扩散型与繁殖型松材线虫数字基因表达谱对比分析
2017-06-19田浩楷柳小龙张志伟赵莉蔺山西农业大学林学院山西太谷0080中国科学院动物研究所农业虫害鼠害综合治理研究国家重点实验室北京000中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室北京009
田浩楷, 张 帅, 柳小龙, 张志伟, 赵莉蔺山西农业大学林学院,山西 太谷 0080; 中国科学院动物研究所,农业虫害鼠害综合治理研究国家重点实验室,北京 000; 中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 009
扩散型与繁殖型松材线虫数字基因表达谱对比分析
田浩楷1,2, 张 帅3, 柳小龙1,2, 张志伟1*, 赵莉蔺2*
1山西农业大学林学院,山西 太谷 030801;2中国科学院动物研究所,农业虫害鼠害综合治理研究国家重点实验室,北京 100101;3中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193
【目的】松材线虫作为林业重大外来入侵种,扩散型幼虫的形成对其传播扩散起着非常重要的作用,但扩散虫态的形成与维持机制尚未阐明。【方法】通过构建松材线虫数字基因表达谱(DGE),从滞育状态的维持、化学感受、代谢途径等方面分析松材线虫不同虫态的基因表达差异。【结果】参考松材线虫基因组数据,鉴定出2种扩散型幼虫(LⅢ,LⅣ)和繁殖型幼虫(Ln)各有11184、8533和10781个基因。相对于繁殖型虫态,大多数基因在LⅣ中下调表达,该虫态中特异上调表达的基因有化感受体基因、核受体基因以及一些代谢相关基因。推测这可能与扩散型线虫滞育状态的维持相关,并在其生理功能如化学感受和媒介/寄主互作中发挥作用。GO和Pathway富集分析显示,多数代谢相关通路在LⅣ中下调表达,而在LⅢ中的表达均活跃。【结论】以上结果与LⅣ处于不进食、总体代谢水平较低等生理状态的表型相一致。
松材线虫; 扩散型虫态; 数字基因表达谱; 差异表达基因
松材线虫Bursaphelenchusxylophilus(Steiner & Buhrer,1934) Nickle,1970是世界公认的检疫性有害生物,是松树萎蔫病(pine wilt disease,PWD)的主要致病因子(Mamiya,1983)。该线虫在原产地北美并未造成大面积危害,但被引入亚洲和欧洲等国家后大规模暴发,短时间内致大量松树死亡,并不断扩散蔓延(詹开瑞等,2013; Wingfieldetal.,1982)。
松材线虫的生活史分为2个周期:繁殖周期(propagative pathway)和扩散周期(dispersal pathway)(Ishibashi & Kondo,1977)。每年夏季,松材线虫在树体内经过卵、繁殖型1~4龄幼虫(L1-L4, Ln)发育为成虫。在环境不利的条件下,如低温、干燥、种群密度过高等,松材线虫转变为扩散型Ⅲ龄幼虫(dispersal third stage larva, LⅢ);LⅢ在媒介天牛存在的情况下转变为扩散型Ⅳ龄幼虫(dispersal fourth stage larva, LⅣ)。LⅢ和LⅣ均为滞育型虫态,有很强的抗逆性,它们即使不进食也可以生存数月(Mamiya,1983; Zhaoetal.,2014)。
滞育型虫态在线虫中普遍存在(Hu,2007)。如模式线虫秀丽隐杆线虫Caenorhabditiselegans(Maupas),在环境适宜的条件下,由受精卵经过L1-L44龄幼虫发育为成虫;而遇到食物缺乏、种群密度增大、温度升高等不利环境条件时,C.elegans会形成一种滞育型虫态dauer。相对于繁殖型幼虫,dauer不再进食且身体结构特化,可以抵御干燥、高温或食物短缺等外界不良环境条件(Cassada & Russell,1975; Golden & Riddle,1984; Riddleetal.,1981);当外界环境恢复时,dauer会转变为L4进入繁殖周期。松材线虫与C.elegans相比,特殊之处在于其可以形成2种滞育虫态:LⅢ相当于C.elegans的pre-dauer,LⅣ相当于C.elegans的dauer;但是,C.elegans的pre-dauer不能长期存在,松材线虫的LⅢ却可以长期稳定存在。LⅣ除抗逆性强以外,还拥有特化的功能,即可以准确识别与定位松墨天牛MonochamusalternatusHope,以便被其携带并传播扩散,LⅢ则不能被携带(Zhaoetal.,2007,2013)。代谢方面,LⅢ仍然可以进食,同时储存大量脂肪粒;LⅣ不再进食,靠分解LⅢ积累的脂类物质维持生命活动或蜕皮转变为成虫。
