食物浓度和温度对萼花臂尾轮虫种群动态的影响
2017-05-17邓平张生元艾桃山喻运珍
邓平 张生元 艾桃山 喻运珍


摘要:以浓度分别为2.5×106、5×106、10×106、20×106cells/mL的蛋白核小球藻为食物,在温度分别为25、28、30℃条件下对萼花臂尾轮虫进行群体累积培养研究。结果表明,食物浓度对轮虫的种群动态、最大种群密度、种群增长率及非混交卵的日产生量动态、产量均有明显影响;温度对轮虫的种群动态、最大种群密度及非混交卵的产量、形成效率、最大形成效率均有显著影响。食物浓度为20×106cells/mL的小球藻投喂轮虫,在25~30 qC均能达到较高的种群密度,但25℃培养的轮虫较28℃和30℃推迟到达峰值。非混交卵的日产生量与种群密度有相似的变化趋势,28℃和30℃培养的轮虫非混交卵较25℃提前到达峰值。当轮虫的种群密度还处于增长阶段,非混交卵日生产量已达峰值。因此在轮虫的培养过程中可考虑监测非混交卵的动态变化预测轮虫密度达到峰值的时间。
关键词:萼花臂尾轮虫;种群;非混交卵;食物浓度;温度
中图分类号:S955.3+2 文献标志码:A 文章编号:1002—1302(2016)01—0251—03
食物浓度和温度是影响轮虫种群动态的2个重要生态因子。有关食物浓度和温度对轮虫种群动态的影响研究已有较多报道,内容涉及温度和食物浓度对轮虫种群增长率、最大种群密度、怀卵率、休眠卵产量及形成效率等方面的影响。研究表明温度和食物浓度对轮虫的种群增长及休眠卵形成均有明显的影响,但不同种类、同种种类的不同品系得出的种群增长最适温度和最适食物浓度各不相同。迄今为止,关于食物浓度和温度对轮虫非混交卵影响方面的研究未见报道。非混交雌体所产的卵为非混交卵,与轮虫种群数量密切相关。
萼花臂尾轮虫(Brachionus calyciflorus)是淡水中常见的轮虫种类,也是淡水轮虫工厂化培养的首选种类。本研究以萼花臂尾轮虫为对象,研究了适宜温度范围内不同食物浓度对轮虫种群及非混交卵的影响,以丰富此方面的内容,同时为该轮虫的规模化培养提供理论参考。
1材料与方法
1.1轮虫来源及饵料
萼花臂尾轮虫于2012年6月采自湖北省武汉市水产科学研究所旁的水库,所采水样首先用多层纱布滤去其中的枝角类和桡足类等大型浮游动物及杂质,然后置于解剖镜下观察,用微吸管吸出其中的萼花臂尾轮虫并接种到事先准备好的轮虫培养液中进行培养。试验时先用纱布过滤,去除在培养轮虫的过程中产生的废弃物和剩余饵料的絮凝,再用300目的筛绢滤取轮虫,并用蒸馏水冲洗轮虫数次。
选用蛋白核小球藻(Chlorella pyrenoidsa)作为培养轮虫的饵料。藻种购自中国科学院水生生物研究所。小球藻采用sE培养液在光暗周期12 h-12 h实验室条件下培养,光照度2 000~3 000 k,培养温度25℃,充气培养。每7~10 d转接1次。投喂轮虫的藻液经300目筛绢过滤,以去除藻液中的絮凝。
