赤星病不同抗性品种蛋白质组差异分析及抗性蛋白挖掘”(2016B04);中国烟草总公司四川省公司科技项目“优质适产烤烟新品种选育与推广”(SCYC201501)
张 玉(1982-),男,助理研究员,在读博士,主要从事烟草遗传育种研究。E-mail:zhangyu@caas.cn。*通信作者,E-mail:wangyuanying@caas.cn
2016-12-22
2017-03-23