在滞育虫态差异表达基因的挖掘方面,Jonesetal.(2001)通过基因表达系列分析(serial analysis of gene expression,SAGE)技术鉴定出358个C.elegansdauer特异表达的转录本,其中,热激蛋白hsp-12.6为高表达的转录本。Wang & Kim (2003)利用基因芯片技术发现了2430个在dauer虫态中差异表达的基因。松材线虫2种特有的扩散型虫态(LⅢ和LⅣ)的形成除了受环境因素的调控外,还与其携带者产生的化学信号诱导有关(Zhaoetal.,2014),推测其在转型发育调控中与C.elegans存在差异。Kikuchietal.(2007)通过对比松材线虫Ln和LⅣ的EST谱发现了一些LⅣ特有的EST;但未涉及LⅢ,且基因分析主要侧重于致病性方面。Yanetal.(2012)对松材线虫与拟松材线虫BursaphelenchusmucronatusMamiya et Enda比较转录组的研究也是集中在2种线虫的致病性与解毒相关基因的对比分析方面。目前,有关松材线虫特有的2种扩散型虫态的形成与维持机制的研究甚少。因此,本研究运用数字基因表达谱分析技术,着重从滞育状态的维持、化学感受、代谢途径等方面分析松材线虫不同虫态的基因表达差异,为研究其扩散型虫态的形成与维持机制提供分子基础。
1 材料与方法
1.1 供试松材线虫来源与培养方法
供试松材线虫均来源于浙江省富阳市新登镇马鞍山。繁殖型线虫分离自采集的疫木,经过分离鉴定后挑取目的虫态,使用常规PDA(potato dextrose agar)培养基(Difco,BD)接种灰葡萄孢菌进行进一步增殖培养。LⅢ分离自采集的疫木,LⅣ分离自采集的松墨天牛成虫。
繁殖型线虫样本制备:将混合龄期繁殖型线虫置于90 mm平板PDA培养基上生长10 d,用灭菌ddH2O将线虫从培养基洗下,3000 r·min-1离心3 min,收集上清液。LⅢ样本制备:把疫木砍成3~5 cm细木条,用贝尔曼漏斗法(baermann funnel technique)分离线虫(Baermann,1917),镜检,去除非LⅢ线虫。LⅣ样本制备:解剖初羽化天牛,置于35 mm培养皿中,加入灭菌ddH2O,浸泡30 min后,选取线虫量多的样品进行贝尔曼漏斗分离,镜检,去除非LⅣ线虫。采用蔗糖悬浮(sucrose flotation)离心方法去除真菌菌丝和其他杂质,进一步纯化线虫(Freckmanetal.,1975),离心,去除上清后将线虫用液氮速冻,2 h后转入-80 ℃冰箱保存待用。
1.2 样本RNA提取、建库及基因表达谱测序
RNA提取使用RNeasy Micro Kit(Qiagen,德国)试剂盒。RNA提取后,使用NanoDrop DN-1000(NanoDrop Technologies,美国)检测样品质量,选取质量好的样本进行建库测序。具体流程:先利用Oligo(dT)的磁珠(beads)富集总RNA中mRNA,并引导反转录合成双链cDNA,采用4碱基识别酶NlaIII酶切双链cDNA,链接Illumina adaptor1,利用MmeI酶切3′端CATG下游17 bp碱基,并在3′端链接Illumina adaptor2;再加入Primer GX1和Primer GX2进行PCR扩增;样本扩增后,通过6% TBE PAGE胶回收95碱基条带,纯化后通过Illumina基因表达测序法测序。
1.3 差异表达基因的筛选
利用泊松分布的算法筛选2个文库间的差异表达基因,对差异检验的P-value做多重假设检验校正,通过控制错误发现率(false discovery rate,FDR) 决定P-value的域值。根据基因的表达量(TPM值),计算该基因在不同样本间的差异表达倍数(Fold)。差异表达基因的筛选条件:FDR≤0.001,Log2>1。FDR值越小,差异倍数越大,说明表达水平差异越显著。当Log2Fold≥1时,表示该基因表达上调;当Log2Fold≤-1时,表示表达下调;当∣Log2Fold∣<1时,表示表达无差异(王海英等,2016; 姚敏磊等,2016)。
1.4 差异表达基因的GO功能和KEGG通路分析
差异表达基因的GO (gene ontology)分析数据来自Gene ontology 数据库(http:∥www. geneontology.org/)。用在线软件Veen (http:∥bioinfogp.cnb.csic.es/tools/venny/index.html)对差异表达基因进行聚类分析。用KEGG数据提供的代谢通路信息进行Pathway富集分析,当Qvalue≤0.05时,表示差异表达基因在该通路中显著富集。
2 结果与分析
2.1 测序数据统计
经过Illumina测序,去除纯接头及含N序列和拷贝数<2的标签后,得到高质量测序标签(clean tags)。各虫态Clean tags统计如表1所示。
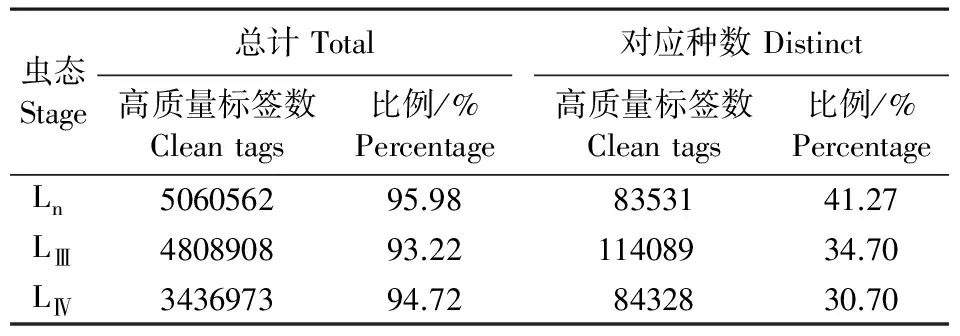
表1 线虫不同虫态clean tag数Table 1 Clean tags from different larval stages of B. xylophilus
测序饱和度曲线表明,当测序tag数大于等于20×100 K(2 M)时,基因数已经趋于饱合(图1A)。clean tags在3个样本中都大于3 M,表明表达谱测序的数据量较为充足。从检测出的基因数量看,Ln和LⅢ接近,且都显著高于LⅣ(约1.5倍)。这表明Ln和LⅢ基因表达比LⅣ活跃,因此有更多的转录本(transcripts)被检测到。
clean tags拷贝数分布统计(图1B)显示,拷贝数大于100的高表达tags在3个样本中占80%左右,数量上占绝对优势;而拷贝数小于5的低表达tags种类非常丰富,达到60%左右。这说明少量种类的丰度极高, 而大部分种类的表达水平很低,该结果符合细胞中mRNA的显著特征。因此,从整体上评估数据是正常的。