1.3计算方法
轮虫及其非混交卵的计数方法、有关参数的定义和计算方法均与文献[7]相同。
1.4统计分析
不同培养条件下的轮虫种群数量及非混交卵日产生量的动态变化采用重复测量的方差分析(repeated measure ANO-VA)。对不同食物浓度、不同温度条件下的轮虫种群最大密度、种群增长率的差异显著性采用两因素方差分析进行分析。对非混交卵的产量、形成效率及最大形成效率的差异显著性也采用两因素方差分析(two-way ANOVA)进行分析(SPSS19.0.P<0.05)。
2结果与分析
2.1食物浓度和温度对轮虫种群增长的影响
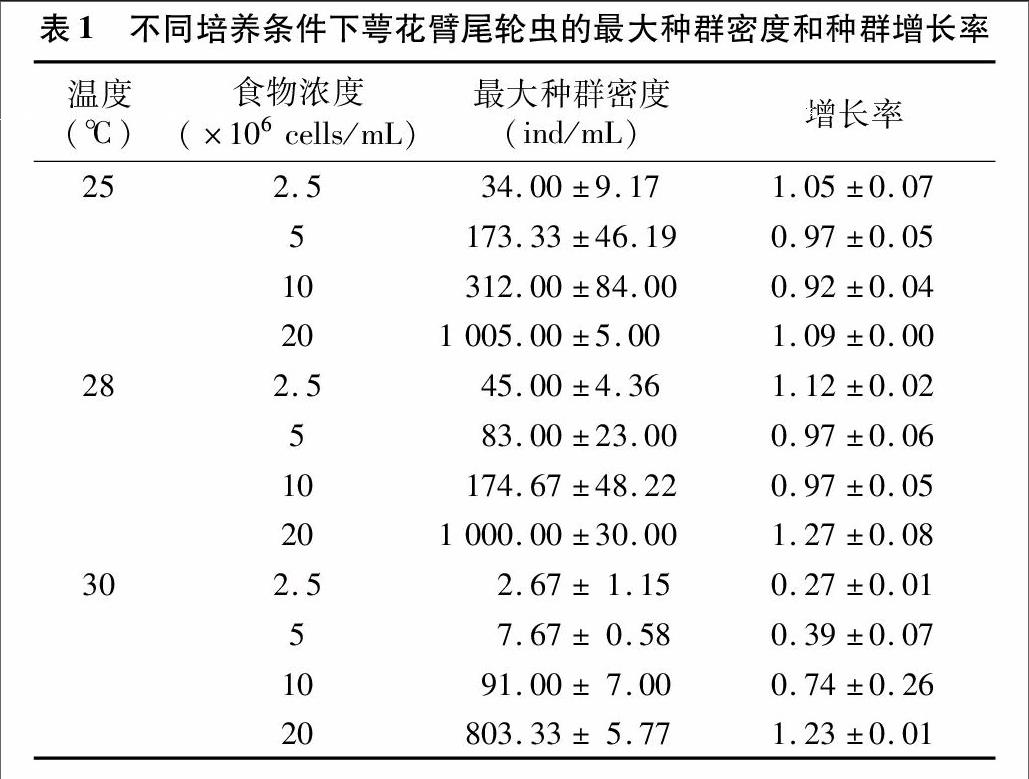
2.1.1食物浓度和温度对轮虫种群数量的影响 在各食物浓度及温度条件下,轮虫种群的动态变化如图1所示。重复测量的方差分析结果表明食物浓度和温度对轮虫种群动态有显著作用(P<0.05)。随着培养溫度的升高,轮虫繁殖高峰值出现的时间也总体上呈现逐渐前移的趋势。温度为28、30℃,食物浓度为20×106 cells/mL,培养4~6 d轮虫种群的密度培养增长极显著(P<0.01),然后又迅速下降。25℃时,20×106 cells/mL食物浓度下,轮虫种群密度从培养5 d开始显著增长(P<0.05)。
2.1.2食物浓度和温度对轮虫最大种群密度和种群增长率的影响各条件组合下,轮虫的最大密度和种群增长率见表1。两因素方差分析结果表明,温度、食物浓度以及两者之间的交互作用均对最大种群密度具有极显著影响(P<0.01)。种群增长率的两因素方差分析结果显示,温度30℃与其他2个温度之间的种群增长率差异极显著(P<0.01),而25℃和28℃条件下种群增长率无明显差异(P>0.05);食物浓度为20×106 cells/mL时轮虫的种群增长率远高于其他3种食物浓度(P<0.01);温度和食物浓度的交互作用对轮虫的种群增长率具有极显著影响(P<0.01)。
2.2食物浓度和温度对轮虫非混交卵的影响
2.2.1食物浓度和温度对轮虫非混交卵日产生量的影响 各食物浓度及温度条件下,轮虫非混交卵日产生量的动态变化如图2所示。重复测量的方差分析结果表明,食物浓度为20×106 cells/mL的非混交卵形成动态与其他各组差异极显著(P<0.01),30℃条件下除外。25℃食物浓度为2.5×106 cells/mL轮虫非混交卵增长不明显,其他3个食物浓度培养4 d非混交卵日产生量开始增加,20×106cells/mL食物浓度的非混交卵日产生量在培养6 d时达到峰值105 ind/mL。28℃条件下各食物浓度均在培养3 d非混交卵日产生量开始增加,20×106 cells/mL食物浓度的非混交卵日产生量在培养4 d达到峰值131 ind/mL。30℃条件下,20×106 cells/mL食物浓度的非混交卵日产生量在培养4 d显著增长,培养5 d达到峰值90 ind/mL,其他3个食物浓度非混交卵日产生量增长不明显。