基于参考基因数据库,产生参考基因总数15971个,其中,含有CATG序列的14144个(占88.56%);同时产生70912个参考tag序列,可以map到单个基因的tag数为69278个(占97.7%)。各虫态检测出的基因总数分别为Ln10781个、LⅢ11184个、LⅣ8533个,其中各有8531(占79.1%)、8846(占79.1%)、7049(占82.6%)个基因被注释。这是EST分析结果的2倍以上,且比EST有更准确的基因表达量信息(Kikuchietal.,2007),有助于不同龄期线虫基因表达水平的比较。
2.2 差异表达基因总体统计
相对于Ln,LⅢ有1585个(占14.2%)基因上调表达,1440个(占12.9%)基因下调表达;相对于Ln,LⅣ有568个(占6.7%)基因上调表达,3723个(占43.6%)基因下调表达。这些数据显示,LⅢ中上、下调表达的基因数量较少且相当,而在LⅣ中仅少量基因的表达上调,约一半基因均下调表达。相对于Ln,LⅢ与LⅣ上、下调表达一致的基因分别为303和1262个;LⅣ下调而LⅢ上调的基因161个,主要包括热激蛋白基因(heat shock protein,HSP)以及一些细胞色素P450基因(cytochrome P450,CYP),可能与LⅢ抗逆性强相关;LⅣ上调而LⅢ下调的基因5个(图2A),其中包括annexin基因,该基因在C.elegans中与其咽部吞食作用有关(Rogersetal.,2001),而在LⅣ中可能与其不再进食的生理状态有关。LⅣ相对于LⅢ,下调基因居多(上调298个,下调4115个)。以上结果表明,LⅢ基因表达较为活跃,而绝大多数基因在LⅣ中下调表达,可能由于LⅣ不再进食,仅依靠LⅢ阶段积累的脂肪类物质提供营养。GO分析显示,相对于Ln,LⅢ与LⅣ差异表达的基因集中在生物过程和分子功能方面(图2B)。
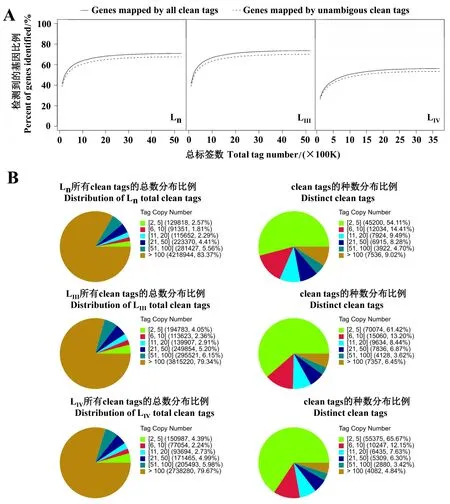
A:测序饱和度分析;B:clean tag拷贝数的分布。A: Analysis of sequencing saturation; B: Distribution of clean tags.图1 测序质量评估Fig.1 Sequencing quality assessment
2.3 不同虫态的高表达转录本分析
高表达转录本(abundantly represented transcripts)是生物体在特定发育阶段内表达量较高的基因,参与该阶段主要生理机能的维持。在LⅢ中,HSP、FMRFamide神经肽相关蛋白、精子主要蛋白(major sperm protein)、山梨醇脱氢酶(sorbitol dehydrogenase)、层黏连蛋白受体(laminin receptor)等为表达量较高的转录本(表2)。
在LⅣ中,高表达转录本主要包括FMRFamide神经肽相关蛋白、肌球蛋白(myosin)、脂肪酸延长相关蛋白、胚胎发育晚期富集蛋白(late embryogenesis abundant-like protein)以及一些功能未知的重要假定蛋白(hypothetical protein)(表3)。这些基因的功能集中在发育调节、神经感受以及中间代谢方面,推测可能与LⅣ维持滞育状态、感受外界环境和降低代谢率相关。
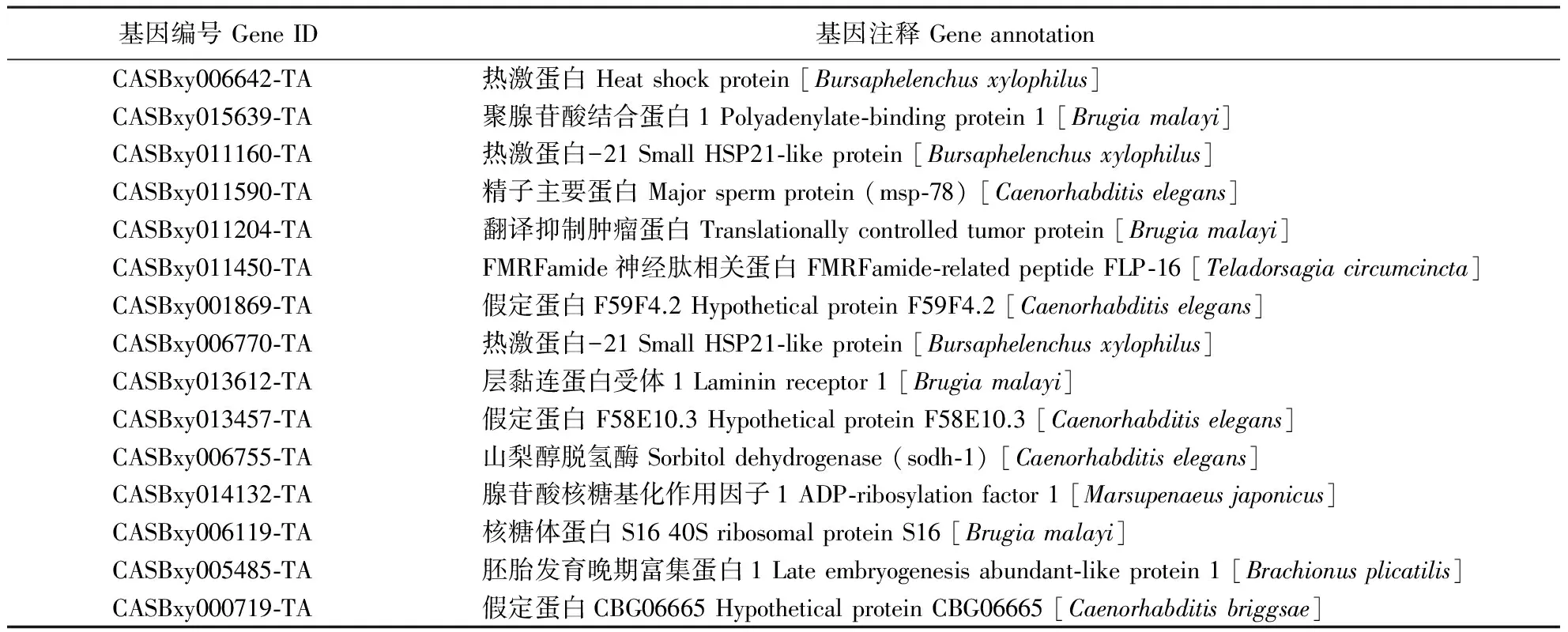
表2 LⅢ表达量最高的15种转录本Table 2 Most abundantly represented transcripts in LⅢ (top 15)
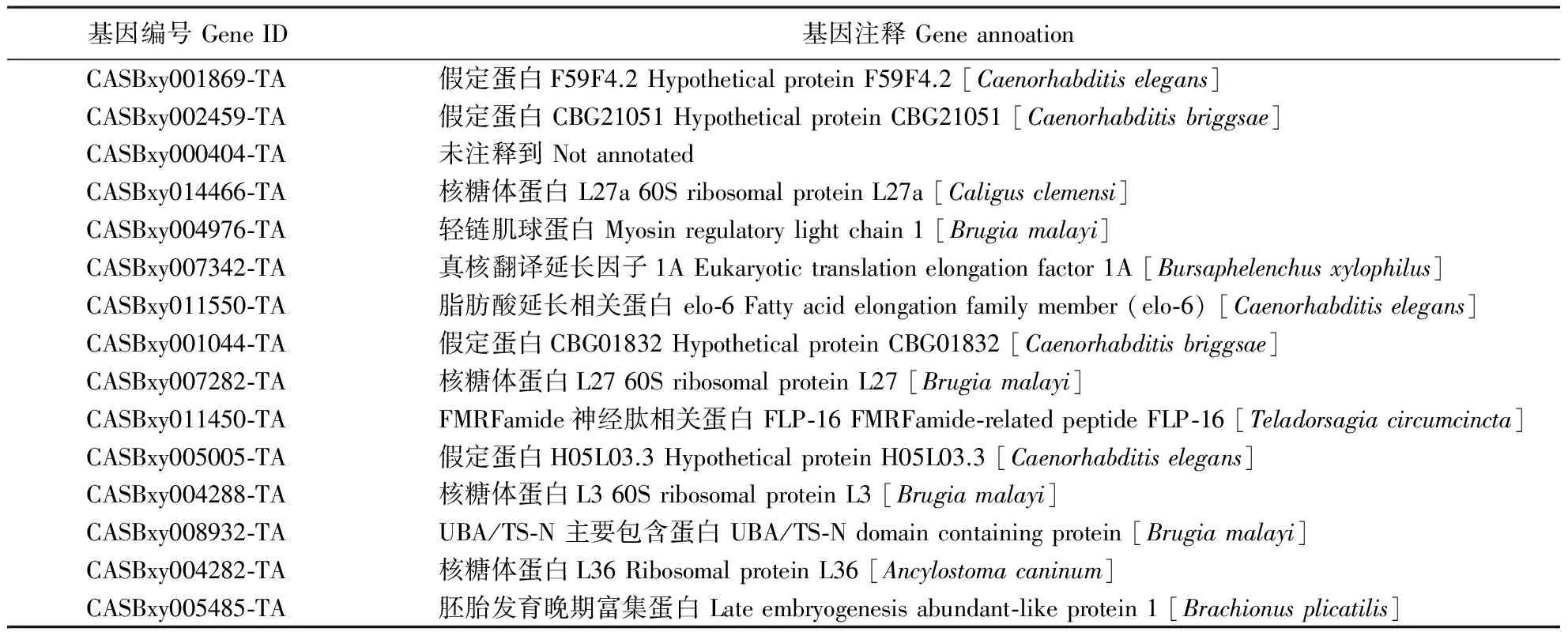
表3 LⅣ表达量最高的15种转录本Table 3 Most abundantly represented transcripts in LⅣ (top 15)
繁殖型线虫中,与LⅢ类似,精子主要蛋白和山梨醇脱氢酶等基因表达量也很高。在线虫中,精子主要蛋白主要参与细胞质骨架的装配以及细胞运动相关功能。与扩散型虫态不同的是,繁殖型线虫中,核糖体蛋白、在糖酵解途径中的3-磷酸甘油醛脱氢酶(glyceraldehyde-3-phosphate dehydrogenase),以及细胞结构相关蛋白如副肌球蛋白(paramyosin)、胶原蛋白(collagen)等表达量相对较高(表4)。这表明繁殖型线虫的细胞生长与分裂活动及代谢水平高于扩散型虫态。此外,在LⅣ和LⅢ中表达量高的基因如胚胎发育晚期丰富蛋白和FMRFamide神经肽等在Ln中的表达量均不高,表明此类基因可能与扩散型虫态滞育状态的维持有关。
2.4 扩散型线虫LⅣ中特异上调表达的基因
相对于Ln,LⅣ特异上调表达的基因有260个(图2A)。表5是去除未成功注释和注释到未知功能蛋白或假定蛋白的基因,同时略去一些功能广泛的蛋白激酶,按照表达量排序筛选出的前25个基因。
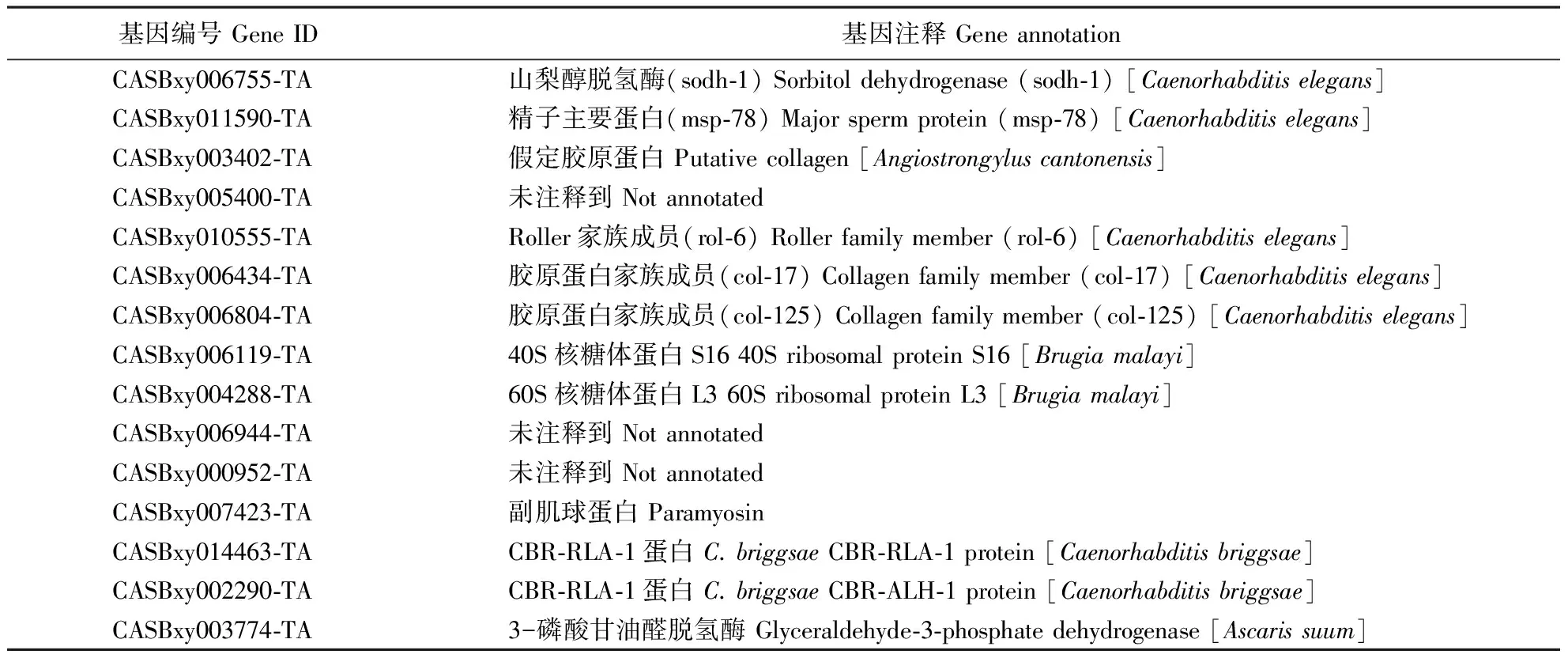
表4 Ln表达量最高的15种转录本Table 4 Most abundantly represented transcripts in Ln (top 15)
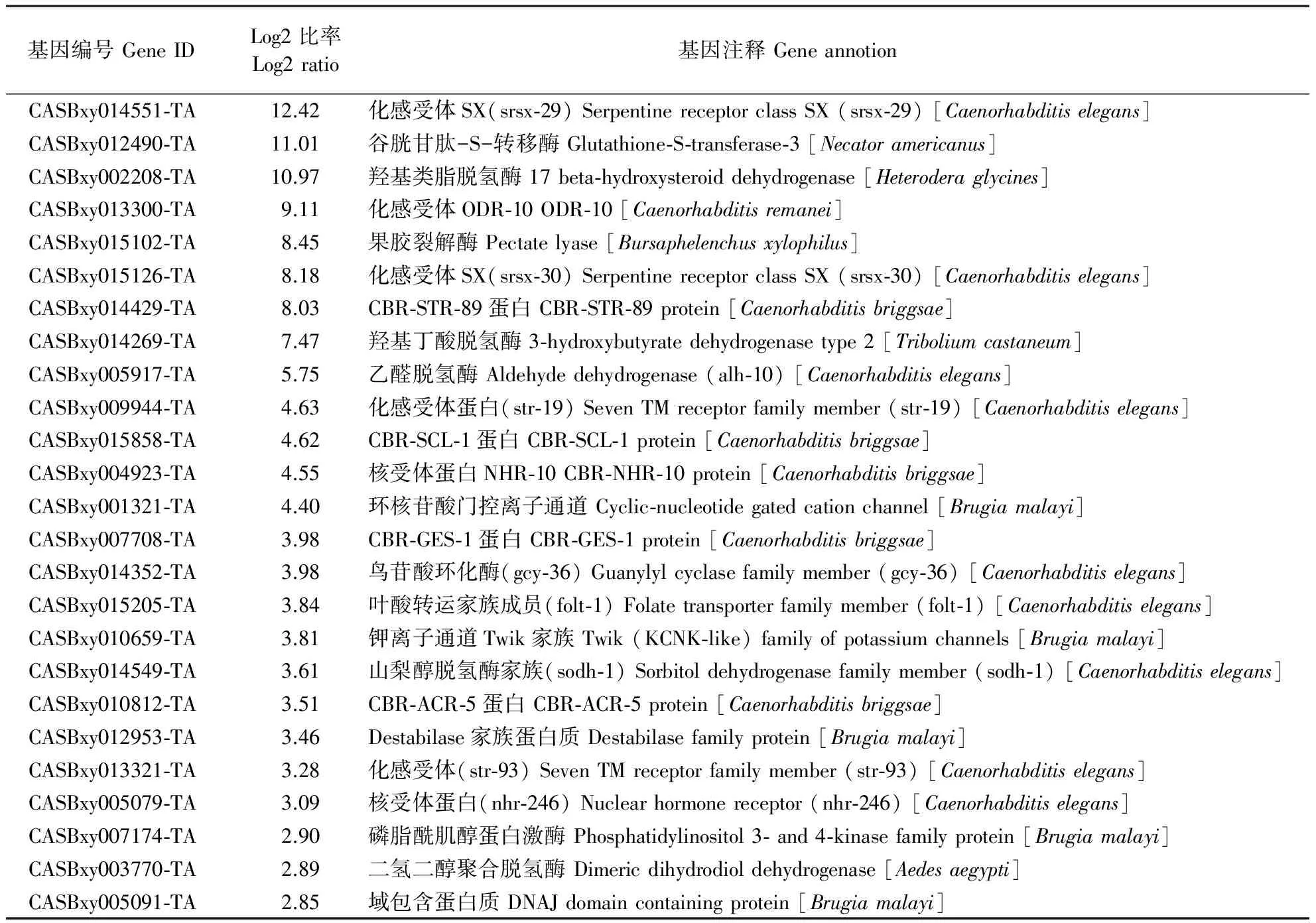
表5 LⅣ特异上调表达的部分基因Table 5 Selected genes specifically up-regulated in LⅣ