2.2.2食物濃度和温度对轮虫非混交卵产量和形成效率的影响 在轮虫非混交卵的第1个形成周期内,各条件组合下非混交卵的产量、形成效率和最大形成效率见表2。两因素方差分析结果显示,在培养全过程中,温度、食物浓度以及两者之间的交互作用均对非混交卵产量具有极显著影响(P<0.01)。对非混交卵形成效率进行的两因素方差分析结果表明,温度对非混交卵的形成效率具有极显著影响(P<0.01);食物浓度对非混交卵形成效率没有明显作用(P>0.05);温度与食物浓度的交互作用对非混交卵的形成效率具有极显著影响(P<0.01)。对最大形成效率进行的两因素方差分析结果显示,温度对非混交卵的最大形成效率具有极显著影响(P<0.01);食物浓度为2.5×106cells/mL的最大形成效率与其他各组差异极显著(P<0.01);温度与食物浓度的交互作用对非混交卵的最大形成效率具有极显著影响(P<0.01)。
3讨论
3.1食物浓度和温度对轮虫种群增长的影响
已有研究结果表明,同一温度下萼花臂尾轮虫种群密度随食物浓度的升高而增加。本研究结果与已有的研究结果一致,25、28、30℃条件下萼花臂尾轮虫的种群动态与食浓度密切相关。在同一食物浓度下,种群密度随温度的升高而增大。本研究中25℃和28℃条件下各食物浓度条件下的种群密度有相同的变化趋势,但30℃条件下各食物浓度没有表现出这种变化趋势,可能与较高温度条件对低起始物密度的轮虫种群增长有抑制作用有关。
黄祥飞研究发现,萼花臂尾轮虫种群的最大密度随温度的升高而有所增加,而种群达到高峰所需的时间则随着温度的升高而缩短。本研究结果也表明,轮虫种群在28℃和30℃条件下较25℃提前1 d达到最大密度,但25℃与28℃条件下的轮虫种群增长率差异不显著,且明显高于30℃。
3.2食物浓度和温度对轮虫非混交卵的影响
已有关于轮虫繁殖方面的研究主要是通过考察轮虫的怀卵量这一指标,怀卵轮虫既有产非混交卵的非混交雌体又有产休眠卵的混交雌体。非混交雌体与轮虫种群增长密切相关,考察非混交雌体所产非混交卵的动态变化将能更准确地预测出轮虫种群密度的变化。本研究中同一温度下萼花臂尾轮虫种群密度随食物浓度的升高而增加,非混交卵的日产生量也表现出相似的变化趋势。同一食物浓度下,非混交卵动态与种群数量动态也表现出极大的相似性,即25℃和28℃条件下各食物浓度条件下的非混交卵日产生量与种群密度有相同的变化趋势,30℃条件下非混交卵日产生量没有表现出这种变化趋势。
温度为25℃时轮虫的非混交卵产量、形成效率和最大形成效率均明显高于28℃和30℃条件下,这可能与25℃条件下轮虫非混交卵的日产生量增长时间较长有关。研究还发现当轮虫的种群密度还处于增长阶段,非混交卵日生产量已达峰值。因此可用非混交卵的动态变化提前预测出轮虫种群的动态变化。