数据显示,化感受体(serpentine receptor)基因在LⅣ中特异上调幅度较大,包括srsx-29、srsx-30、str-19、str-89、str-93、odr-10共6个相关基因,表明此类基因可能与LⅣ虫态对外界化学信号更为敏感有关。在自然条件下,LⅣ通过有效识别媒介天牛释放的化学信号而进入天牛体内,传播扩散至新的寄主松树后选择合适的时间从天牛体内逸出,这些过程都需要化感受体基因的参与。
上调幅度较大的还有核受体(nuclear receptor)基因nhr-10和nhr-246。核受体基因一般直接调控下游效应基因的表达,以启动特定的代谢或发育通路。这些核受体可能调控LⅣ特异的代谢或发育途径。一些代谢酶类也位列其中,如谷胱甘肽-S-转移酶(glutathione-S-transferase)、羟基类脂脱氢酶(17beta-hydroxysteroid dehydrogenase)、羟基丁酸脱氢酶(3-hydroxybutyrate dehydrogenase type 2)、二氢二醇聚合脱氢酶(dimeric dihydrodiol dehydrogenase)、UDP-葡萄糖醛酸转移酶(UDP-glucuronosyl transferase)等。
2.5 扩散型线虫代谢途径分析
通过KEGG pathway的富集分析,着重筛选了不同虫态在三大代谢途径中的差异基因,这些差异基因均为Ln、LⅢ和LⅣ表达谱中共同注释到的转录本。
糖酵解途径中,相对于Ln,LⅢ有2个基因上调表达,4个基因下调表达,其余基因表达无显著差异。上调表达的基因有果糖—二磷酸醛缩酶(fructose-bisphosphate aldolase)和乙醛脱氢酶(aldehyde dehydrogenase),下调表达的基因有葡萄糖激酶(glucokinase)、3-磷酸甘油醛脱氢酶等。而LⅣ在该途径中大部分基因下调表达。这可能是由于LⅣ线虫需要的能量较少;但LⅢ糖酵解途径仍然保持一定的活跃程度,维持着主要的能量输出(图3)。
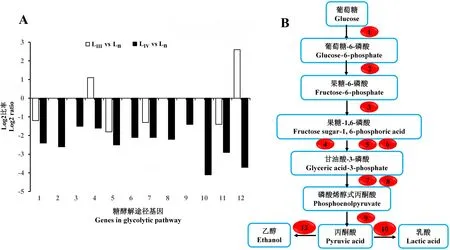
A:糖酵解途径中部分基因的表达差异;B:差异基因在糖酵解途径中的分布。A: The differentially genes expression in glycolytic pathway; B: The distribution of differential genes in glycolytic pathway.1:葡萄糖激酶 Glucokinase; 2:葡萄糖-6-磷酸异构酶 Glucose-6-phosphate isomerase; 3:果糖-1,6-二磷酸酶 Fructose-1,6-bisphosphate aldolase (FBA); 4:果糖—二磷酸醛缩酶 Fructose-bisphosphate aldolase; 5:3-磷酸甘油醛脱氢酶 Glyceraldehyde 3-phosphate dehydrogenase; 6:磷酸甘油酸激酶 Phosphoglycerate kinase; 7:磷酸甘油变位酶 Phosphoglycerate mutase; 8:磷酸丙酮酸水合酶 Phosphopyruvate hydratase; 9:丙酮酸激酶 Pyruvate kinase; 10:乳酸脱氢酶连接酶 L-lactate dehydrogenase; 11:乙酰辅酶A连接酶 Acetate-CoA ligase; 12:醛脱氢酶 Aldehyde dehydrogenase (NAD+)。图3 LⅢ、LⅣ相对于Ln糖酵解途径基因表达变化Fig.3 Expression profiling of genes in glycolytic pathway of LⅢand LⅣ vs Ln
从三羧酸循环(tricarboxylic acid cycle,TCA)来看,LⅢ相对于Ln有3个基因上调表达,4个基因表达量下调(图4)。上、下调基因较少且数目相近,呈平衡状态,且该通路的其他基因表达量无变化,表明松材线虫LⅢ有氧代谢仍比较旺盛。而LⅣ相对于Ln共有11个基因的表达量改变,其中,只有丙酮酸脱氢酶复合体E1组分的部分Tag表达量上调,其余基因全部下调表达(图4)。这表明TCA循环通路的基因在LⅣ中的表达低于Ln和LⅢ。TCA是线粒体中能量代谢的主要通路,LⅣ有氧代谢水平降低,与其不进食、不活跃的生理状态相符。
脂肪酸代谢方面,LⅢ相比繁殖型线虫较为活跃,通路大多数基因上调表达,包括脂肪酰CoA氧化酶(fatty acyl-CoA oxidase)、脂肪酰CoA脱氢酶(fatty acyl-CoA dehydrogenase)、3-羟基脂肪酰CoA脱氢酶(3-hydroxyacyl-CoA dehydrogenase)等。而在LⅣ中,仅脂肪酰CoA氧化酶上调表达,其余大多数基因都下调表达(图5)。
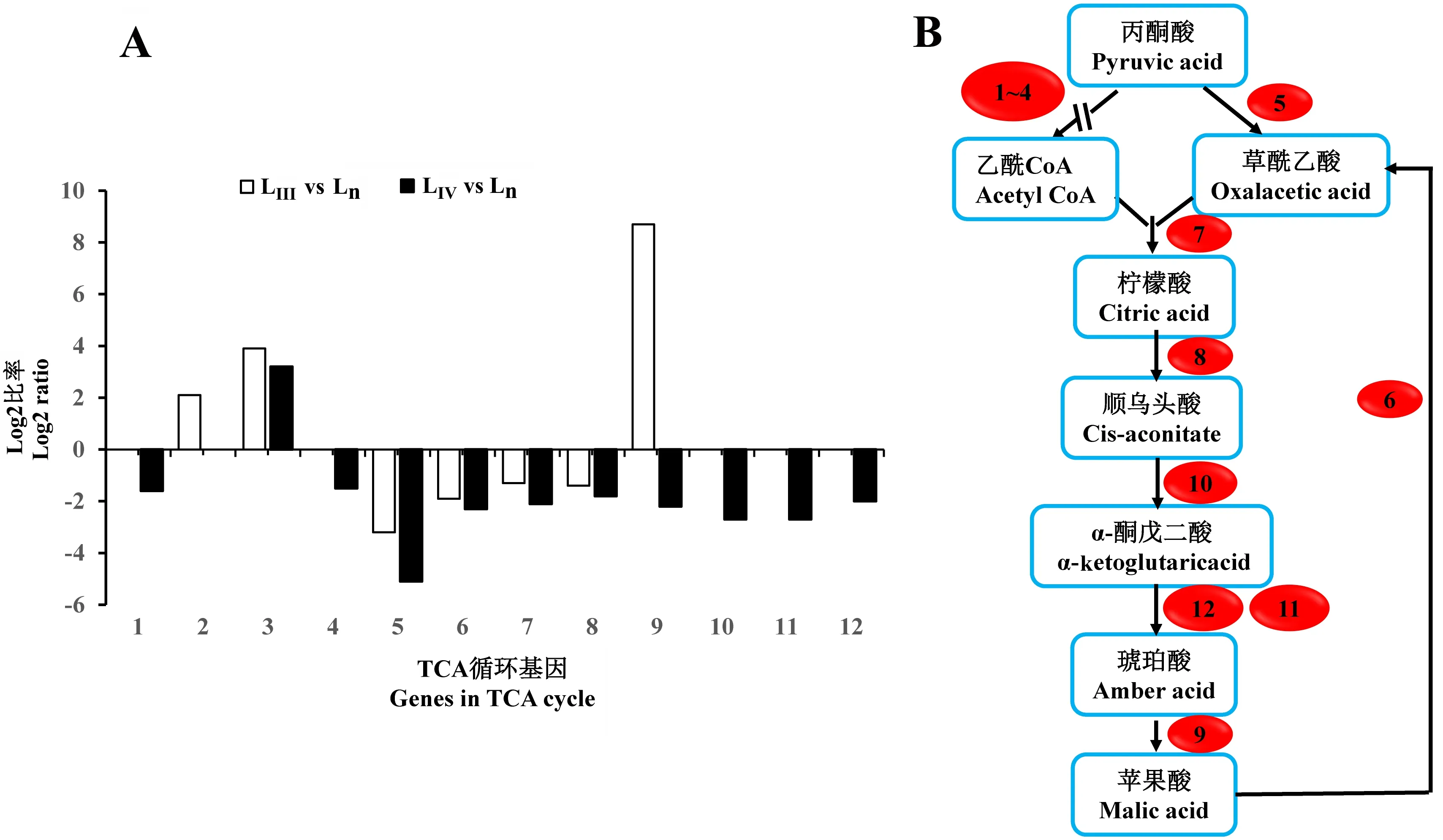
A:TCA循环中部分基因的表达差异;B:差异基因在TCA循环中的分布。A: The differentially genes expression in TCA cycle; B: The distribution of differential genes in TCA cycle.1:磷酸烯醇式丙酮酸羧化酶 Phosphoenolpyruvate carboxykinase; 2:丙酮酸脱氢酶复合体E2 Pyruvate dehydrogenase complex E2 component; 3:丙酮酸脱氢酶复合体E1 Pyruvate dehydrogenase complex E1 component; 4:二氢硫辛酰胺脱氢酶 Dihydrolipoamide dehydrogenase; 5:丙酮酸羧化酶 Pyruvate carboxylase; 6:苹果酸脱氢酶 Malate dehydrogenase; 7:柠檬酸合酶 Citrate synthase; 8:顺乌头酸酶 Aconitase; 9:琥珀酸脱氢酶 Succinate dehydrogenase; 10:异柠檬酸脱氢酶 Isocitrate dehydrogenase;11:琥珀酰-CoA合成酶 Succinyl-CoA synthetase; 12:α-酮戊二酸脱氢酶 α-ketoglutarate dehydrogenase。图4 LⅢ、LⅣ相对于Ln TCA循环基因表达变化Fig.4 Expression profiling of genes in TCA cycle of LⅢ and LⅣ vs Ln
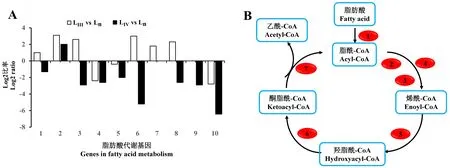
A:脂肪酸代谢中部分基因的表达差异;B:差异基因在脂肪酸代谢通路中的分布。A: The differentially genes expression in fatty acid metabolism; B: The distribution of differential genes in the pathway of fatty acid metabolism.1:长链脂肪酸-CoA连接酶 Long chain fatty acid-CoA ligase; 2:脂肪酰CoA氧化酶 Fattyacyl-CoA oxidase; 3:脂肪酰CoA脱氢酶 Fatty acyl-CoA dehydrogenase; 4:琥珀酸脱氢酶 Succinate dehydrogenase; 5:不饱和脂肪酰CoA水合酶 Unsaturated acyl-CoA hydratase; 6:3-羟基脂肪酰CoA脱氢酶 3-hydroxyacyl-CoA dehydrogenase; 7:3-酮基脂肪酰硫解酶 3-ketoacyl-CoA thiolase; 8:十二氯烷烃辅酶A异构酶 Dodecenoyl-CoA isomerase; 9:乙酰辅酶A酰基转移酶 Acetyl-CoA C-acetyltransferase; 10:乙醇脱氢酶 Alcohol dehydrogenase。图5 LⅢ、LⅣ相对于Ln脂肪酸代谢基因表达变化Fig.5 Expression profiling of genes in fatty acid metabolism of LⅢ and LⅣ vs Ln
3 讨论与结论
本研究表明,相对于Ln而言,LⅢ基因表达仍较为活跃,而LⅣ大多数基因下调表达。LⅣ上调表达的基因主要集中于维持滞育状态、提高对不良环境的耐受力和感受外界信号方面。从基因表达模式看,LⅣ更接近于C.elegans的dauer虫态,而LⅢ更接近一个正在转变的状态,对应于C.elegans中L1转变为dauer的过程(Jonesetal.,2001; Wang & Kim,2003)。
不同于繁殖型线虫中细胞生长与分裂相关转录本的高表达,扩散型线虫共有的高表达转录本以及上调表达的基因则集中体现在滞育状态的维持及对不良环境耐受力的提高等方面。在LⅢ和LⅣ表达谱的高表达转录本中均检测到了FMRFamide神经肽相关蛋白;在LⅢ高表达转录本中检测到山梨醇脱氢酶;而山梨醇脱氢酶、谷胱甘肽S-转移酶等在LⅣ中特异上调表达。此结果与Kikuchietal.(2007)通过EST研究发现的扩散型线虫特异的高表达转录本基本一致。其中,松材线虫中的FMRFamide神经肽相关蛋白基因与C.elegans中的flp-16基因同源,flp-16基因在C.elegans中参与抑制咽部肌肉动作电位的传导(Rogersetal.,2001)。此基因在松材线虫LⅣ中大量表达,可能与其咽部活动的抑制有关,这与LⅣ不再进食和咽部食道球不活动的表型相符。谷胱甘肽S-转移酶在C.elegansdauer虫态中也上调表达,它与内源或外源有毒代谢物质的排出有关(Liuetal.,2004; McElweeetal.,2004)。另外,细胞色素P450、短链化合物脱氢酶/还原酶、UDP-葡萄糖醛酸基转移酶等基因在C.elegans中也具有类似的作用(McElweeetal.,2004),这些基因在松材线虫LⅣ中同样上调表达,说明此类基因在LⅣ生理状态的维持中可能起重要作用。综上所述,松材线虫扩散型虫态的高表达基因与C.elegansdauer虫态具有相似之处,进一步体现了线虫滞育虫态的普遍性与一致性。
在松材线虫LⅣ特异上调表达的基因中,上调表达最显著的是一些化感受体基因,这些化感受体都属于G蛋白偶联受体(G protein-coupled receptor,GPCR)家族;而在C.elegansdauer虫态显著上调的基因中未见这些感受体(Jonesetal.,2001; Wang & Kim,2003)。这表明作为植物寄生线虫,松材线虫LⅣ与自由生活的C.elegansdauer虫态在化学感受方面存在差异。Kimetal.(2009)研究表明,2个多次跨膜G蛋白偶联受体(srbc-64和srbc-66)相应的突变体在dauer诱导信息素存在的条件下不能形成滞育虫态,从而证明该受体参与C.elegans对dauer信息素(C6、C7、C9)的响应。这表明化感受体参与了线虫的转型发育。此外,化感受体的表达具有时间与空间差异性(Bargmann,2006; Chenetal.,2005; Thomasetal.,2005,2008)。上调显著的6个化感受体基因中,2个属于srsx家族(srsx-29、srsx-30),3个属于str家族(str-19、str-89、str-93),1个属于odr家族(odr-10)。这些基因中除了odr-10被证明在C.elegans中感受双乙酰(Senguptaetal.,1996)外,其他基因的具体功能尚不清楚。在松材线虫中这些基因可能与LⅣ对外界化学信号的感受相关。本研究的前期工作证实了松材线虫LⅢ向天牛蛹室的聚集、LⅢ到LⅣ的转变以及LⅣ识别与定位初羽化的媒介天牛并向其气管系统迁移等过程都受到化学信号的介导(Zhaoetal.,2013,2014)。当松材线虫被天牛携带转移至新的寄主后,需要在适当的时机逸出天牛体内,此过程可能也需要感受特定的化学信号。有研究显示,健康松树产生的β-香叶烯(β-myrcene)可以促进线虫逸出天牛体内(Enda & Ikeda,1983; Ishikawaetal.,1986; Stamps & Linit,1998)。Burnell & O′Halloran (2004)研究表明,昆虫寄生性线虫Heterorhabditisbacteriophora的化学感受谱在其扩散型虫态形成过程中会产生适应性的变化,体现在对细菌代谢物敏感性的降低和对长链醇及昆虫特异挥发物敏感性的增加。总之,线虫需要特定的化感受体去感受这些化学信号。因此推测,松材线虫LⅣ中特异上调表达的化感受体,对其准确地进出媒介天牛并进一步传播扩散起重要作用;但具体的感应机制有待深入研究。
代谢相关基因分析表明,松材线虫LⅣ的总体代谢水平低于Ln,与C.elegansdauer虫态总体代谢率的降低相符(Houthoofdetal.,2002; O′Riordan & Burnell,1989; Vanfleteren & De Vreese,1996)。这是由糖酵解、氧化磷酸化、TCA循环、糖异生等代谢途径中的酶活性下降所致(O′Riordan & Burnell,1989)。芯片和SAGE研究发现,甘油三酯在C.elegansdauer虫态中被转变为糖(Holt & Riddle,2003; Jonesetal.,2001; McElweeetal.,2004; Wang & Kim,2003),表明该虫态可以利用脂类物质为自身提供能量。但是,在松材线虫表达谱中,相对于繁殖型虫态,扩散型LⅣ在三大代谢通路中的大部分基因下调表达,只有个别基因上调表达,推测松材线虫LⅣ的基础代谢低于C.elegansdauer虫态,两者的代谢与维持机制有所差异,具体的差别有待深入研究。另外,松材线虫LⅢ和LⅣ中山梨醇脱氢酶基因的表达量均上调,与该基因在C.elegansdauer虫态中的表达相一致。该酶在果糖到山梨醇的可逆氧化还原反应过程中起催化作用(Holt & Riddle,2003),可能参与无氧状态下的乙醇发酵(ethanol fermentation)过程(Hu,2007; McElweeetal.,2006)。推测此基因在松材线虫中具有类似作用,可能是为适应不良的环境条件而启动的特殊代谢方式。LⅢ在脂肪酸β-氧化/脂肪酸合成代谢途径中相关基因显著上调表达,说明此发育阶段在大量积累脂质,为转变为LⅣ做准备。
总之,在生理状态的维持、对不良环境的耐受以及基础代谢率降低等方面,松材线虫扩散型虫态的基因表达模式与模式线虫C.elegansdauer虫态类似;但是,作为植物寄生线虫,松材线虫在传播扩散过程中又有媒介昆虫的参与,使其基因表达模式变得更加复杂和特殊。
王海英, 魏广兵, 刘影霞, 杜贝贝, 岳玲, 黄涛, 王立鹏, 徐世清, 司马杨虎, 2016. 基于数字基因表达谱(DGE)分析异烟肼诱导家蚕脂肪体损伤的差异表达基因. 蚕业科学, 42(3): 404-414.
姚敏磊, 张璟曜, 周汐, 韩少怀, 谢凤斌, 朱月林, 盖钧镒, 杨守萍, 2016. 大豆响应低磷胁迫的数字基因表达谱分析. 大豆科学, 35(2): 213-221.
詹开瑞, 游源浅, 张晓燕, 陈艳, 2013. 甲酸乙酯对松材线虫的熏蒸效果. 生物安全学报, 22(2): 127-131.
BAERMANN G, 1917. Eine einfache methode zur auffindung von ankylostomum (nematoden) larven in Erdproben.GeneeskundigTijdschriftvoorNederlandsch-IndieBatavia, 57: 131-137.
BARGMANN C I, 2006.ChemosensationinC.elegans. (2006-10-25)[2016-11-02]. https:∥www.ncbi.nlm.nih.gov/books/NBK19746/.
BURNELL A M, O′HALLORAN D M, 2004. Chemoreceptor genes: what can we learn fromCaenorhabditiselegansand how can we apply this information to studies on other nematodes?NematologyMonographs&Perspectives, 2: 707-714.CASSADA R C, RUSSELL R L, 1975. The dauerlarva, a post-embryonic developmental variant of the nematodeCaenorhabditiselegans.DevelopmentalBiology, 46(2): 326-342.CHEN N S, PAI S, ZHAO Z Y, MAH A, NEWBURY R, JOHNSEN R C, ALTUN Z, MOERMAN D G, BAILLIE D L, STEIN L D, 2005. Identification of a nematode chemosensory gene family.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 102(1): 146-151.
ENDA N, IKEDA T, 1983. Role of volatiles of a pine tree as emerging stimulants for attracting the pine wood nematode from the pine sawyer.Transactionsofthe94thAnnualMeetingJapaneseForestrySociety, 94: 479-480.
FRECKMAN D W, MANKAU R, FERRIS H, 1975. Nematode community structure in desert soils — nematode recovery.JournalofNematology, 7(4): 343-346.GOLDEN J W, RIDDLE D L, 1984. TheCaenorhabditiselegansdauer larva: developmental effects of pheromone, food, and temperature.DevelopmentalBiology, 102(2): 368-378.HOLT S J, RIDDLE D L, 2003. SAGE surveysC.eleganscarbohydrate metabolism: evidence for an anaerobic shift in the long-lived dauer larva.MechanismsofAgeingandDevelopment, 124(7): 779-800.
HOUTHOOFD K, BRAECKMAN B P, LENAERTS I, BRYS K, DE VREESE A, VAN EYGEN S, VANFLETEREN J R, 2002. Ageing is reversed, and metabolism is reset to young levels in recovering dauer larvae ofC.elegans.ExperimentalGerontology, 37(8): 1015-1021.
HU P J, 2007.Dauer. (2007-08-08)[2016-10-06]. https:∥www.ncbi.nlm.nih.gov/books/NBK116082/.ISHIBASHI N, KONDO E, 1977. Occurrence and survival of the dispersal forms of pine wood nematode,Bursaphelenchuslignicolus.AppliedEntomologyandZoology, 12(4): 293-302.ISHIKAWA M, SHUTO Y, WATANABE H, 1986. Beta-myrcene, a potent attractant component of pine wood for the pine wood nematode,Bursaphelenchusxylophilus.AgriculturalandBiologicalChemistry, 50(7): 1863-1866.
JONES S J M, RIDDLE D L, POUZYREV A T, VELCULESCU V E, HILLIER L, EDDY S R, STRICKLIN S L, BAILLIE D L, WATERSTON R, MARRA M A, 2001. Changes in gene expression associated with developmental arrest and longevity inCaenorhabditiselegans.GenomeResearch, 11(8): 1346-1352.
KIKUCHI T, AIKAWA T, KOSAKA H, PRITCHARD L, OGURA N, JONES J T, 2007. Expressed sequence tag (EST) analysis of the pine wood nematodeBursaphelenchusxylophilusandB.mucronatus.MolecularandBiochemicalParasitology, 155(1): 9-17.KIM K, SATO K, SHIBUYA M, ZEIGER D M, BUTCHER R A, RAGAINS J R, CLARDY J, TOUHARA K, SENGUPTA P, 2009. Two chemoreceptors mediate developmental effects of dauer pheromone inC.elegans.Science, 326: 994-998.LIU T, ZIMMERMAN K K, PATTERSON G I, 2004. Regulation of signaling genes by TGF-beta during entry into dauer diapause inC.elegans.BMCDevelopmentalBiology, 4(1): 1.MAMIYA Y, 1983. Pathology of the pine wilt disease caused byBursaphelenchusxylophilus.AnnualReviewofPhytopathology, 21(1): 201-220.
MCELWEE J J, SCHUSTER E, BLANC E, THOMAS J H, GEMS D, 2004. Shared transcriptional signature inCaenorhabditiselegansdauer larvae and long-lived daf-2 mutants implicates detoxification system in longevity assurance.JournalofBiologicalChemistry, 279: 44533-44543.
MCELWEE J J, SCHUSTER E, BLANC E, THORNTON J, GEMS D, 2006. Diapause-associated metabolic traits reiterated in long-lived daf-2 mutants in the nematodeCaenorhabditiselegans.MechanismsofAgeingandDevelopment, 127(12): 922-936.
O′RIORDAN V B, BURNELL A M, 1989. Intermediary metabolism in the dauer larva of the nematodeCaenorhabditiselegans: 1. glycolysis, gluconeogenesis, oxidative-phosphorylation and the tricarboxylic-acid cycle.ComparativeBiochemistryandPhysiologyB—Biochemistry&MolecularBiology, 92(2): 233-238.
RIDDLE D L, SWANSON M M, ALBERT P S, 1981. Interacting genes in nematode dauer larva formation.Nature, 290: 668-671.ROGERS C M, FRANKS C J, WALKER R J, BURKE J F, HOLDEN-DYE L, 2001. Regulation of the pharynx ofCaenorhabditiselegansby 5-HT, octopamine, and FMRFamide-like neuropeptides.JournalofNeurobiology, 49(3): 235-244.SENGUPTA P, CHOU J H, BARGMANN C I, 1996. odr-10 encodes a seven transmembrane domain olfactory receptor required for responses to the odorant diacetyl.Cell, 84(6): 899-909.STAMPS W, LINIT M, 1998. Chemotactic response of propagative and dispersal forms of the pinewood nematodeBursaphelenchusxylophilusto beetle and pine derived compounds.FundamentalandAppliedNematology, 21(3): 243-250.THOMAS J H, KELLEY J L, ROBERTSON H M, LY K, SWANSON W J, 2005. Adaptive evolution in the SRZ chemoreceptor families ofCaenorhabditiselegansandCaenorhabditisbriggsae.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 102(12): 4476-4481.
THOMAS J H, ROBERTSON H M, 2008. TheCaenorhabditischemoreceptor gene families.BMCBiology, 6(1): 42.
VANFLETEREN J R, DE VREESE A, 1996. Rate of aerobic metabolism and superoxide production rate potential in the nematodeCaenorhabditiselegans.JournalofExperimentalZoology, 274(2): 93-100.
WANG J, KIM S K, 2003. Global analysis of dauer gene expression inCaenorhabditiselegans.Development, 130(8): 1621-1634.
WINGFIELD M J, BLANCHETTE R A, NICHOLLS T H, ROBBINS K, 1982. The pine wood nematode: a comparison of the situation in the United-states and Japan.CanadianJournalofForestResearch—RevueCanadienneDeRechercheForestiere, 12(1): 71-75.YAN X, CHENG X Y, WANG Y S, LUO J, MAO Z C, FERRIS V R, XIE B Y, 2012. Comparative transcriptomics of two pathogenic pinewood nematodes yields insights into parasitic adaptation to life on pine hosts.Gene, 505(1): 81-90.ZHAO L, MOTA M, VIEIRA P, BUTCHER R A, SUN J, 2014. Interspecific communication between pinewood nematode, its insect vector, and associated microbes.TrendsinParasitology, 30(6): 299-308.
ZHAO L, WEI W, LIU X, KANG L, SUN J, 2007. A novel rapid sampling method for pinewood nematode,Bursaphelenchusxylophilus(Nematoda : Parasitaphelenchidae).CanadianJournalofForestResearch—RevueCanadienneDeRechercheForestiere, 37(10): 1867-1872.
ZHAO L, ZHANG S, WER W, HAO H, ZHANG B, BUTCHER R A, SUN J, 2013. Chemical signals synchronize the life cycles of a plant-parasitic nematode and its vector beetle.CurrentBiology, 23(20): 2038-2043.
(责任编辑:杨郁霞)
Comparative analysis of dispersal and propagative larval stages in the pinewood nematode (Bursaphelenchusxylophilus) based on a digital gene expression library
TIAN Haokai1,2, ZHANG Shuai3, LIU Xiaolong1,2, ZHANG Zhiwei1*, ZHAO Lilin2*
1CollegeofForestry,ShanxiAgriculturalUniversity,Taigu,Shanxi030801,China;2StateKeyLaboratoryofIntegratedManagementofPestInsectsandRodents,InstituteofZoology,ChineseAcademyofSciences,Beijing100101,China;3StateKeyLaboratoryforBiologyofPlantDiseasesandInsectPests,InstituteofPlantProtection,ChineseAcademyofAgriculturalSciences,Beijing100193,China
【Aim】 The formation of the dispersal larva plays an important role in the transmission and spreading of pinewood nematode (Bursaphelenchusxylophilus) which is a major alien invasive pests of forest worldwide. However, its mechanism about formation and maintenance remains unclear. 【Method】 In this study, digital gene expression (DGE) libraries of two dispersal larvae [3rd instar (LⅢ), and 4th instar (LⅣ)] and of the propagative larva (Ln) were constructed to analyze the differences of gene expression of different developmental stages from aspects of maintenance of diapause, chemoreception and metabolic pathways. 【Result】 A total of 11184, 8533, 10781 genes were found for LⅢ, LⅣand Lnaccording to reference genome, respectively. Compared with Ln, most of the genes in LⅣwere down-regulated, and some genes, including chemoreceptor genes, nuclear hormone receptors and other metabolism related genes were up-regulated. The probable function of these genes is in regulating diapause stages, chemosensation and vector/host interaction in the dispersing larva. Gene ontology and pathway clustering analysis showed that most metabolic pathway in LⅣwere down-regulated. However, genes related to metabolic pathways in LⅢexpressed vigorously. 【Conclusion】 This results are consistent with phenotype of the physiological status of LⅣ, which does not feed and has low level of overall metabolism.
pinewood nematode; dispersal larva; digital gene expression; differential gene expression
2016-12-11 接受日期(Accepted): 2017-01-20
林业公益性行业科研专项(201204501); 国家自然科学基金(31572272,31272323); 中国科学院创新工程项目(KSCX2-EW-J-2); 中国科学院前沿科学重点研究项目(QYZDB-SSW-SMC014); 国家高技术研究发展计划(“863”计划: 2014AA020529)
田浩楷, 男, 硕士研究生。 研究方向: 森林保护学。 E-mail: tianhaokai2013@163.com
*通信作者(Author for correspondence): 张志伟, E-mail: nmgzhiwei@163.com; 赵莉蔺, E-mail: zhaoll@ioz.ac.cn
10. 3969/j.issn.2095-1787.2017.02.002
